

















« Prev Next »
In humans and many other animal species, sex is determined by specific chromosomes. But how did researchers discover these so-called sex chromosomes? The chromosomes that carry those genes related to sexual characteristics have a long and intriguing story of exploration. As often happens during such a long course of discovery, scientists noted the appearance of these chromosomes before knowing exactly what these structures were.
The X Element
The first inkling that sex chromosomes were unique from all other chromosomes came from experiments conducted by German biologist Hermann Henking in 1891. By this point in time, scientists could easily view mitotic cell division using light microscopy. Indeed, while using this method to study sperm formation in wasps, Henking saw that some wasp sperm cells had 12 chromosomes, while others had only 11. Moreover, while observing the stages of meiosis that formed these sperm cells, Henking noticed that the mysterious twelfth chromosome looked different from all the others. He thus named this chromosome the "X element," to represent its unknown nature. Interestingly, in subsequent studies of female grasshopper egg formation, Henking was unable to spot the X element (Brown, 2003; Pierce, 2005). Based on these observations, Henking hypothesized that this extra chromosome must play some role in sex determination in insects. However, he was unable to gather any direct evidence that supported this hypothesis.
The Accessory Chromosome
About 10 years after Henking's studies, American zoologist C. E. McClung began extensive research into the idea that the X element must have something to do with sex determination. First, McClung asserted that this strange chromosome be called the "accessory chromosome," because it appeared to have a separate purpose compared to the other chromosomes. Next, he decided to undertake a wide comparative study of spermatogenesis not just in insects but in many different organisms, including spiders and mice. While observing sperm generation in grasshoppers, McClung noted that the accessory chromosome did not behave as the other chromosomes did during the first meiotic division. Specifically, this "twelfth chromosome" was not distributed equally to the four sperm cells that arose from one grasshopper spermatogonium; instead, it was present in only half of them. This consistent pattern of segregation in the spermatogenesis of many different organisms caused McClung to speculate that there must be some fundamental reason for the generation of two types of sperm cells. McClung thought about the consequences of this 50/50 distribution of the accessory chromosome, and he pondered how this distribution probably resulted in two different kinds of zygotes. He also noted that there was really only one characteristic that varied among zygotes of many different species in 50/50 proportions, and that characteristic was sex. Thus, when he assembled all the evidence together, McClung came up with the proposal that the accessory chromosome must influence the determination of sex in a zygote (Brown, 2003). McClung was so eager to formally communicate his hypothesis that he wrote a preview of this idea (McClung, 1901) that was published as a short letter approximately a year before his full manuscript appeared in a journal (McClung, 1902).
At around the same time, Walter Sutton, who was one of McClung's former students, published a separate paper that fully examined the chromosomes in the grasshopper and provided data that strongly supported McClung's hypothesis regarding the accessory chromosome. In his study, Sutton closely followed the path of chromosomes throughout meiosis in sperm cells, and he demonstrated with drawings of successive meiotic stages that sperm cells contained half the total number of chromosomes present in grasshopper zygotes. Sutton's careful consideration of how chromatic material segregated into sperm cells became the launching point for the chromosome theory of inheritance. To develop this theory, Sutton counted every chromosome present in grasshopper sperm cells and measured their size. He then ranked each chromosome according to these size measurements. Sutton's careful observation of chromosome number and segregation led him to conclude that gametes carry one-half of the total chromosomal collection in a zygote; he also determined that the merging of two gametes during fertilization must be the mechanism by which chromosome pairs are reformed.
Throughout the many observations that led to this conclusion, Sutton made careful note of the path of the accessory chromosome. Based on these observations, he asserted that, though it was different in its segregation into sperm cells, the accessory chromosome was indeed a true chromosome (Sutton, 1902).
Counting Chromosomes
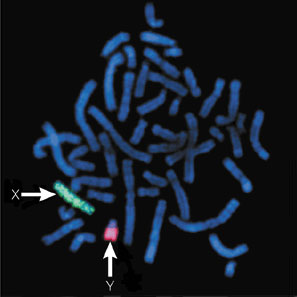
Figure 1: Male human chromosomes.
X and Y chromosomes are indicated by arrows.
Courtesy of Betsy Hirsch and Olga Shilko. All rights reserved. 
Following Wilson's discovery, there emerged some confusion about what to call these apparent differences in chromosome sorting seen in each sex. Part of this confusion related to the fact that numerous species were being examined, and these species varied in terms of chromosomal number and arrangement. Thus, challenges remained to the development of a unifying theory about how chromosomes actually determine an organism's sex.
Sex Is Determined by Different Mechanisms in Different Organisms
One primary question facing the scientists of the time was whether maleness was associated with the absence or the presence of the misshapen chromosome. Yet again, E. B. Wilson set out to find the answer. Wilson took a deep interest in the work of his colleague Nettie Stevens, and he set out to combine her surveys of multiple beetle species with his. Stevens (1905) had observed that in the Tenebrio species of beetle, there was an unusual-looking pair of chromosomes that separated to form different sperm cells. Based on comparisons of chromosome appearance in other cells of male and female Tenebrio, she proposed that these accessory chromosomes were likely related to the inheritance of sex. Wilson independently observed similar patterns in a variety of insects. Bolstered by Stevens' work, Wilson thus proposed that, among the two recognizable "sex chromosomes," it was possible that one was a "female-determinant" and the other was a "male-determinant" (Wilson, 1905). However, in some insects, including wasps, maleness was associated with the absence of an extra chromosome. How, then, could the observations in beetles be reconciled with those in wasps?
In an attempt to solve this dilemma, Wilson tried to organize recent data from his and other labs along with the older observations of Henking, and he began with chromosomal counts. Specifically, Wilson compared the number of chromosomes in the regular somatic cells of each species with the number of chromosomes in these species' germ cells. Based on his findings, he proposed that two categories of chromosomal pairing patterns existed, and he denoted these categories Type A and Type B (Table 1).
Table 1: Chromosome Counts in the Somatic Cells of Several Type A and Type B Species
|
Species (Type A) |
# Chromosomes in Female Cells | # Chromosomes in Male Cells |
Species (Type B) |
# Chromosomes in Female Cells | # Chromosomes in Male Cells |
| Protenor belfragei (broad-headed bug) | 12 | 11 |
Lygaeus turcicus (milkweed bug) |
14 | 14 |
| Anasa tristis (squash bug) | 22 | 21 |
Euschistus fissilis
(stink bug) |
14 | 14 |
|
Alydus pilosulus
(broad-headed bug) |
14 | 13 |
Coenus delius (stink bug) |
14 | 14 |
(Adapted from Wilson, 1905)
As illustrated in Table 1, the somatic cells from Type A males had one less chromosome than the somatic cells of females of the same species. In the Type B species, however, the chromosome number was the same in both sexes. Therefore, sex could not be determined in a universal way by the number of chromosomes in organisms' regular somatic cells.
But how did these chromosome counts in somatic cells relate to the counts in gametes? Wilson argued that the number of chromosomes in an organism's gametes depended on whether the organism was from a Type A species or a Type B species. Wilson then defined the two groups based on how their sex chromosomes separated into gametes. His descriptions were as follows, and they are also summarized in Table 2:
- Type A species: In organisms of these species, male somatic cells have one unpaired chromosome, the accessory. So, while females have n chromosomes, males will always have n-1 chromosomes. Furthermore, female gametes (eggs) will always have n/2 chromosomes. On the other hand, in sperm formation, the unpaired accessory chromosome passes into only half of the sperm cells, so half will have this chromosome, while the other half will be without it.
- Type B species: In organisms of these species, all chromosome pairs are of equal size in female somatic cells, and each chromosome has a partner. In male somatic cells, all chromosome pairs are of equal size except for one pair—these are the "idiochromosomes." In fact, the smaller, more crumpled-looking member of the pair is only present in male cells. Both eggs and sperm from these species contain n/2 chromosomes. However, in sperm cells, the unequally sized chromosomes separate into two different types of male gametes, one with the larger idiochromosome X and one with the smaller, crumpled looking idiochromosome.
Table 2: Chromosome Counts in Somatic Cells and Gametes of Type A and Type B Species
| Type A | Type B | ||||
| Somatic Cell | Gamete | Somatic Cell | Gamete | ||
| Female | Chromosome Number | n | n/2 | n | n/2 |
| Chromosome Appearance | Pairs are of similar size | All eggs contain one of each chromosome pair | All pairs are of equal size | All eggs contain one of each chromosome pair | |
| Male | Chromosome Number | n-1 |
(n-1)/2 = 50%; n/2 = 50% |
n | n/2 |
| Chromosome Appearance | One unpaired chromosome | 50% of cells contain the accessory chromosome; 50% do not | One pair is mismatched in size (i.e., the idiochromosomes) | 50% of cells contain the smaller chromosome; 50% contain the larger one | |
Wilson's Model: Type A and Type B Sex-Determination Systems
E. B. Wilson's proposal from his observations about chromosome number was that there were two types of systems by which these mismatched chromosomes separated into gametes. The Type A system resulted in an X or no-X distribution among sperm, while the Type B system resulted in an X or crumpled-looking X partner distribution among sperm. The relationship between the observations made in the gametes and the counts in the species' somatic cells confirmed that patterns seen in gametes directly corresponded with the sex of individual organisms. The Type A system later became known as the XX/XO system, wherein sperm cells with no X chromosome determined maleness. The Type B system became known as the XX/XY system, wherein the sperm cells with the crumpled-looking X partner (renamed the Y chromosome) determined maleness. Later research showed that males aren't always the sex with the deviation from identical chromosome pairs, however. For instance, in birds, snakes, and various insects, there is a ZZ/ZW system, wherein females carry the mismatched chromosome pair (ZW).
Lessons Learned
With this combination of evidence from different species, a connection was observed between specific chromosomes and the sex of an individual. Interestingly, it was the pursuit of sex determination that enabled scientists to see a direct link between chromosome distribution and specific traits. Sutton's suggestion that the gamete chromosome separation pattern was similar to the proportions of male and female offspring opened up the possibility that chromosomes had something to do with inherited traits. But it wasn't until a wide variety of species were investigated that some of the mysteries of the accessory chromosome could be solved. For his broad analysis of different species' chromosome counts, E .B. Wilson is credited with the idea that there can be different types of sex chromosome systems among different animals (Wilson, 1910). However, the initial idea that sexual characteristics were physically represented by chromosomes and inherited through the transfer of chromatin originated with Sutton and his careful chromosome counts in the grasshopper.
References and Recommended Reading
Brown, S. Entomological contributions to genetics: Studies on insect germ cells linked genes to chromosomes and chromosomes to Mendelian inheritance. Archives of Insect Biochemistry and Physiology 53, 115–118 (2003)
Henking, H. Über Spermatogenese und deren Beziehung zur Entwicklung bei Pyrrhocoris apterus L. Zeitschrift für wissenschaftliche Zoologie 51, 685–736 (1891)
McClung, C. E. Notes on the accessory chromosome. Anatomischer Anzeiger 20, 220–226 (1901) (link to article)
———. The accessory chromosome-Sex determinant? Biological Bulletin 3, 43–84 (1902) (link to article)
Pierce, B. Genetics: A Conceptual Approach, 2nd ed. (New York, W. H. Freeman, 2005)
Stevens, N. M. Studies in spermatogenesis with especial reference to the "accessory chromosome." Carnegie Institution of Washington Publication 36, 1–33 (1905) (link to article)
———. Studies in spermatogenesis, part II: A comparative study of the heterochromosomes in certain species of Coleoptera, Hemiptera, and Lepidoptera, with especial reference to sex determination. Carnegie Institution of Washington Publication 36, 1–43 (1906) (link to article)
Sutton, W. S. On the morphology of the chromosome group in Brachystola magna. Biological Bulletin 4, 24–39 (1902) (link to article)
Wilson, E. B. The chromosomes in relation to determination of sex in insects. Science 22, 500–502 (1905) (link to article)
———. Selective fertilization and the relation of the chromosomes to sex-production. Science 32, 242–244 (1910) doi:10.1126/science.32.816.242 (link to article)










