
Abstract
Controlled sediment flushing operations (CSFOs) allow to recover reservoirs storage loss while rebalancing the sediment flux interrupted by dams but, at the same time, may cause unacceptable ecological impact. In this study, we investigated the responses of the food web of an upland stream to a CSFO, focusing on the effects of fine sediment deposition detected in three different mesohabitats, i.e., a pool, a riffle, and a step-pool. The field campaign lasted two years and included repeated measurements of fine sediment deposits, and sampling of periphyton, benthic macroinvertebrates and fishes. A moderate and patchy deposition occurred due to the CSFO with short and medium-term ecological impact on the lower trophic levels of the food web, which may affect the whole ecosystem functioning. The monitoring of all available mesohabitats in the investigated stream allowed to detect variations in the ecological response to CSFO, providing a more adequate assessment of the impact. As expected, sedimentation was larger in the pool but, in contrast to our hypotheses, the impact was lower and the recovery was longer for the benthic organisms inhabiting the riffle. In the case of fishes, no lethal impact of both brown trout and bullhead was recorded in the short term but the occurrence of longer lasting effects could not be excluded. To date, this is one of the few studies dealing with a detailed integrative assessment of the downstream impact of sediment management from reservoir on both abiotic and biotic components of stream ecosystem.
Similar content being viewed by others

Introduction
The natural sediment flux is severely reduced by dams, causing loss of reservoirs storage and downstream sediment imbalance, propagating up to the sea shorelines1,2. The awareness that storage provided by reservoirs is a strategic non-renewable resource, requiring integrated sustainable management to guarantee intergeneration equity is now firmly accepted worldwide3,4,5,6. Therefore, improving sediment management in regulated catchments is a global imperative.
The evacuation of fine sediment (d < 2 mm7, where d is particle diameter) through the low-level outlets of a dam after complete drawdown (i.e., “empty flushing” or simply “flushing”1,8) is currently performed to tackle reservoirs loss of storage by siltation and is considered a relatively simple and low-priced management alternative4,9. On the one hand, evacuating fine sediment downstream from reservoirs can rebalance, at least partly, the natural sediment flux interrupted by dam structures, thus mitigating the drawbacks of sediment trapping by reservoirs and related habitats loss4,10. However, several issues arise from the sudden increase of sediment load and subsequent deposition characterizing the management of reservoir siltation by sediment flushing operations11,12. In this perspective, sediment sluicing during floods offers improved environmental compatibility though it cannot always be implemented due to hydrological and technical constraints (e.g., absence of high-capacity low-level outlets1,13,14).
The increase in sediment loading during flushing can severely affect the biota in river reaches downstream from the flushed reservoir, at least in the short term12,15,16. Consequently, in the last years, attempts to control sediment flushing operations (i.e., controlled sediment flushing operations—CSFO) in order to limit their downstream impact have been performed and documented in the literature17,18,19. In summary (for details see Espa et al.11), CSFOs are performed as far as possible according to predefined schedules, setting the time window, the duration, and specific thresholds of the sediment concentration in the evacuated waters. These parameters are determined based on the expected effects of increased sediment loading on selected biological targets (e.g., a fish species and/or the benthic macroinvertebrate community). Control of the sediment concentration during a CSFO is a complex task, with several operational difficulties; it is generally achieved by a combined regulation of bottom sluices opening and water flows and, in some cases, employing mechanical excavators18,20. Real-time measurements of sediment concentrations are a necessary support to CSFOs and are used to modify the workflow in case of unexpected results13. Environmental monitoring and assessment are frequently performed in selected river reaches subjected to the sediment pulse before/during/after the CSFOs to evaluate impact and recovery after the desilting works17,19,21,22.
A further key issue when considering the potential drawbacks of sediment flushing is represented by excessive sediment deposition, i.e., severe alteration of channel morphology, in the river reaches downstream from the flushed reservoir. In addition to compromising possible water use and instream structures1, high sediment accumulation can threaten the freshwater biota in the medium or long term15,23. In fact, excessive deposition of fine sediment can impair the entire food web, from algae composing periphyton to benthic macroinvertebrates and fishes, thus compromising the whole ecosystem functioning24,25,26,27,28,29. Consequently, the extent of downstream deposition deserves special attention when assessing the environmental effects of CSFOs12. Depositional patterns after a sediment pulse may significantly differ between mesohabitats (i.e., relatively homogeneous localized areas of the channel that differ in morphology, depth, velocity, and substrate characteristics from adjoining areas30) due to locally different hydraulic conditions23. In turn, the effects of fine sediment deposits on the biocoenosis inhabiting different mesohabitats can be different, in particular for bottom-dwelling organisms31,32. For these reasons, a thorough assessment of the ecological impact of CSFOs would consider in our opinion possible differences between mesohabitats, especially in case of patchy upland streams33.
Although reservoirs desilted by flushing are commonly small to medium sized8,14, they can have storages encompassing several orders of magnitude. Accordingly, the river reaches downstream from the flushed reservoirs can vary from small mountain streams to large rivers, and consequently the ecological impact of related sediment pulses can be different. Upland streams, whose fine-sediment loading is intermittent (flow related) and are generally characterized by coarser substrates and by the presence of more sediment-sensitive species than large lowland rivers, are likely to display the heavier effects34.
According to available peer reviewed literature, major interest on sediment flushing through reservoirs and related environmental problems is concentrated in the European Alps11,19,21,35,36 and in Eastern Asia37,38,39,40,41,42, where discharging sediment downstream is a commonly adopted management strategy, also in very large reservoirs8,43,44. More recently, also as a consequence of the estimated drop in total reservoir capacity of almost 5% (i.e., 40 Gm3) in the last thirty years, a general rethinking of current approaches is taking place also in the US, with growing interest towards available sediment management strategies, including flushing2,5,9. Anyway, further global issues will likely increase the attention towards sustainable sediment management in regulated river systems over the next few years. These issues include the increasing demand for regulated water and hydroelectricity, pushing the construction of new dams and reservoirs45, and the alteration of rainfall/runoff patterns driven by climate change, which is predicted to intensify sediment pressures on freshwater ecosystems46,47.
In this paper, we present the results of a detailed field investigation performed in a reach of an upland stream after a CSFO through a small hydropower reservoir located in the Central Italian Alps. To our knowledge, this is one of the first attempts to fully describe fine sediment deposition after CSFOs (i.e., amount and temporal evolution of deposits) and the related responses of the entire food web (i.e., periphyton, benthic macroinvertebrates and fishes). Moreover, the monitoring activity involved wider spatial and temporal scales compared to previous studies on the effects of CSFOs11,19,21. In fact, the study reach included three distinct mesohabitats, i.e., a pool, a riffle, and a step-pool, which were repeatedly surveyed for two years in order to answer the following research questions:
-
(i)
How much is the fine sediment settled on/into the superficial substrate of the permanently wetted streambed? What are the depositional patterns and the temporal dynamics of this deposit? More specifically, are there significant differences of depositional patterns and temporal dynamics of the deposit between distinct but contiguous mesohabitats?
-
(ii)
Is there any feedback between differences in sediment deposition and the impact/recovery of the biota after the CSFO? Are all components of the stream food web comparably affected?
-
(iii)
Do our findings provide useful information for improving management and/or monitoring of CSFOs in the investigated setting and in comparable contexts?
We note that these research questions have received so far limited attention by the specialized literature22, though they play a fundamental role in improving the sustainability of sediment management through dammed river systems.
Material and methods
Study context
The monitoring campaign was performed downstream from the Valgrosina Reservoir, regularly flushed in the last years to recover its storage (1.3 Mm3). Here, CSFOs have been carried out almost annually since 2006 (i.e., annually from 2006 to 2012, and then in 2014, 2015, and 2018), following a consolidated protocol20,48 aimed at preserving the good ecological status, sensu Water Framework Directive (2000/60/EC), of the aquatic environments downstream of the flushed reservoir. The CSFOs took place between August and September over approximately two weeks. According to the CSFO schedule, suspended sediment concentration (SSC) in the downstream watercourse usually increased during daytime up to 10 g L−1, and decreased by one order of magnitude overnight, when the dislodging operations by mechanical equipment were interrupted. SSC averaged over the whole operation was constrained below the maximum allowed value of 4 g L−1, calculated with the concentration-duration fish response model developed by Newcombe & Jensen49, aiming to limit the mortality of brown trout Salmo trutta L., i.e., the target species in the study area. Moreover, during the CSFO, the average water discharge was usually quite close to the mean annual natural flow (4.5 m3 s−1). Evacuated sediment, predominantly silt (0.0039–0.0625 mm), amounted to around 20,000 tons per CSFO.
In this work, we investigated the effects of the CSFO performed from September 10 to 21, 2018. It followed the mentioned protocol, though the average SSC was slightly above the mentioned threshold (4.65 g L−1), resulting in 23,400 t of sediment (mostly—80%—composed by silt, see Supplementary Note 1) being evacuated.
Study area
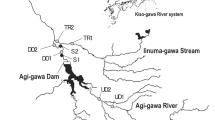
The Valgrosina Reservoir, located at 1,210 m amsl (top of the active pool), is directly supplied by the northern branch of the Roasco Stream and through a short diversion-channel by the eastern branch. These two branches join one kilometer downstream from the reservoir, then the stream flows for 7.8 km up to the Adda River, the main tributary to Lake Como (Fig. 1).

Plane views of the study area, showing the location of the flushed reservoir and the monitored stream reach in northern Italy, with pictures taken during the CSFO performed in September 2018. Top right: satellite imagery from Google Earth Version 7.3.6.9796 (Imagery date: 9/24/2021), accessed on 2/22/2024 (https://earth.google.com/web/@46.35941266,10.21224767,-22238.90048506a,43264.60491051d,35y,-1.76281336h,9.76106037t,359.9611r/data=OgMKATA). Bottom: unpublished photographs by authors S. Quadroni & P. Espa.
The Roasco is a high gradient (0.08 average slope) confined stream, with large stable boulders, and abrupt drops. It flows mainly through a deeply incised canyon in predominantly gneissic rocks. Our field investigation, as well as the control section of the operation, took place along the final 2 km, where the elevation profile is milder (0.025 to 0.045 slope), and access was suitable and safe (Fig. 1).
The natural catchment (145 km2) at the dam section develops in a mountainous area (3,200 m amsl max height), lacking significant human activities; nutrient and organic pollution is indeed negligible in the Roasco Stream50. Additionally, the Valgrosina Reservoir is supplied by a 21—km long tunnel canal, collecting water from an upstream power station plus some diverted streams20, and increasing the overall catchment area (i.e., natural plus artificially connected) to over 700 km2. Accordingly, despite its relatively small storage capacity, the Valgrosina Reservoir supplies the larger peak energy plant of the area (0.43 GW installed capacity, 600 m effective head, 80 m3 s−1 maximum power station discharge), performing regulation of the water volumes coming from the mentioned canal mostly on a daily scale.
Natural runoff in the catchment is essentially driven by snowmelt during spring and early summer, and by rainfall, occasionally intense, in summer and autumn. The Valgrosina Dam was completed in 1960, though the hydropower development of the Roasco Stream dates back to the 1920s, by a small dam located about half a kilometer below the present dam site. Mandatory seasonally-modulated minimum flows of 0.24–0.41 m3 s−1 have been releasing in the Roasco Stream since 2009, and an average streamflow of approximately 1 m3 s−1 was gauged at the study reach during 2010–2014, also due to the contribution by the 15 km2 unexploited catchment50. The stream reach downstream from the Valgrosina Dam is therefore a residual-flow reach, characterized by limited seasonal flow variability, and infrequent flood peaks associated to overspilling of the upstream water-diversion structures.
As previously introduced, we monitored three mesohabitats (i.e., a pool, a riffle and a step-pool), spaced each other by 250 m along the study reach (Fig. 2). Cumulative size distributions from Wolman pebble counts are quite different for the three mesohabitats, particularly for the lower characteristic diameters: d16 was < 2, 20 and 11 mm, d50 was 46, 62 and 58 mm, d84 was 231, 248 and 339 mm for pool, riffle and step-pool, respectively.

Geometric scales of the three mesohabitats considered in this study (L = length, W1 and W2 = upstream and downstream wetted-width, D = average depth measured by sampling at least 15 points at flow rate of 1.2 m3 s−1), and pictures taken before and on three occasions after the CSFO (unpublished photographs by authors S. Quadroni & P. Espa). Pie charts above mesohabitat schemes provide visually-assessed substrate composition for benthic macroinvertebrate sampling (PS = psammal—0.006–2 mm, AK = akal—0.2–2 cm, MIC = microlithal—2–6 cm, MES = mesolithal—6–20 cm, MAC = macrolithal—20–40 cm, MGL = megalithal—> 40 cm).
Sampling of fine sediment deposits, periphyton, and benthic macroinvertebrates was performed five days before (September 5, 2018) and on ten occasions after the CSFO. Specifically, the first post-CSFO survey took place about two weeks after the flushing (October 8, 2018). The further ones were spaced by time intervals of two–three months (December 3, 2018 and March 21, July 9, September 12, 2019 for the first post-CSFO year; October 28, December 9, 2019 and February 10, July 8, September 14, 2020 for the second post-CSFO year). In order to consider all the biological components of the food web, fish sampling was carried out in the final part of the investigated reach, both before (August 21, 2018) and, on two occasions, after the CSFO. Post-CSFO fish sampling occurred less than one month (October 10, 2018) and more than one year after the flushing (January 24, 2020).
Deposition of fine sediment: sampling and analyses
Deposition of fine sediment settled on/into the superficial substrate of the permanently wetted streambed after the CSFO was quantified by a resuspension technique, following Espa et al.20. On each sampling occasion, mesohabitats were sampled in three points, two near the stream edge and one in the center. One sample (1 L) of turbid water per point was collected keeping a McNeil corer51 firmly placed on the streambed, and stirring up suspendable sediment within the tube52. The water depth within the corer was recorded as well to calculate the overall water volume. The samples were gravimetrically analyzed by filtering 200 mL on cellulose filters (0.47 μm pore size) and weighting them after drying in oven at 105 °C for 2 h53. Finally, the mass of fine sediment per unit area was simply computed by dividing sediment mass (product of SSC and water volume in the corer) by the cross-sectional area of the corer tube.
Periphyton sampling and analyses
On each sampling occasion, we collected an integrated sample of periphyton per mesohabitat. Five cobbles were randomly picked up. Periphyton was tooth-brushed from the upper side of the cobbles within two square areas of 10 cm2 each, using a plastic square frame as reference, and then collected in 200 mL water in dark glass bottles. Samples were preserved at 4 °C and analyzed in the laboratory within 24 h. Each sample was filtered on a glass-fiber filter (0.7 μm pore size) until the pores were clogged. Then, chlorophyll-a content (mg m−2) was determined through extraction with acetone and spectrophotometric detection54. The values of the chlorophyll-a content were normalized for the volume filtered.
Benthic macroinvertebrates sampling and analyses
Benthic macroinvertebrates were sampled by Surber sampler with 0.1 m2 area and 500 μm mesh. A proportional stratified random sampling was adopted as indicated by the Water Framework Directive (2000/60/EC): specifically, five subsamples were collected in representative microhabitats, proportionally to their relative abundance in each mesohabitat (Fig. 2), and then integrated in a single sample per mesohabitat.
Samples were fixed with ethanol (99%) and transported to the laboratory, where invertebrates were sorted, identified to genus (only Plecoptera and Ephemeroptera, due to easier identification and dichotomous key availability for Italian benthic macroinvertebrates55,56) or family (all other taxa) level, and counted. Density (individuals m−2) was determined for each taxon. Moreover, for each sampling date, a pool (up to 20 individuals depending on the size) of each taxon was oven-dried at 80 °C for 24 h, and the dry weight was measured to calculate the biomass per unit area. When taxa were poorly represented, it was not possible to achieve the minimum number of individuals necessary to obtain reliable weight estimates on each sampling date, and the average value of the recorded weights was used.
The following community metrics were calculated (Supplementary Table 1): total taxon richness, total density, EPT (Ephemeroptera, Plecoptera and Trichoptera, i.e., the most sensitive insect orders) richness and relative abundance, density of macroinvertebrates belonging to Ecological Group A (ECOgA, sensu Usseglio-Polatera et al.57), i.e., rheophilous taxa preferring coarse substrates, typical of oligotrophic, alpine habitats, and the biomass of trophic guilds (see section below). Moreover, the Siltation Index for LoTic EcoSystems (SILTES), i.e., an index specifically developed to detect siltation impact in the Alpine context, was considered. SILTES is based on taxon and EPT richness, and ECOgA. It was calculated as the average of the three mentioned metrics, scaled over the entire dataset. Specifically, the minimum value of the metric in the dataset was subtracted to the sample value, and the result was divided by the min–max range, according to the following formula:
where m is the observed value of the metric, mmin is the minimum observed value of the metric in the dataset and mmax is the maximum observed value of the metric in the dataset. SILTES ranges from 0 (worst condition) to 1 (best condition), for additional details see Doretto et al.58,59.
Fish sampling and analyses
Three quantitative samplings were carried out by electrofishing (removal method with two passes). Caught fishes were counted, measured for total length and weighted. Density and biomass of the different species were computed by dividing the number and weight of all caught specimens to the sampled area (324 m2). The density reduction obtained by comparing before/after CSFO sampling was considered as apparent mortality, i.e., possible fish migration occurred during the time interval between the two samplings was not accounted for. Moreover, in the investigated reach, fishing is allowed and brown trout juveniles are occasionally restocked, but no data quantifying these practices are available.
Food web analyses
Possible changes in the food web associated to the CSFO were assessed adopting all the biological components monitored in this study: periphyton, fish, and the different trophic guilds in the benthic macroinvertebrate community. For this purpose, macroinvertebrates were pooled according to four trophic guilds60: (i) grazers or scrapers, feeding on periphyton; (ii) shredders, feeding on coarse particulate organic matter (CPOM), i.e., fragments of leaves, plant tissues and wood debris; (iii) collectors, including gatherers and filterers, feeding on fine particulate organic matter (FPOM) and dissolved organic matter (DOM), respectively; (iv) predators, feeding on small animals. The biomass of these trophic guilds was calculated according to the information available in the freshwaterecology.info database v. 7.0–10/201661. Specifically, a fuzzy data coding was used, i.e., each family/genus in the database was associated to ten feeding types, with a value ranging from 0 (no preference) to 10 (complete selectivity for one feeding type). Per each taxon, we multiplied this value by the weight of the individuals counted. The total weight of a single feeding group was calculated summing all the values referring to the feeding type and dividing by 10. The results were finally summarized in the four guilds mentioned above (gatherers-collectors and active and passive filter feeders were grouped into one category, i.e., collectors, while miners, parasites, xylophagous taxa and other feeding types were not considered since they were scarcely represented; Supplementary Table 2).
In order to measure the biomass of the different biological components by a single unit, we estimated the organic carbon content. Epilithic chlorophyll a was converted to algal carbon by assuming a carbon to chlorophyll ratio of 2762,63. The carbon content of fish was estimated by applying a conversion rate of 14.25% of the wet weight, assuming a carbon content of 47.5% of dry weight of fish and a water content of 70%64. Finally, the organic carbon content of benthic macroinvertebrates was estimated by assuming that ash free dry weight is 93.5% of dry weight, and organic carbon is 51% of ash free dry weight, based on the mean value of data reported by Salonen65 and Benke et al.66 for the major insect taxa found in the study area.
Statistical analyses
We analyzed each fine sediment sample separately. Then, we performed both mesohabitat and reach averaging, i.e., we calculated the mean value of the three sampling points per mesohabitat (mesohabitat average), and the mean value of all the nine samples (reach average). Coefficient of variation (CV, i.e., the ratio between standard deviation and average) was computed to assess spatial variability at both mesohabitat and reach scale for each sampling date. The modified Mann–Kendall test67 was applied to test for significant decreasing trend after the CSFO.
In the case of periphyton samples, CV was calculated to assess spatial variability at the reach scale for all the sampling dates. Two-way analysis of variance (ANOVA) and post-hoc Tukey test were performed to test chlorophyll-a content for significant (α < 0.05) differences between mesohabitats during the first (from Oct-18 to Sep-19) and the second (from Oct-19 to Sep-20) post-CSFO year.
Compositional dissimilarity between macroinvertebrates samples was quantified by the Bray–Curtis index. This index ranges from 0 (complete similarity) to 1 (complete dissimilarity). Differences in the community composition between mesohabitats and sampling occasions were visually and statistically examined by non-metric multidimensional scaling (NMDS) and two-way permutational analysis of variance (PERMANOVA) along with permutational multivariate analysis of dispersion (PERMDISP), respectively. After data transformation (density data were log-transformed and percentage data were logit-transformed), two-way ANOVA and post-hoc Tukey test were performed to test each community metric and each trophic guild for significant (α < 0.05) differences between mesohabitats during the first (from Oct-18 to Sep-19) and the second (from Oct-19 to Sep-20) post-CSFO year. Moreover, one-way ANOVA and post-hoc Tukey test were performed to detect differences between reference conditions and the first (Dec-18, Mar-19 and Jul-19) and the second (Dec-19, Feb-20 and Jul-20) post-CSFO year at reach scale. As reference values we assumed the metrics recorded in the study reach before the first documented CSFO, occurred in 200620. Related sampling took place on Nov-05, Mar-06 and Aug-06 (Supplementary Table 1).
Linear relationships between metrics of the benthic macroinvertebrate community and fine sediment deposition per unit area were tested by Pearson product-moment correlation test. Moreover, the correlation between the values of spatial beta-diversity (i.e., total pairwise dissimilarities between mesohabitats68) and the values of the difference in average fine sediment deposition between mesohabitats was assessed.
Statistical analyses were performed using XLSTAT 2014 and R 4.4.1 (vegan package69) software.
Results
Fine sediment deposits
Temporal trajectories of fine sediment deposition were similar, on average, between the three mesohabitats (Fig. 3), showing a significant decreasing trend at reach scale after the CSFO (Mann–Kendall test: S = -21, p = 0.037) (see Supplementary Note 2).

Time series of fine sediment deposition (three points average and corresponding standard deviation), periphyton biomass (measured as chlorophyll a content), and metrics of the benthic macroinvertebrate community (including the Siltation Index for LoTic EcoSystems—SILTES), detected in each mesohabitat before (i.e., five days before the CSFO starting date in September 2018) and after the CSFO (the days since the end of the CSFO are reported in brackets). The average trajectory at the reach scale is indicated by a line.
Before the CSFO (i.e., five days before, Sep-18), the reach-averaged mass of fine sediment per unit area was about 100 g m−2, and mesohabitat averages were very close each other (i.e., from 90 to 110 g m−2). The point values varied between few tens to two hundreds g m−2 (i.e., roughly spanning one order of magnitude).
After the CSFO (i.e., 17 days after, Oct-18), the reach-averaged mass of fine sediment per unit area increased to about 1,200 g m−2, i.e., more than one order of magnitude larger than before. Mesohabitat averages were comparable, though displaying a certain difference, ranging from 700 g m−2 in the riffle to 1,800 g m−2 in the pool. The point values varied between three hundreds to almost three thousands g m−2 (i.e., as for the pre-CSFO sampling, roughly spanning one order of magnitude, but more than ten times larger).
The amount of fine sediment settled on/into the superficial substrate of the permanently wetted streambed decreased in the following months, by a rather irregular pattern. About 13–15 months after the CSFO (i.e., 402 and 444 days after, Oct-19 and Dec-19), the deposits approached comparable values as in the pre-CSFO sampling. This was observed in terms of reach-average (120 and 80 g m−2, respectively), mesohabitat averages (from 110 to 130 g m−2 in Oct-19, and from 60 to 100 g m−2 in Dec-19), as well as point values (15–230 g m−2 in Oct-19, and 40–160 g m−2 in Dec-19). However, the 2020 samples evidenced a certain increase, mainly in the pool, where fine sediment mass per unit area was on average 2–3 times larger than before the CSFO.
The spatial distribution of the deposits was rather heterogeneous, at both reach and mesohabitat scale. Maximum values, both at mesohabitat scale and at single points, were generally (but not always) detected in the pool, while the minimum values were measured in the riffle.
At the reach scale, CV approximatively ranged between 50 and 150%. Interestingly, we observed comparable CVs before and after the CSFO (60 and 70% in Sep-18 and Oct-18, respectively), followed by increased CVs (110 and 150% in Mar-19 and Jul-19, respectively). When in Oct-19 and Dec-19 pre-CSFO levels of fine sediment deposition were recovered, the CV decreased as well (70 and 60%, respectively). Similarly, CVs at mesohabitat scale ranged from 20–40% to 120–140%, and the result was consistent when the adopted methodology was applied in triplicate within the same mesohabitat (Supplementary Note 3).
Periphyton
The periphyton amount, expressed as chlorophyll-a content, varied over time, but its pattern was more influenced by seasonality than by the sediment pulse in all mesohabitats (Fig. 3). In fact, no significant differences were detected between mesohabitats and between the first and the second post-CSFO year (two-way ANOVA and Tukey test, p > 0.05). On the one hand, a reduction of 20–50% was detected in the first post-CSFO sample; moreover, the absolute minima (1.6–3 mg m−2) were recorded on this sampling date. However, relevant increases (up to about 50 mg m−2 as reach average) were observed in the late autumn (December) and winter (February–March) samples, both the first and the second post-CSFO year. Specifically, during the collection of these samples, we ascertained the presence of the macroalga Hydrurus foetidus besides the biofilm of microalgae on some cobbles. Maximum and minimum periphyton amount was generally detected in the riffle and pool, respectively. At the reach scale, CV ranged from 3% in Sep-19 to 76% in Feb-20. The former value was even lower than the corresponding CV calculated for the riffle area (9%) to assess the intra-mesohabitat variability (see Supplementary Note 3).
Benthic macroinvertebrates
Gamma-diversity in the study reach amounted to 29 taxa: 26 insects, 2 oligochaetes and 1 hydrachnidia (see the whole taxa list in Supplementary Table 3). On average, the total richness was 14 (± 4), 15 (± 4) and 16 (± 4), and the total density 843 (± 1288), 950 (± 1167), 1518 (± 2225) ind. m−2 in the step-pool, pool, and riffle, respectively. At the reach scale, the most abundant taxa were the Diptera Chironomidae (22%), the Ephemeroptera Baetis (19%), the Trichoptera Limnephilidae (17%), the Ephemeroptera Rhitrogena (12%), the Plecoptera Leuctra (8%), and the Diptera Simuliidae (4%). Most EPT taxa displayed higher density in the riffle, while Limnephilidae, Lumbricidae and the Diptera Limoniidae and Athericidae were more abundant in the pool.
Two-way PERMANOVA and NMDS ordination (Fig. 4) evidenced significant variation in taxonomic composition of macroinvertebrate communities between sampling occasions (F10,33 = 4.15; p = 0.0001), but not between mesohabitats (F2,33 = 1.35; p = 0.139). The statistically non-significant results of PERMDISP (sampling occasions: F10,33 = 0.75; p = 0.676; mesohabitat: F2,33 = 0.58; p = 0.569) indicated location but not dispersion effects. Summer and early autumn samples (i.e., Jul., Sep. and Oct.) were mainly positioned on the right side of the panel. Specifically, September samples, including the pre-CSFO ones (Sep-18), were very close each other as well as to July samples, while October samples, including the first post-CSFO ones (Oct-18), were separated and positioned on the bottom right of the figure. In contrast, late autumn and winter samples (i.e., Dec., Feb. and Mar.) were located on the left side of the panel and separated, depending on the year.

Non-Metric Multidimensional Scaling (NMDS) of the composition of benthic macroinvertebrate assemblages detected in the three mesohabitats and at reach scale, before (i.e., Sept-18) and after (i.e., ten following samplings) the CSFO.
As for the PERMANOVA, the two-way ANOVA did not show significant (p > 0.05) differences of all the considered metrics between mesohabitats, but it evidenced significant (p < 0.05) differences between the first and the second post-CSFO year (Supplementary Table 4). In the three monitored mesohabitats, the temporal patterns of all the metrics (Fig. 3 and Supplementary Table 1) were similar. A general drop characterized the post-CSFO sample, and the less accentuated contraction took place in the riffle. Minimum values were indeed detected in the pool and/or in the step-pool, depending on the metric. Overall, metrics related to density (i.e., total density and ECOgA) were more affected than metrics based on richness (i.e., total taxon richness and EPT richness), with corresponding decreases of 66–96% and 16–56%, respectively. Substantial recovery to pre-CSFO standard was detected 73–181 days after the CSFO (sampling of Dec-18 and Mar-19, respectively), with the exception of richness metrics in the riffle, which fully recovered almost one year later (444 days after the CSFO, sampling of Dec-19). The relative abundance of EPT (% EPT) decreased in the first post-CSFO sample, even if its minimum was reached later on in the riffle and step-pool, and, in these two mesohabitats, also the recovery time was longer (Jul-19 for the riffle and Oct-19 for the step-pool). Among EPT taxa, Plecoptera and Ephemeroptera taxa generally displayed different trends than Trichoptera families: the former (with the exception of Baetis) were poorly represented during the first year after the CSFO, while the latter showed a more stable presence over the whole study period, as well as Diptera families. However, regardless the insect order, a considerable increase of density of most of the taxa detected in the study reach was recorded in the second post-CSFO winter (Dec-19 and Feb-20), differently from the first post-CSFO winter (Dec-18 and Mar-19). This occurrence influenced the pattern of the SILTES index, approaching its maximum in Feb-20. Similarly to the other metrics, this index clearly detected the impairment of the macroinvertebrate assemblages in the first post-CSFO sample, scoring 0 in the pool and in the step-pool. In these two mesohabitats, SILTES recovered to pre-CSFO standard in Mar-19 (181 days after the CSFO) even though, mainly for the step-pool, further drops of the index occurred also later on, until Dec-19. In the riffle, SILTES was stably below the half of the pre-CSFO standard until this month, reflecting the trend of the richness metrics composing the index.
At the reach scale (i.e., considering the average values of the three mesohabitats) a full recovery of most of the metrics occurred in Mar-19, i.e., 181 days after the CSFO. Considering all the metrics, the benthic macroinvertebrate community during the second post-CSFO year was not statistically different from the reference conditions (Fig. 5), displaying comparable seasonal pattern (Supplementary Table 1). In contrast, in the first post-CSFO year, both total and EPT richness were significantly lower than in the reference year. The average values of all the remaining metrics were also lower than reference, but differences were not statistically significant due to the high standard deviation (connected to high seasonal variability) (Fig. 5).

Comparison of metrics describing the benthic macroinvertebrate community at the study reach between the reference conditions and the first and second post-CSFO year. Averages plus standard deviations are reported for each metric. Lowercase letters (a,b) indicate significant differences between years (Tukey test, p < 0.05). Unit of measurement: individuals m−2 for density and ECOgA density, and mg dry weight m−2 for the biomass of each trophic guild.
Considering the whole dataset, significant negative correlation was detected between fine sediment deposition per unit area and total richness (r = − 0.42, p = 0.016), EPT richness (r = − 0.50, p = 0.003), SILTES (r = − 0.35, p = 0.046), and relative abundance of EPT (r = − 0.50, p = 0.003). Moreover, the difference in the average fine sediment deposition per unit area between mesohabitats was significantly correlated to their beta-diversity (r = 0.45, p = 0.008) (see Supplementary Note 4).
Fishes
Two fish species were sampled in the study reach, brown trout and European bullhead Cottus gobio L. In the first post-CSFO sample (< 1 month after the CSFO), trout density remained the same as before the CSFO (1380 individuals ha−1) and bullhead density even increased (from 1280 ind. ha−1 in Aug-18 to 1620 ind. ha−1 in Oct-18). In the second post-CSFO sample (Jan-20, i.e., > 1 year after the CSFO), the density of both species decreased (trout 950 ind. ha−1 and bullhead 630 ind. ha−1). Fish biomass followed a similar pattern, with the exception of the trout biomass which slightly decreased in the first post-CSFO sample (trout 59, 52, 39 kg ha−1 and bullhead 30, 37, 14 kg ha−1 in Aug-18, Oct-18 and Jan-20, respectively).
Food web
Before the CSFO (Aug-18/Sep-18, Fig. 6) the study reach was markedly heterotrophic, being predators the main component of the food web in terms of carbon mass. Periphyton mass was 1.5 times higher than grazers mass, and the proportions of benthic macroinvertebrate carbon among the four macroinvertebrate trophic guilds were 25%, 6%, 40% and 29% for grazers, shredders, collectors and predators, respectively. Fishes had the highest carbon mass; in particular, the trout mass was almost twice as the bullhead one, and three times larger than the benthic macroinvertebrate mass.

Scheme of the stream food web showing the mass of organic carbon (g ha−1) detected for the different biological components once before and twice after the CSFO (less than one month and more than one year later). Coarse (CPOM), fine (FPOM) and dissolved (DOM) organic matter are indicated to complete the food web scheme.
Except bullhead, after the CSFO (< 1 month later, Oct-18, Fig. 6) the other components of the food web decreased in terms of biomass, and the ratios among them changed. The ratio between periphyton and grazers carbon mass increased up to 7.7 times. Predators became the dominant component among macroinvertebrates (45%), followed by collectors (33%), shredders (11%) and grazers (11%). Since the mass of fishes remained almost unvaried, their prevalence compared to the other components of the food web increased by far.
More than one year after the CSFO (Jan-20/Feb-20, Fig. 6), the distribution of carbon in the food web was completely different. Periphyton became the dominant component, followed by grazers with comparable carbon mass. Consequently, the proportions of the four trophic guilds of benthic macroinvertebrates changed as follows: grazers (49%), collectors (21%), shredders (17%), and predators (13%). Although the higher availability of prey, the mass of both fish species decreased. The temporal pattern of the functional composition of benthic macroinvertebrate assemblages evidenced a relevant change during the second post-CSFO winter (Dec-19 and Feb-20), with a marked increase of grazers and of the autotrophy of the system compared to all the other sampling occasions (see Supplementary Note 5 and Supplementary Table 2). This increase of grazers in winter was also detected during the reference year (Supplementary Table 2) and, in general, significant differences of the average biomass of the trophic guilds (grazers and collectors) compared to the reference year were detected only for the first post-CSFO year (Fig. 5).
Discussion
CSFO effects on the streambed substrate
Fine sediment evacuated from reservoirs by flushing can determine a wide range of morphological effects on the downstream river systems, up to severe alterations at the reach scale (e.g., thalweg raising70) and at the mesohabitat scale (e.g., pool filling71). Changes in riverbed morphology of such magnitude can have complex feedbacks on subsequent fine sediment dynamics35. Less macroscopic consequences as fine sediment infiltration into formerly coarser riverbed substrata (clogging) can interfere with hyporheic exchanges, inducing relevant biotic effects72,73. However, the quantification of the mass of fine sediment deposited after flushing operations, its spatial distribution and temporal dynamics have been poorly documented so far74, probably as a consequence of the intrinsic difficulties of this kind of field investigation75,76. In particular, the hydro-morphological patchiness of upland stream environments determines complex and highly variable depositional patterns, making assessment efforts especially challenging. Accordingly, coefficients of variation of sediment deposition per unit area exceeded in some instances 100%, at both reach and mesohabitat scale. Moreover, the CVs increase in some post-CSFO sampling suggested selective removal processes, strongly conditioned by local (microhabitat) conditions. In this perspective, our monitoring of fine sediment coverage after a CSFO, during a time span of two years (temporal component) and considering three different mesohabitats (spatial component), provide new insights into sustainable management of reservoirs siltation.
As expected, after the CSFO, the fine sediment content in the three monitored mesohabitats increased. Specifically, the average mass of fine sediment per unit area recorded few days after the CSFO ranged from 0.6 to 1.7 kg m−2, with an overall increase at the reach scale of approximately one order of magnitude compared to the pre-CSFO standard. Comparable results were obtained after the spot monitoring of the CSFO performed in 2008 from the same hydropower reservoir (1–2.5 kg m−2)20, as well as after a CSFO performed in 2011 from another reservoir in the same area (0.5–1 kg m−2)13. It should be underlined that the mentioned flushing events, investigated streams, and related measurement techniques shared basic similarity. In fact, the flushed sediment was predominantly silt, the study reaches had relatively high channel slope (> 0.01), and only the streambed wetted at baseflow was sampled by resuspension. Under these circumstances, mountain streams have considerable transport capacity, and deposition is typically a negligible fraction in comparison to sediment flux. In contrast, much larger deposition per unit area, up to two/three orders of magnitude (i.e., 30–300 kg m−2), was measured in comparable settings, when the flushed sediment was coarser, predominantly in the sand range15,77.
As expected considering increased water depth and slower flow velocity, major streambed alteration due to deposition of fine sediment after the CSFO was detected in the pool. However, the time pattern of fine sediment deposition in the surveyed mesohabitats was similar: although with some oscillations, the values decreased over the study period, recovering to pre-CSFO standard after approximately one year. Further increases at the end of the study period could be related to possible upstream depositional areas, acting as source of fine sediment for the downstream reaches, activated concurrently to rainfall events and subsequent increased runoff in the Roasco Stream. Specifically, daily rainfall depths gauged in a nearby station led us to exclude massive flooding over the study period, though on a few occasions daily rainfall depth was between 60 and 80 mm (fall 2018 and summer 2020).
CSFO effects on benthic organisms
As quantified by comparing pre and post-flushing samples, both periphyton and benthic macroinvertebrates were negatively affected by the CSFO in the short term in all three mesohabitats (i.e., the minimum values of all metrics were detected immediately after the CSFO). The minor deposition of fine sediment detected in the riffle could explain the minor contraction of the benthic macroinvertebrate community in terms of density and richness observed in this mesohabitat. However, in the pool and in the step-pool all the metrics considered in this study recovered within five months, while the total taxon richness and the richness of taxa belonging to EPT orders in the riffle required more than one year to regain pre-CSFO standard. This might be justified by the higher richness characterizing this mesohabitat before the sediment pulse. In fact, riffles are usually the richest mesohabitats: composed of a mix of coarse particles subjected to limited fine sediment deposition, they provide the most favorable habitat to several reophilous taxa78,79,80, mostly belonging to EPT orders and included in the ECOgA, which are sensitive to sediment pressure and thus need longer time for recolonization after sediment disturbance19,20. These results contradict our expectation, as we hypothesized a major contraction of the riffle assemblage due to its higher biodiversity, and that the hydro-morphologic characteristics of the riffle, supporting efficient washing-out of fine sediment from the streambed, could have locally speeded up recolonization processes.
Seasonality plays an important role when assessing the CSFO impact over benthic assemblages in upland streams, deserving careful attention. In fact, in regulated residual-flow reaches of Alpine streams, these assemblages usually display maximum density in winter, followed by a contraction during summer50,81. In our investigation, this regular pattern seemed to superimpose to that forced by the sediment pulse. However, the metrics of benthic macroinvertebrate community were significantly higher in the second post-CSFO year than in the first one, approaching the reference standard and thus suggesting a CSFO impact over the medium term. The similarity in the composition and metric values between summer samples collected immediately, one and two years after the CSFO highlighted the selected flushing period as the best choice to minimize this impact.
All the mentioned differences in the impact and recovery patterns of the benthic macroinvertebrate assemblages were detected by the SILTES index, confirming a reliable biomonitoring tool for assessing the ecological impact of CSFOs in alpine streams33,59.
The detailed analysis of the macroinvertebrate community structure, also in terms of beta-diversity (see Supplementary Note 4), revealed that increased difference in riverbed sedimentation between mesohabitats corresponded to higher difference in the taxonomic composition between mesohabitats (spatial beta-diversity). However, the lack of significant differences led us to pool the data from different mesohabitats in a single dataset, thus finding significant inverse relationships at the reach scale between fine sediment deposition per unit area and some metrics (sediment-sensitive metrics), basically confirming the results of previous studies32,58,82.
CSFO effects on fish
Despite the drop detected for the lower trophic levels, no lethal impact on fishes was measured in the short term. However, the temporal resolution of fish sampling in this study is not adequate to assess if the contraction of both trout and bullhead populations observed in the second post-CSFO winter could be related to long-term effects of the CSFO. A previous study in the same stream20 suggested that contraction of fish density might be explained by an overall reduction of the carrying capacity of the environment after CSFOs, inducing reduction of suitable habitat and low availability of food. Nevertheless, our results of riverbed alteration and abundance of benthic organisms seem to exclude this possibility. In contrast, possible sublethal effects such as gill or other tissues abrasion can slowly reduce the fitness of fishes, favoring parasite, viral and bacterial infections and thus decreasing the rate of survival26.
The permitted sediment load during the CSFO was evaluated by the Newcombe & Jensen49 model, that confirmed an effective tool to providing preliminary estimate of the short-term impact of CSFOs on fish species, as demonstrated in several studies11,16,83. Despite its simplicity, the model was also applied in eco-hydraulic modeling studies focused on a detailed quantification of the effects of increased SSC during CSFOs over fish assemblages12,84,85. Moreover, Cattanèo et al.18 recently compared the impact over fish assemblage of a controlled partially-drawdown flushing of the Verbois Reservoir to a previous uncontrolled empty flushing operation. When the SSC of the evacuated waters was controlled according to the mentioned benchmark, the impact on fish was mostly reduced to behavioral impairment. Specifically, the CSFO schedule included a SSC limit over the entire operation (5 g L−1), as in this study, and two additional limits over shorter durations (10 and 15 g L−1 for 6 and 0.5 h, respectively). Revised versions of the Newcombe & Jensen49 model have been recently provided for salmonids86 and a cyprinid species87. In particular, Courtice et al.86 confirmed that duration of exposure should be considered as well as SSC in the regulatory guidelines aimed to reduce the environmental risk of suspended sediment. Moreover, they suggested the adoption of a threshold on suspended sediment dose, computed as the product of SSC and duration of exposure, especially for temporary and scheduled releases, such as CSFOs.
CSFO effects on stream food web
Interrelation between components of community biomass supports proper understanding of the trophic structure of aquatic communities, and changes in the biomass of community components can indicate perturbations to the ecosystem88. Therefore, analyzing the biomass of the main biological components in the Roasco Stream food web could reveal the effects of CSFOs at the ecosystem level. Although our analysis was limited to three sampling occasions and could be biased by the incomplete description of the system (including fishing-related practices), we clearly detected a heterotrophic system, dominated by predators, both in the pre and in the first post-CSFO sampling, even if in the latter case an evident grazers reduction was observed. More than one year later, a completely different picture was obtained, characterized by the dominance of periphyton and grazers.
Previous studies based on the River Continuum Concept89,90 evidenced that, in small forested headwaters, most carbon enters the stream as terrestrial food sources91,92. In particular, the results of a field experiment by Nakano et al.93 revealed that terrestrial arthropod inputs from riparian forest canopies are a primary factor controlling cascading trophic interactions between predatory fish, herbivorous aquatic arthropods, and benthic periphyton in headwater stream ecosystems. More generally, transfers of energy and biomass from more productive donor systems are often prominent for sustaining biotic communities in less productive recipient systems. However, even in headwaters where terrestrial inputs dominate, primary consumers evidence the preferential assimilation of high-quality periphyton over low-quality leaves94,95. This suggests that the short-term reduction of periphyton due to the CSFO could have been a co-occurring factor determining grazers reduction and temporary unbalance of the stream food web. Accordingly, the periphyton increase detected in the second post-CSFO winter favored grazers taxa which increased in density and became the dominant functional group among benthic macroinvertebrates. In these terms, our winter samplings depict a picture markedly different from what expected96, in particular the evident increase of the autotrophy of the system. Results on the functional composition of the benthic macroinvertebrate community showed that this increase was not recorded in the first post-CSFO winter, thus evidencing a medium-term impairment of the whole ecosystem functioning after the sediment pulse. This finding was also supported by the significant inverse relationship between the degree of autotrophy of the system, estimated through the ratio between grazers and the sum of shredders and collectors, and the amount of fine sediment per unit area (Supplementary Note 5). In fact, deposition of fine sediment can prevent attachment to substrate and smother periphyton, thus directly altering the physical composition of periphyton and diminishing its quantity and quality as a food source for benthic macroinvertebrates, and, in turn, for fishes25,26. On the other hand, the increase in abundance of macroinvertebrates could have also been caused by the decrease of fish density, and thus of the top-down control of macroinvertebrate populations by fish predation, mainly by brown trout97.
Conclusions
Detailed integrative assessment of the impact of CSFOs on the downstream abiotic and biotic (on various trophic levels) components are so far limited to only few studies22. Related field work is particularly challenging in mountain streams, where spatial and temporal variability of sediment deposition is especially complex35,74.
In this context, the results of this study confirm the major findings of previous works on the ecological impact of CSFOs11,18,19,20,21,33,48,59 and provide further information for improving the management and monitoring of CSFOs, in the investigated and in comparable settings. Specifically:
-
(i)
Overall, the standard protocol adopted for the CSFO (i.e., two weeks of duration in summer, with average SSC constrained below 4 g L−1 over the whole operation) provides an acceptable balance among technical, economic and ecological issues.
-
(ii)
If the flushed sediment is mainly silt, in high-gradient streams such as the Roasco, a moderate and patchy deposition can be expected after the sediment pulse, inducing a moderate ecological impact limited to the medium term. Full recovery of the pre-CSFO standard in terms of fine sediment content of the streambed substrate can be expected within the following year. A similar recovery time can be expected for the biological community, particularly if, as in the study stream, an undisturbed tributary can act as recolonization source. Therefore, more severe impact and longer recovery time may occur in stream reaches not supplied by substantial tributary flow and closer to the flushed reservoir.
-
(iii)
Detailed monitoring of different mesohabitats evidenced a certain variability in the abiotic (fine sediment deposition per unit area) and biotic response to the CSFO. For instance, if the assessment would have focused only on the step-pool, a larger short-term contraction of benthic macroinvertebrate assemblages and a faster recovery would have been estimated compared to the riffle. In this perspective, standard monitoring restricted to a single hydro-morphologic unit (mostly riffle) could be integrated by detailed mesohabitat monitoring when the hydro-morphologic characteristics of the receiving stream and the grain-size of the flushed sediment lead to predict significant difference of depositional patterns.
-
(iv)
The three biological groups investigated in this study responded differently to the CSFO, and their integrated analysis suggests possible effects on the whole ecosystem functioning. Future research is required to improve our comprehension of the effects of CSFOs (and, more generally, of sediment pulses) over stream food webs, possibly increasing temporal resolution, including further components (e.g., POM and terrestrial arthropods), and measuring stream metabolism in terms of respiration and primary productivity.
-
(v)
Our study deals with a managed environment. In particular, due to hydropower development, the streamflow is significantly lower than the natural flow, and has reduced seasonal variability. This flow pattern increases the persistence of fine sediment deposits, likely extending recovery times. Though the release of clean water (flushing flow) is commonly implemented after CSFOs as a mitigation measure, the subject is poorly documented so far73. Specifically, proper quantification of flushing flow magnitude, in light of clearly designed environmental objectives and related cost–benefit analyses could support a more informed and accepted diffusion of the practice, supporting the transition from reservoir desilting to a more sustainable management of the hydro-sedimentary regime in regulated rivers12. Similarly, sport fishing and related restocking of salmonids usually bias estimates of CSFOs effects over fish assemblages. Focusing increasing attention on bullhead or further species, deserving conservational interest but not subjected to this kind of dynamics, could improve the reliability of these results.
Data availability
Main data are provided within the manuscript and supplementary material file. Further information is available from the corresponding author upon reasonable request.
References
Morris, G. L. Classification of management alternatives to combat reservoir sedimentation. Water https://doi.org/10.3390/w12030861 (2020).
Tullos, D., Nelson, P. A., Hotchkiss, R. H. & Wegner, D. Sediment mismanagement puts reservoirs and ecosystems at risk. Eos https://doi.org/10.1029/2021EO157145 (2022).
Annandale, G. W., Morris, G. L. & Pravin, K. Extending the Life of Reservoirs, Sustainable Sediment Management for Dams and Run-of-River Hydropower (International Bank for Reconstruction and Development/The World Bank, 2016).
Hauer, C. et al. State of the art, shortcomings and future challenges for a sustainable sediment management in hydropower: A review. Renew. Sust. Energ. Rev. 98, 40–55. https://doi.org/10.1016/j.rser.2018.08.031 (2018).
Randle, T. J. et al. Sustaining United States reservoir storage capacity: Need for a new paradigm. J. Hydrol. 602, 126686. https://doi.org/10.1016/j.jhydrol.2021.126686 (2021).
Wu, X., Shen, X., Wei, C., Xie, X. & Li, J. Reservoir operation sequence-and equity principle-based multi-objective ecological operation of reservoir group: A case study in a basin of northeast china. Sustainability 14(10), 6150. https://doi.org/10.3390/su14106150 (2022).
Wentworth, C. K. A scale of grade and class terms for clastic sediments. J. Geol. 30(5), 377–392. https://doi.org/10.1086/622910 (1922).
Morris, G. L. & Fan, J. Reservoir Sedimentation Handbook: Design and Management of Dams, Reservoirs, and Watersheds for Sustainable Use (McGraw Hill Professional, 1998).
Shelley, J., Hotchkiss, R. H., Boyd, P. & Gibson, S. Discharging sediment downstream: Case studies in cost effective, environmentally acceptable reservoir sediment management in the United States. J. Water Res. Plan. Man. 148(2), 05021028. https://doi.org/10.1061/(ASCE)WR.1943-5452.0001494 (2022).
Petts, G. E. & Gurnell, A. M. Dams and geomorphology: Research progress and future directions. Geomorphology 71, 27–47. https://doi.org/10.1016/j.geomorph.2004.02.015 (2005).
Espa, P. et al. Tackling reservoir siltation by controlled sediment flushing: Impact on downstream fauna and related management issues. PLoS ONE 14, e0218822. https://doi.org/10.1371/journal.pone.0218822 (2019).
Panthi, M. et al. Effects of sediment flushing operations versus natural floods on Chinook salmon survival. Sci. Rep-UK 12(1), 15354. https://doi.org/10.1038/s41598-022-19294-2 (2022).
Espa, P., Brignoli, M. L., Crosa, G., Gentili, G. & Quadroni, S. Controlled sediment flushing at the Cancano Reservoir (Italian Alps): Management of the operation and downstream environmental impact. J. Environ. Manage. 182, 1–12. https://doi.org/10.1016/j.jenvman.2016.07.021 (2016).
Petkovšek, G., Roca, M. & Kitamura, Y. Sediment flushing from reservoirs: A review. Dams Reserv. 30(1), 12–21. https://doi.org/10.1680/jdare.20.00005 (2020).
Quadroni, S. et al. Effects of sediment flushing from a small Alpine reservoir on downstream aquatic fauna. Ecohydrology 9, 1276–1288. https://doi.org/10.1002/eco.1725 (2016).
Grimardias, D., Guillard, J. & Cattanéo, F. Drawdown flushing of a hydroelectric reservoir on the Rhone River: Impacts on the fish community and implications for the sediment management. J. Environ. Manage. 197, 239–249. https://doi.org/10.1016/j.jenvman.2017.03.096 (2017).
Espa, P., Crosa, G., Gentili, G., Quadroni, S. & Petts, G. Downstream ecological impacts of controlled sediment flushing in an Alpine valley river: A case study. River Res. Appl. 31(8), 931–942. https://doi.org/10.1002/rra.2788 (2015).
Cattanéo, F., Guillard, J., Diouf, S., O’Rourke, J. & Grimardias, D. Mitigation of ecological impacts on fish of large reservoir sediment management through controlled flushing–The case of the Verbois dam (Rhône River, Switzerland). Sci. Total Environ. 756, 144053. https://doi.org/10.1016/j.scitotenv.2020.144053 (2021).
Folegot, S. et al. The effects of a sediment flushing on Alpine macroinvertebrate communities. Hydrobiologia https://doi.org/10.1007/s10750-021-04608-8 (2021).
Espa, P., Castelli, E., Crosa, G. & Gentili, G. Environmental effects of storage preservation practices: Controlled flushing of fine sediment from a small hydropower reservoir. Environ. Manage. 52, 261–276. https://doi.org/10.1007/s00267-013-0090-0 (2013).
Doretto, A. et al. Effectiveness of artificial floods for benthic community recovery after sediment flushing from a dam. Environ. Monit. Assess. 191, 88. https://doi.org/10.1007/s10661-019-7232-7 (2019).
Hauer, C. et al. Controlled reservoir drawdown—challenges for sediment management and integrative monitoring: An Austrian case study—Part A: Reach scale. Water 12(4), 1058. https://doi.org/10.3390/w12041058 (2020).
Salmaso, F., Espa, P., Crosa, G. & Quadroni, S. Impacts of fine sediment input on river macroinvertebrates: The role of the abiotic characteristics at mesohabitat scale. Hydrobiologia https://doi.org/10.1007/s10750-021-04632-8 (2021).
Wood, P. J. & Armitage, P. D. Biological effects of fine sediment in the lotic environment. Environ. Manage. 21, 203–217 (1997).
Izagirre, O., Serra, A., Guasch, H. & Elosegi, A. Effects of sediment deposition on periphytic biomass, photosynthetic activity and algal community structure. Sci. Total Environ. 407(21), 5694–5700. https://doi.org/10.1016/j.scitotenv.2009.06.049 (2009).
Kemp, P., Sear, D., Collins, A., Naden, P. & Jones, I. The impacts of fine sediment on riverine fish. Hydrol. Process. 25, 1800–1821. https://doi.org/10.1002/hyp.7940 (2011).
Jones, J. I. et al. The impact of fine sediment on macroinvertebrates. River Res. Appl. 28, 1055–1071. https://doi.org/10.1002/rra.1516 (2012).
Jones, J. I., Duerdoth, C. P., Collins, A. L., Naden, P. S. & Sear, D. A. Interactions between diatoms and fine sediment. Hydrol. Process. 28, 1226–1237. https://doi.org/10.1002/hyp.9671 (2014).
Sear, D. A. et al. Does fine sediment source as well as quantity affect salmonid embryo mortality and development?. Sci. Total Environ. 541, 957–968. https://doi.org/10.1016/j.scitotenv.2015.09.155 (2016).
Bisson, P. A., Montgomery, D. R. & Buffington, J. M. Valley segments, stream reaches, and channel units. In Methods in Stream Ecology Vol. 1 (eds Lamberti, G. A. & Hauer, F. R.) 21–47 (Academic Press, 2017).
Luce, J. J., Lapointe, M. F., Roy, A. G. & Ketterling, D. B. The effects of sand abrasion of a predominantly stable stream bed on periphyton biomass losses. Ecohydrology 6(4), 689–699. https://doi.org/10.1002/eco.1332 (2013).
Buendia, C., Gibbins, C. N., Vericat, D., Batalla, R. J. & Douglas, A. Detecting the structural and functional impacts of fine sediment on stream invertebrates. Ecol. Indic. 25, 184–196. https://doi.org/10.1016/j.ecolind.2012.09.027 (2013).
Doretto, A., Espa, P., Salmaso, F., Crosa, G. & Quadroni, S. Considering mesohabitat scale in ecological impact assessment of sediment flushing. Knowl. Manag. Aquat. Ecol. 423, 2. https://doi.org/10.1051/kmae/2021037 (2022).
Mathers, K. L., Doretto, A., Fenoglio, S., Hill, M. J. & Wood, P. J. Temporal effects of fine sediment deposition on benthic macroinvertebrate community structure, function and biodiversity likely reflects landscape setting. Sci. Total Environ. 829, 154612. https://doi.org/10.1016/j.scitotenv.2022.154612 (2022).
Antoine, G., Camenen, B., Jodeau, M., Némery, J. & Esteves, M. Downstream erosion and deposition dynamics of fine suspended sediments due to dam flushing. J. Hydrol. 585, 124763. https://doi.org/10.1016/j.jhydrol.2020.124763 (2020).
Lepage, H. et al. Impact of dam flushing operations on sediment dynamics and quality in the upper Rhône River, France. J. Environ. Manage. 255, 109886. https://doi.org/10.1016/j.jenvman.2019.109886 (2020).
Sumi, T., Kantoush, S., Esmaeili, T. & Ock, G. Reservoir sediment flushing and replenishment below dams: Insights from Japanese case studies. Gravel-Bed Rivers: Processes and Disasters 385–414 (Wiley, 2017).
Wang, H. W., Kondolf, M., Tullos, D. & Kuo, W. C. Sediment management in Taiwan’s reservoirs and barriers to implementation. Water 10(8), 1034. https://doi.org/10.3390/w10081034 (2018).
Nukazawa, K., Kajiwara, S., Saito, T. & Suzuki, Y. Preliminary assessment of the impacts of sediment sluicing events on stream insects in the Mimi River, Japan. Ecol. Eng. 145, 105726. https://doi.org/10.1016/j.ecoleng.2020.105726 (2020).
Esmaeili, T. et al. Numerical study of discharge adjustment effects on reservoir morphodynamics and flushing efficiency: An outlook for the Unazuki Reservoir, Japan. Water 13, 1624. https://doi.org/10.3390/w13121624 (2021).
Chen, L. et al. Multi-objective water-sediment optimal operation of cascade reservoirs in the Yellow River Basin. J. Hydrol. 609, 127744. https://doi.org/10.1016/j.jhydrol.2022.127744 (2022).
Shrestha, J. P., Pahlow, M. & Cochrane, T. A. Managing reservoir sedimentation through coordinated operation of a transboundary system of reservoirs in the Mekong. J. Hydrol. https://doi.org/10.1016/j.jhydrol.2022.127930 (2022).
Ren, S., Zhang, B., Wang, W. J., Yuan, Y. & Guo, C. Sedimentation and its response to management strategies of the Three Gorges Reservoir, Yangtze River, China. Catena 199, 105096. https://doi.org/10.1016/j.catena.2020.105096 (2021).
Zhang, B., Wu, B., Zhang, R., Ren, S. & Li, M. 3D numerical modelling of asynchronous propagation characteristics of flood and sediment peaks in Three Gorges Reservoir. J. Hydrol. 593, 125896. https://doi.org/10.1016/j.jhydrol.2020.125896 (2021).
WWAP - World Water Assessment Programme, The United Nations World Water Development Report 2018 (United Nations Educational, Scientific and Cultural Organization, 2018). www.unwater.org/publications/world-water-development-report-2018/
Burt, T., Boardman, J., Foster, I. & Howden, N. More rain, less soil: Long-term changes in rainfall intensity with climate change. Earth Surf. Proc. Land. 41, 563–566. https://doi.org/10.1002/esp.3868 (2016).
Maruffi, L., Stucchi, L., Casale, F. & Bocchiola, D. Soil erosion and sediment transport under climate change for Mera River, in Italian Alps of Valchiavenna. Sci. Total Environ. 806, 150651. https://doi.org/10.1016/j.scitotenv.2021.150651 (2022).
Crosa, G., Castelli, E., Gentili, G. & Espa, P. Effects of suspended sediments from reservoir flushing on fish and macroinvertebrates in an alpine stream. Aquat. Sci. 72, 85–95. https://doi.org/10.1007/s00027-009-0117-z (2010).
Newcombe, C. P. & Jensen, J. O. Channel suspended sediment and fisheries: A synthesis for quantitative assessment of risk and impact. N. Am. J. Fish. Manage. 16(4), 693–727. https://doi.org/10.1577/1548-8675(1996)016%3c0693:CSSAFA%3e2.3.CO;2 (1996).
Quadroni, S., Crosa, G., Gentili, G. & Espa, P. Response of stream benthic macroinvertebrates to current water management in Alpine catchments massively developed for hydropower. Sci. Total Environ. 609, 484–496. https://doi.org/10.1016/j.scitotenv.2017.07.099 (2017).
McNeil, W. F. & Ahnell, W. H. Success of Pink Salmon Spawning Relative to Size of Spawning Bed Materials 469 (US Fish and Wildlife Service Special Scientific Report Fisheries, 1964).
Rex, J. F. & Carmichael, N. B. Guidelines for Monitoring Fine Sediment Deposition in Streams (Resources Information Standards Committee, 2002).
APAT, IRSA-CNR. Metodi analitici per le acque - Indicatori biologici - 2090. Solidi (APAT Manuali e linee guida 29/2003, 2003).
APAT, IRSA-CNR. Metodi Analitici per le Acque - Indicatori Biologici - 9020. Determinazione della Clorofilla: Metodo Spettrofotometrico (APAT Manuali e linee guida 29/2003, 2003).
Campaioli, S., Ghetti, P. F., Minelli, A. & Ruffo, S. Manuale per il Riconoscimento dei Macroinvertebrati delle Acque dolci Italiane Vol 1 357 (Provincia Autonoma di Trento, 1994).
Campaioli, S., Ghetti, P. F., Minelli, A. & Ruffo, S. Manuale per il Riconoscimento dei Macroinvertebrati Delle acque Dolci Italiane Vol 2 127 (Provincia Autonoma di Trento, 1999).
Usseglio-Polatera, P., Bournaud, M., Richoux, P. & Tachet, H. Biological and ecological traits of benthic freshwater macroinvertebrates: Relationships and definition of groups with similar traits. Freshw. Biol. 43, 175–205. https://doi.org/10.1046/j.1365-2427.2000.00535.x (2000).
Doretto, A., Piano, E., Bona, F. & Fenoglio, S. How to assess the impact of fine sediments on the macroinvertebrate communities of alpine streams? A selection of the best metrics. Ecol. Indic. 84, 60–69. https://doi.org/10.1016/j.ecolind.2017.08.041 (2018).
Doretto, A. et al. Beta-diversity and stressor specific index reveal patterns of macroinvertebrate community response to sediment flushing. Ecol. Indic. 122, 107256. https://doi.org/10.1016/j.ecolind.2020.107256 (2021).
Fenoglio, S., Tierno de Figueroa, J. M., Doretto, A., Falasco, E. & Bona, F. Aquatic insects and benthic diatoms: A history of biotic relationships in freshwater ecosystems. Water 12(10), 2934. https://doi.org/10.3390/w12102934 (2020).
Schmidt-Kloiber, A. & Hering, D. www.freshwaterecology.info: An online tool that unifies, standardises and codifies more than 20,000 European freshwater organisms and their ecological preferences. Ecol. Indic. 53, 271–282. https://doi.org/10.1016/j.ecolind.2015.02.007 (2015).
Gosselain, V., Hamilton, P. B. & Descy, J. P. Estimating phytoplankton carbon from microscopic counts: An application for riverine systems. Hydrobiologia 438(1), 75–90. https://doi.org/10.1023/A:1004161928957 (2000).
Carr, G. M., Morin, A. & Chambers, P. A. Bacteria and algae in stream periphyton along a nutrient gradient. Freshw. Biol. 50(8), 1337–1350. https://doi.org/10.1111/j.1365-2427.2005.01401.x (2005).
Elliott, J. M., Lyle, A. A. & Campbell, R. N. B. A preliminary evaluation of migratory salmonids as vectors of organic carbon between marine and freshwater environments. Sci. Total Environ. 194, 219–223. https://doi.org/10.1016/S0048-9697(96)05366-1 (1997).
Salonen, K., Sarvala, J., Hakala, I. & Viljanen, M. L. The relation of energy and organic carbon in aquatic invertebrates 1. Limnol. Oceanogr. 21(5), 724–730. https://doi.org/10.4319/lo.1976.21.5.0724 (1976).
Benke, A. C., Huryn, A. D., Smock, L. A. & Wallace, J. B. Length-mass relationships for freshwater macroinvertebrates in North America with particular reference to the southeastern United States. J. N. Am. Benthol. Soc. 18(3), 308–343. https://doi.org/10.2307/1468447 (1999).
Hamed, K. H. & Rao, A. R. A modified Mann-Kendall trend test for autocorrelated data. J. Hydrol. 204(1–4), 182–196. https://doi.org/10.1016/S0022-1694(97)00125-X (1998).
Carvalho, J. C., Cardoso, P. & Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Glob. Ecol. Biogeogr. 21(7), 760–771. https://doi.org/10.1111/j.1466-8238.2011.00694.x (2012).
Oksanen, J. et al. Package ‘vegan’. Community Ecol. Package Version 2(9), 1–295 (2013).
Brandt, S. A. & Swenning, J. Sedimentological and geomorphological effects of reservoir flushing: The Cachí Reservoir, Costa Rica, 1996. Geogr. Ann. A 81(3), 391–407. https://doi.org/10.1111/j.0435-3676.1999.00069.x (1999).
Wohl, E. E. & Cenderelli, D. A. Sediment deposition and transport patterns following a reservoir sediment release. Water Resour. Res. 36(1), 319–333. https://doi.org/10.1029/1999WR900272 (2000).
Descloux, S., Datry, T. & Marmonier, P. Benthic and hyporheic invertebrate assemblages along a gradient of increasing streambed colmation by fine sediment. Aquat. Sci. 75, 493–507. https://doi.org/10.1007/s00027-013-0295-6 (2013).
Dubuis, R. & De Cesare, G. The clogging of riverbeds: A review of the physical processes. Earth-Sci. Rev. 239, 104374. https://doi.org/10.1016/j.earscirev.2023.104374 (2023).
Legout, C., Droppo, I. G., Coutaz, J., Bel, C. & Jodeau, M. Assessment of erosion and settling properties of fine sediments stored in cobble bed rivers: The Arc and Isère alpine rivers before and after reservoir flushing. Earth Surf. Proc. Land. 43(6), 1295–1309. https://doi.org/10.1002/esp.4314 (2018).
Hedrick, L. B., Anderson, J. T., Welsh, S. A. & Lin, L. S. Sedimentation in mountain streams: A review of methods of measurement. Nat. Res. 4(1), 29185. https://doi.org/10.4236/nr.2013.41011 (2013).
Thollet, F. et al. Long term high frequency sediment observatory in an alpine catchment: The Arc-Isère rivers, France. Hydrol. Process. 35(2), e14044. https://doi.org/10.1002/hyp.14044 (2021).
Brignoli, M. L., Espa, P. & Batalla, R. J. Sediment transport below a small alpine reservoir desilted by controlled flushing: Field assessment and one-dimensional numerical simulation. J. Soils Sedim. 17, 2187–2201. https://doi.org/10.1007/s11368-017-1661-0 (2017).
Gordon, N. D., McMahon, T. A., Finlayson, B. L., Gippel, C. J. & Nathan, R. J. Stream Hydrology: An Introduction for Ecologists (Wiley, 2004).
Barnes, J. B., Vaughan, I. P. & Ormerod, S. J. Reappraising the effects of habitat structure on river macroinvertebrates. Freshw. Biol. 58, 2154–2167. https://doi.org/10.1111/fwb.12198 (2013).
Herbst, D. B., Cooper, S. D., Medhurst, R. B., Wiseman, S. W. & Hunsaker, C. T. A comparison of the taxonomic and trait structure of macroinvertebrate communities between the riffles and pools of montane headwater streams. Hydrobiologia 820(1), 115–133. https://doi.org/10.1007/s10750-018-3646-4 (2018).
Quadroni, S., Salmaso, F., Gentili, G., Crosa, G. & Espa, P. Response of benthic macroinvertebrates to different hydropower off-stream diversion schemes. Ecohydrology 14(5), e2267. https://doi.org/10.1002/eco.2267 (2021).
Larsen, S., Vaughan, I. P. & Ormerod, S. J. Scale-dependent effects of fine sediments on temperate headwater invertebrates. Freshw. Biol. 54(1), 203–219. https://doi.org/10.1111/j.1365-2427.2008.02093.x (2009).
Xu, F. et al. Short informative title: Quantitative assessment of acute impacts of suspended sediment on carp in the Yellow River. River Res. Appl. 34(10), 1298–1303. https://doi.org/10.1002/rra.3375 (2018).
Tritthart, M., Haimann, M., Habersack, H. & Hauer, C. Spatio-temporal variability of suspended sediments in rivers and ecological implications of reservoir flushing operations. River Res. Appl. 35(7), 918–931. https://doi.org/10.1002/rra.3492 (2019).
Pisaturo, G. R., Folegot, S., Menapace, A. & Righetti, M. Modelling fish habitat influenced by sediment flushing operations from an Alpine reservoir. Ecol. Eng. 173, 106439. https://doi.org/10.1016/j.ecoleng.2021.106439 (2021).
Courtice, G., Bauer, B., Cahill, C., Naser, G. & Paul, A. A categorical assessment of dose-response dynamics for managing suspended sediment effects on salmonids. Sci. Total Environ. 807, 150844. https://doi.org/10.1016/j.scitotenv.2021.150844 (2022).
Li, X. et al. Experimental study on acute impacts of hyper-concentrated flow on Gymnocypris eckloni and implication for sediment management. River Res. Appl. https://doi.org/10.1002/rra.4104 (2023).
Fraley, K. M., Warburton, H. J., Jellyman, P. G., Kelly, D. & McIntosh, A. R. The influence of pastoral and native forest land cover, flooding disturbance, and stream size on the trophic ecology of New Zealand streams. Austral. Ecol. 46(5), 833–846. https://doi.org/10.1111/aec.13028 (2021).
Vannote, R. L., Minshall, G. W., Cummins, K. W., Sedell, J. R. & Cushing, C. E. The river continuum concept. Can. J. Fish. Aquat. Sci. 37(1), 130–137. https://doi.org/10.1139/f80-017 (1980).
Doretto, A., Piano, E. & Larson, C. E. The River Continuum Concept: Lessons from the past and perspectives for the future. Can. J. Fish. Aquat. Sci. 77(11), 1853–1864. https://doi.org/10.1139/cjfas-2020-0039 (2020).
Webster, J. R. & Meyer, J. L. Organic matter budgets for streams: A synthesis. J. N. Am. Benthol. Soc. 16, 141–161. https://doi.org/10.2307/1468247 (1997).
Wolff, L. L., Carniatto, N. & Hahn, N. S. Longitudinal use of feeding resources and distribution of fish trophic guilds in a coastal Atlantic stream, southern Brazil. Neotrop. Ichthyol. 11, 375–386. https://doi.org/10.1590/S1679-62252013005000005 (2013).
Nakano, S., Miyasaka, H. & Kuhara, N. Terrestrial–aquatic linkages: Riparian arthropod inputs alter trophic cascades in a stream food web. Ecology 80(7), 2435–2441. https://doi.org/10.1890/0012-9658(1999)080[2435:TALRAI]2.0.CO;2 (1999).
Kühmayer, T. et al. Preferential retention of algal carbon in benthic invertebrates: Stable isotope and fatty acid evidence from an outdoor flume experiment. Freshw. Biol. 65(7), 1200–1209. https://doi.org/10.1111/fwb.13492 (2020).
Guo, F. et al. Longitudinal variation in the nutritional quality of basal food sources and its effect on invertebrates and fish in subalpine rivers. J. Anim. Ecol. 90(11), 2678–2691. https://doi.org/10.1111/1365-2656.13574 (2021).
Thompson, R. M. & Townsend, C. R. The effect of seasonal variation on the community structure and food-web attributes of two streams: Implications for food-web science. Oikos https://doi.org/10.2307/3546998 (1999).
Huryn, A. D. Ecosystem-level evidence for top-down and bottom-up control of production in a grassland stream system. Oecologia 115(1), 173–183. https://doi.org/10.1007/s004420050505 (1998).
Acknowledgements
The authors thank Dr. Andrea Facchin, and Dr. Andrea Longhi for their field and lab work, and Dr. Gaetano Gentili for providing data on fishes and general information on the CSFO carried out in 2018.
Author information
Authors and Affiliations
Contributions
Silvia Quadroni and Paolo Espa conceptualized and planned the study, carried out the investigation and analyses, and wrote the manuscript. Silvia Quadroni created the graphs and illustrations. Livia Servanzi contributed to the field investigation and lab work. Giuseppe Crosa supervised the research. All authors contributed to the critical revision of the manuscript and gave their approval to the final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Quadroni, S., Servanzi, L., Crosa, G. et al. Two-year assessment of the effects of controlled sediment flushing on stream habitats and biota at reach scale. Sci Rep 14, 21048 (2024). https://doi.org/10.1038/s41598-024-72015-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-72015-9
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
