
Abstract
Failure in irrigation management of grapevines grown in the Brazilian semiarid region can affect bud fertility. Adequate irrigation, considering both the development of bunches in the current cycle and the formation of fertile buds for subsequent cycles, can bring significant advances to viticulture. Therefore, the objective of this research was to investigate the effect of different irrigation levels during flowering on the formation of buds and potential bunches of 'Arra 15' grapevine and its relationship with metabolic processes. A field experiment was carried out in a commercial vineyard in Petrolina, Pernambuco, Brazil, during the 2021 and 2022 seasons. The experiment was designed in randomized blocks with four replications and five irrigation levels (70; 85; 100; 115 and 130% of crop evapotranspiration – ETc) during three production cycles. The variables fertile bud, vegetative bud, dead bud, potential fertility of the basal, median, and apical regions of the branches, number of potential bunches, reducing sugar, total soluble sugar, net photosynthesis, stomatal conductance, transpiration, and relative chlorophyll index were evaluated. The 115% ETc irrigation level improved the number of fertile buds and number of potential bunches. Irrigation level above 115% ETc increased gas exchange and relative chlorophyll index, while 70% ETc increased leaf sugar content. The most appropriate irrigation strategy is the application of 115% ETc during the flowering stage, for the increase of fertile buds and potential bunches of the next cycle, without influencing the vine metabolism. Total soluble sugars are a promising indicator of water deficit during flowering and as an indicator of vegetative bud formation for the next cycle.
Similar content being viewed by others


Introduction
One of the major challenges for irrigated viticulture in the Brazilian semi-arid region is the reduced productivity due to inconsistent bud fertility. Bud fertility is a quantitative measure of the potential to produce fruit, or an indicator of the number of bunches that will be harvested in the next season1. Thus, fertile bud formation has great economic relevance, since the number of bunches harvested is the major factor determining grapevine yield.
It should be noted that the management practices adopted in the vineyard should be focused not only on the development of the bunches, but also on favoring the formation of fertile buds for subsequent cycles. The position of the bud on the shoot is one of the main factors affecting bud fertility. The tendency for fertility to increase from the middle portion of the stems is due to a higher accumulation of carbohydrates in this region2. The low fertility of the basal buds may also be influenced by the lower light incidence on these buds3.
In grapevine, the formation of fertile buds is the result of the differentiation of undifferentiated primordia into reproductive primordia. The formation of inflorescence primordia is the most sensitive stage in the development of the reproductive organs in grapevines4. Inflorescence primordia is influenced by a set of environmental and endogenous factors, such as hormonal balance, branch vigor, ambient temperature, light intensity, water availability, mineral nutrition, and crop management5. Among these, water availability deserves to be highlighted, mainly because it influences the bud formation6 and is involved in morphological, biochemical, and physiological processes in plants7.
Water stress during flowering in the previous season determines most of the variations in bud fertility in the next season6. Cabral et al.8 found that 25% of ETc is an appropriate strategy for ensuring yield, quality and vegetative vigor in Touriga Franca vines grown in Mediterranean regions. Williams et al.9 found that irrigation with 140% of ETc increases vegetative vigor and shading in the renewal zone.
Bud fertility can be affected by both soil and climate conditions and cropping system1. Thus, carrying out appropriate management practices for each cultivar in each region is essential. In this regard, the hypothesis of this research is that during the inflorescence primordia formation step, the reduction of the irrigation rate can interfere in metabolic events of the vine without impairing the differentiation of buds for the next cycle.
Deepening this field of knowledge may bring advances in viticulture management, contributing to the increase of yield over several cycles. In this perspective, the objective of this study was to investigate the effect of different irrigation rates during flowering on the formation of buds and potential bunches of 'Arra 15' grapevine and its relationship with metabolic processes.
Results
Table 1 contains the p-value of the F test for each source of variation and variables evaluated. There was a significant effect (p-value < 0.01) on irrigation rates for the variables fertile bud (FB), vegetative bud (VB), dead bud (DB), potential fertility of basal branch region (PFBB), middle branch region (PFMB), apical branch region (PFAB), number of potential bunches (NPB), reducing sugar content (RS), total soluble sugar (TSS), net photosynthesis (A), stomatal conductance (gs), and transpiration (E) and relative chlorophyll index (RCI). The interaction between water levels and cycle indicates that there are other climatic factors affecting these variables.
Percentage of buds
For the variables related to the process of bud differentiation by the presence or absence of inflorescence primordia, a quadratic response was observed for fertile buds (FB), with a maximum mean value of 86.8% achieved with the 115.8% ETc (Fig. 1A). For vegetative buds (VB), there was a linear response, with a higher mean value of 9.7% (Fig. 1B) and 9.1% for dead bud (DB) (Fig. 1C), reached, with the 70 and 130% ETc rates, respectively.

Means of the percentage of buds by presence or absence of inflorescence primordial in 'Arra15' grapevines grown under different irrigation rates in semi-arid conditions. (A) Fertile bud—FB. (B) Vegetative bud—VB. (C) Dead bud—DB. * and **Significant at 5% and 1% probability, respectively.
Potential fertility and number of potential bunches
For the variables related to bud potential fertility, quadratic responses were observed with maximum points of 89.64% for potential fertility of the basal branches -PFBB (Fig. 2A), 96.97% for potential fertility of the middle branches—PFMB (Fig. 2B) and 83.43% for potential fertility of the apical branches—PFAB (Fig. 2C), reached with the rates of 125.78% 113.35% and 113.39% ETc, respectively.

Means of potential fertility of 'Arra15' grapevines grown under different irrigation rates in semi-arid conditions. (A) Potential fertility of the basal branches—PFBB. (B) Potential fertility of the middle branches—PFMB. (C) Potential fertility of the apical branches—PFAB. * and **Significant at 5% and 1% probability, respectively.
Regarding the variable number of potential bunch (NPB), a quadratic response was observed with maximum points of 389.11 reached under condition of 116.82% ETc (Fig. 3).

Means of number of potential bunches (NPB) of 'Arra15' grapevines grown under different irrigation rates in semi-arid conditions. * and **Significant at 5% and 1% probability, respectively.
Physiological variables
In Fig. 4, the regression equation allowed estimating the effect of irrigation rates on leaf physiological variables. A decreasing linear response was observed as a function of irrigation rates, with the highest mean values of 307.08 and 4700.51 mg g–1 for RS and TSS, respectively, both reached with the 70% ETc rate.

Means of leaf reducing sugars of 'Arra15' grapevines grown under different irrigation rates in semi-arid condition. (A) Reducing sugar content—RS. (B) Total soluble sugar content—TSS. * and **Significant regression at 5% and 1% probability, respectively.
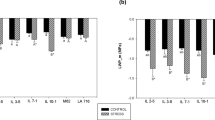
According to the regression equations for the variables related to gas exchange, an increasing linear response was observed, with the highest mean value of 17.32 µmol m–2 s–1 for net photosynthesis (Fig. 5A), 4.24 mmol m–2 s–1 for transpiration (Fig. 5B), and 0.16 mol m–2 s–1 for stomatal conductance (Fig. 5C), respectively, achieved with 130% ETc.

Means of leaf gas exchange in 'Arra15' grapevines grown under different irrigation rates in semi-arid conditions. (A) Net photosynthesis—A. (B) Stomatal conductance—gs. (C) Transpiration—E. * and **Significant at 5% and 1% probability, respectively.
It can be seen in Fig. 6 that the regression equation allowed estimating the effect of irrigation rates on the relative chlorophyll index (RCI), with an increasing linear response as a function of irrigation rates, with the highest average value of 46.42 RCI unit, reached with the irrigation rate of 130% ETc.

Mean relative chlorophyll index—(RCI) in leaves of 'Arra15' grapevines grown under different irrigation rates in semi-arid conditions. * and **Significant regression at 5% and 1% probability, respectively.
Canonical variables analysis
By canonical variables analysis (Fig. 7), the colored dots represent the irrigation rates (70; 85; 100; 115 and 130% ETc), while the arrows near and in the same direction demonstrate correlated variables (≥ ± 0.0). The first two canonical variables (DIM1 and DIM2) explained 81.4% of the total variation contained in the original dataset.

Biplot for Canonical Variables analysis in 'Arra 15' grapevine under different water regimes, studied as a function of the variables fertile buds (FB), vegetative buds (VB), dead buds (DB), potential fertility of basal branches (PFBB), potential fertility of middle branches (PFMB), potential fertility of apical branches (PFAB), number of potential bunches (NPB), reducing sugar (RS), total soluble sugar (TSS), relative chlorophyll index (CRI), net photosynthesis (A), stomatal conductance (gs), transpiration (E).
In Fig. 7, it is observed that the 115 and 130% of ETc rates are similar to each other, and positively favor the formation of FB, PFBB, PFMB, and PFAB, resulting in a higher NPB. The 115% and 130% ETc irrigation rate also exerted an influence on DB and RCI variables, which were inversely proportional to RS. The variables related to gas exchange were favored when they were close to the irrigation rate of 100% ETc. On the other hand, the variables VB and TSS were favored under 70% ETc condition.
Pearson correlation
According to the Biplot presented in Fig. 8, high, positive, and significant correlations (P < 0.01) were observed between FB × NPB (0.960), A × gs (0.990), A × E (0.921), and gs × E (0.933). The other variables are associated with different magnitudes, highlighting the positive and significant correlations (P < 0.01) between FB x A (0.706) and DB × RCI (0.868).

Biplot for Pearson's correlation analysis between the variables fertile bud (FB), vegetative bud (VB), dead bud (DB), potential fertility of basal branches (PFBB), middle branches (FPMB) and apical branches (PFAB), number of potential bunches (NPB), reducing sugar content (RS), total soluble sugar content (TSS), relative chlorophyll index (RCI), net photosynthesis (A), stomatal conductance (gs), and transpiration (E). Values shown in black indicate the overall correlation, while colored values indicate the correlation between irrigation managements. *, ** and ***Significant at 5, 1 and 0.1% probability by t test, respectively.
Discussion
However, it was expected that the treatment 130% ETc would have a greater influence on the percentage of fertile buds, considering the higher photosynthetic rate in this treatment (Fig. 8A), but this was not verified here. The decrease in sugars under 30% ETc condition may have influenced physiological disturbance, affecting the formation of inflorescence primordial and hence contributing to the incidence of DB by necrosis. It is worth noting that most of the soluble non-structural carbohydrate content of grapevine leaves consists of reducing sugars (glucose and fructose) and total soluble sugars (hexoses such as sucrose)10.
In grapevine, the process of differentiation of the vegetative bud into a fertile bud occurs at three well-defined stages: formation of the undifferentiated primordium, differentiation into an inflorescence primordium, and bloom formation5. The formation of the undifferentiated primordium and the differentiation into an inflorescence primordium occur during the growth of the branches from the previous cycle, lasting 45 to 60 days from sprouting, while bloom formation occurs shortly before and during the sprouting of the next cycle4.
In this regard, the results found here indicate that the irrigation rate applied at the formation of the inflorescence primordia in a cycle influences the number of fertile buds for the next cycle in 'Arra 15' grapevines. Guilpart et al.6 when evaluating 'Shiraz' and 'Aranel' vines grown in Mediterranean climate, found that irrigation with 60% ETc during flowering of the previous season determines most of the variations in bud fertility of the next season, supporting our findings.
During the period from 45 to 60 DAPP, the irrigation rate of 70% of ETc possibly caused an excessive reduction of water, disfavoring the process of bud differentiation, resulting in an increased percentage of vegetative buds in the following cycle. A contrasting pattern was found in 'Shiraz' and 'Aranel' grapes grown in Mediterranean climate conditions, where a mild water deficit (60% ETP) during flowering in the previous season led to an increased number of fertile buds in the subsequent season4.
Bud necrosis in grapevine is a physiological disorder, usually characterized by abortion of primary and secondary buds, and is associated with inadequate carbohydrate levels11. However, it is likely that excessive water availability may have an adverse effect on the formation of fertile buds, since the 130% ETc condition led to an increase in dead buds. Similarly, irrigation with 140% of ETc increased bud necrosis in 'Thompson Seedless' vines grown in California, as a consequence of increased vegetative vigor and shading in the renewal zone9.
Irrigation rates below 100% ETc, applied at the stage of inflorescence primordia formation of 'Arra 15' grapevine can reduce bud fertility in any position of fertile buds on the branch. According to Guilpart et al.6, water shortage during the floral differentiation process leads to decreases in bud fertility in the subsequent cycle, supporting the findings reported here.
Higher fertility in the middle branches when compared to the basal and apical region, regardless of the applied irrigation rate, may be related to genetic traits of the cultivar itself. In 'Arra 15' vines grown in a desert climate, there was also an increase in bud fertility from the basal buds to the middle buds and a gradual decrease towards the distal buds12.
However, the amount of water made available somehow influences the deviation of photoassimilates from branch growth to the development of inflorescence primordia4. Baeza et al.3 reported that a higher water availability in the grapevine canopy region forms a microclimate around the bud and may influence bud fertility. Meanwhile, Vilar et al.1 reported that the concentration of fertile buds in the more distal region of the branch denotes a higher vegetative activity.
The formation of inflorescence primordia in the buds during the previous cycle defines the potential number of bunches that the vine will produce in the subsequent cycle1. In this scenario, it is evident that the use of water supply from 100% ETc during the inflorescence primordia formation has a greater influence on the potential number of bunches per plant for the next crop cycle. This information is of considerable interest to the winegrower, as it allows him to estimate the production and pruning management for the next cycle.
The determination of the number of potential bunches is affected by water availability during the initial stages of vegetative bud differentiation into fertile ones from the previous cycle6. Thus, water supply above 100% ETc may be an appropriate strategy for the bud differentiation process in 'Arra15' grapevine, favoring the number of potential bunches per plant for the following cycle.
According to the canonical variables, the 115 and 130% ETc levels had a stronger influence on the bud differentiation process, since they favored the formation of FB, PFBB, PFMB, PFAB, and consequently the NPB. This means that maintaining high irrigation levels between 45 and 50 DAPP positively influences bud fertility in 'Arra 15' grapevines. According to Balint and Reynolds13, to achieve high bud fertility and productive potential for the next cycle, the vine needs high water demand during the differentiation of the inflorescence primordia, supporting the findings of this study.
The gradual reduction of the irrigation rate during the stage of inflorescence primordia formation, across all three crop cycles, causes an increase in RS and SST levels in 'Arra15' leaves. These results are similar to those obtained in Malbec vines grown in arid climates, where the lowest level of irrigation (25% ETo) showed a higher content of total soluble sugars than the vines under the highest level of irrigation (100% ETo)14.
Increasing sugars under irrigation level of 70% ETc is likely due to the reduction of translocation of these photoassimilates to the drains. Accumulation of free sugars in the leaves may be due to the disturbances caused by stressful conditions such as the reduction of water content in leaf tissues, reducing the translocation of sugars to other organs15.
Fahim et al.16 reported the synthesis of soluble sugars in the cytosol is probably prioritized to maintain the plant's metabolic activity, even when water deficit reduces the rate of CO2 assimilation. Meanwhile, Oliveira et al.15 reported that the accumulation of free sugars in the leaves with decreasing water content in the soil maintains cell turgor contributing to plant functioning. However, in order to confirm osmotic adjustment, assessing other physiological parameters that were not investigated in these studies is necessary.
By canonical variables analysis, VB and TSS were more sensitive to a reduced irrigation level when compared to the other variables. Possibly, buds under 70% ETc condition receive less carbohydrates from photosynthetically active leaves, negatively affecting the differentiation process of meristematic tissues in inflorescence primordia. Water status influences the formation of inflorescence primordia, either directly through the water volume available for the biosynthetic processes that occur during cell division and growth, or indirectly through its effect on photosynthetic activity, mineral nutrition, and hormonal balance7.
According to Monteiro et al.5, several endogenous and environmental factors influence the differentiation of meristematic tissue, where the vegetative bud instead of developing into inflorescence primordia, develops into tendril primordia, or sprout primordia. Thus, the irrigation rate application with 70% of ETc associated with an increase in leaf total soluble sugars between 45 and 50 DAPP is an indication of greater VB formation for the next cycle. The determination of TSS as an indicator of VB is advantageous because it is easier to measure, faster, labor-saving, and provides results regarding disturbances in the bud differentiation process.
The positive and strong correlation between FB and NPB was already expected, since bud fertility is a quantitative measure of the vine's potential to produce fruit or an indicator of the number of bunches that will be harvested in the next season1.
The decreases in A, E and gs with reduced irrigation are associated with stomatal closure and limitations in CO2 assimilation and transpiration. A similar result was found in 'Cabernet Sauvignon' grapevine grown in continental climate in USA2, on 'Chardonnay' grapevine grown in a temperate climate in Canada13 and in 'Syrah' grapevine grown in a semi-arid climate Brazil17.
Under drought condition, guard cells reduce stomatal opening, decreasing transpiration to avoid leaf dehydration, which limits CO2 diffusion into the leaf mesophyll and reduces photosynthesis7. Vines under water deficit tend to close their stomata to reduce canopy water loss, which prevents sharp declines in plant water potential and saves water for use during the growing season18. In this regard, the 'Arra 15' vine is sensitive to water deficit, as it induces the closure of stomata, causing reductions in transpiration and photosynthesis. Thus, the application of the irrigation level at 70% ETc during the period from 45 to 50 DAPP can reduce gas exchange, providing a decrease in the production of photoassimilates.
Each grapevine cultivar has different stomatal response to a given soil water potential19. Thus, the greater influence of the 100% ETc level on the variables A, E, gs, indicates that the soil water status provided by this level allows a higher capacity to keep the leaf active.
The strong and positive correlations between A, E and gs is also expected, as variations in gas exchange occur as a function of stomatal movement. Keller et al. (2016) found a strong and positive correlation between irrigation rates and A, E, and gs in Cabernet Sauvignon vines grown in a continental climate in the Columbia Valley, Washington, USA, corroborating our findings. Sequestering nutrients from photosynthesis for developing reproductive organs is common in some grapevine cultivars10. Thus, the correlation between FB and A indicates that the higher the production of photoassimilates from photosynthesis, the higher the formation of fertile buds.
Reductions in chlorophyll levels may also represent an acclimation response to water restriction, as the plant tends to conserve energy and hence capture less light energy, possibly avoiding photooxidative stress15. Thus, given the relationship between energy absorption and transfer, the levels of photosynthetic pigments may be related to net photosynthesis as well as to the growth and adaptation of 'Arra 15' to stress condition.
The decrease in photosynthetic pigment content due to water deficit (25% of field capacity) was diagnosed in Iranian native grape cultivars16. This authors reported that the loss of photosynthetic pigment under water deficit conditions occurs due to damage to the enzymes associated with photosynthesis, supporting the results obtained here.
The positive correlation between RCI and DB suggests that the increase in RCI may indicate disturbances in the bud differentiation process. It is likely that the intensity of this correlation is favored in conditions of excess soil water, which is supported by the canonical analysis, which showed an influence of the 130% ETc irrigation level on DB and RCI. Excessive irrigation increases the necrosis of grapevine buds, as a result of a higher vegetative vigor and shading in the renewal zone9.
The "b" chlorophyll content tends to increase in environments with a higher level of shading to enable the capture of energy in other wavelengths and transfer it to the "a" chlorophyll, which is responsible for photochemical reactions of photosynthesis20. It is possible that the increased RCI reflects an increase in "b" chlorophylls, and may indicate disturbances in the bud differentiation process. Thus, through RCI measurements taken between 45 and 50 DAPP, it is possible to adjust irrigation level to reduce the percentage of dead buds for the next cycle.
In light of the above, the water supply and its relationship with the physiology and morphology of reproductive organs may contribute to the decision-making by winegrowers. The findings reported in this research help to improve irrigation management from one cycle to another. Further studies should be focused on the processes of inflorescence primordium formation and how and when its metabolic pathways are affected, as it is a process greatly influenced by the water status of the vine.
The most appropriate irrigation strategy is the application of 115% ETc during 45 to 50 DAPP for the increase of fertile buds and potential bunches of the next cycle. The 115% ETc irrigation level does not impair the metabolism of 'Arra 15' grapevine grown in semi-arid condition. The highest percentage of fertile buds in 'Arra 15' is found in the middle region of the branch. TSS can be used as an indicator of water deficit during flowering and as an indicator of VB formation in the next cycle.
Material and methods
Experimental design and plant material
Field experiment was carried out in a commercial vineyard in the Senador Nilo Coelho Irrigated Project, located in the municipality of Petrolina, Pernambuco, Brazil (09° 22′ 26.75" S and 40° 38′ 4.29" O), during the years 2021 and 2022, period corresponding to three production cycles. Each production cycle began with production pruning. In the first cycle, production pruning was carried out on August 9, 2021; in the second cycle, production pruning was carried out on January 6, 2022; and in the third cycle, production pruning was carried out on August 15, 2022.
The vines assessed were nine-year-old 'Arra15', propagated by grafting (rootstock '1103 Paulson'), with a spacing between plants of 2.5 × 4.0 m, conducted in pergola system. The experimental research followed relevant institutional, national and international guidelines and legislation.
The American cultivar 'Arra 15' or 'ARRAFIFTEEN' was developed by the Grapa Company together with the Guimarra Vineyards Corporation in California, USA21. It is a white seedless grape that has high natural fertility of the buds, with two bunches per shoot on average, resistance to cracking of the berries due to rain during the ripening period, a low rate of degranulation, and a high yield with around 40 to 50 bunches per vine21.
The climate of the region according to the Köeppen classification is BSh type, which means semi-arid region, very warm and with irregular rainy season. The region has annual rainfall averages of approximately 500 mm, irregularly distributed and concentrated from November to April. It has an average annual relative humidity of 66% and an average annual air temperature of 26.5 °C, with the highest peaks from October onwards22. The average monthly values of maximum and minimum air temperature (°C) and precipitation (mm) over the three study cycles were obtained from the Automatic Agricultural Weather Station installed near the experiment site, and are shown in Fig. 9.

Seasonal variations of mean temperature (T mean), reference evapotranspiration (ET0), and precipitation of three consecutive crop cycles of 'Arra 15' grapevine. DAPP days after production pruning. Production pruning of cycle 1 carried out on 08/09/2021, cycle 2 on 01/06/2022 and cycle 3 on 08/15/2022.
Irrigation system consisted of two drip lines per planting row, installed at 2 m above the ground, with drippers spaced by 0.4 m. The irrigation shift was daily with a flow rate of 2.0 L h–1. Crop management, phytosanitary control, management of fertilization and vegetation were carried out by the farm according to the recommendations for viticulture in the São Francisco Valley1. Soil analysis were performed before implementation/beginning of the experiment at depths 0–30 cm and 30–60 cm (Table 2).
The experimental design used was randomized blocks with four replications, and the treatments consisted of five irrigation rates (70; 85; 100; 115 and 130% of crop evapotranspiration, —ETc) analyzed in three crop cycles. The experiment comprised a total of 20 plots, where each experimental plot consisted of 5 plants, with the three central plants being evaluated as the experimental units, totaling 60 useful plants.
The factor irrigation rates was quantified using the equation: ETc = ET0 × Kl × Kc. Reference Evapotranspiration (ET0) was calculated by the Penman–Monteith method23, using data collected by a weather station installed near the experimental area. The location effect (Kl) was determined according to Albuquerque24, using a value of Kl = 1 when the shaded area in the vineyard reached 75%. The value of 100% ETc corresponded to the Kc already determined for seedless grapevine cultivars in the Submédio São Francisco region25. The Kc was then adjusted for the other treatments.
The values of the Crop Coefficient (Kc) applied to the flowering and pea-size berries (30 to 50 days after pruning—DAPP) were: 0.25 (70% of ETc); 0.40 (85% of ETc); 0.55 (100% of ETc); 0.85 (115% of ETc); and 1.1 (130% of ETc).
Variables assessed
At 50 Days After Production Pruning (DAPP), between 9 a.m. and 10 a.m., gas exchange and relative chlorophyll index assessments, and leaf collection for biochemical analysis were performed in three useful plants of the plots. All assessments were performed on mature and fully expanded leaves exposed to the sun and located in the middle third of the plants. It is worth noting that 50 DAPP corresponds to the stage of undifferentiated primordium formation, which occurs during the growth of branches from the previous cycle, lasting 45 to 60 days after pruning26.
The parameters related to gas exchange were: net photosynthesis (A), stomatal conductance (gs), and transpiration (E), obtained through a portable infrared gas analyzer—IRGA (Model LI-6400XT, Li-Cor, Lincoln, NE, USA). The relative chlorophyll index (RCI) was obtained using a portable chlorophyll meter (Falker, model: ChlorofiLOG). The biochemical variables evaluated included the leaf contents of reducing sugars (RS), quantified by the dinitrosalicylic acid method27, and total soluble sugars (TSS), quantified by the antrona method28.
At 140 DAPP or 30 days after harvest, one branch per useful plant was collected from each plot, each containing ten buds, totaling 120 buds per treatment for bud analysis. 140 DAPP corresponded to the period when the inflorescence primordium is fully developed, taking the form of an axis with numerous protuberances similar in appearance to a bunch of grapes, and these protuberances correspond to the future flowers to be formed26. The bud analysis was carried out according to the methodology proposed by11, where the buds were dissected and then classified as fertile (FB), vegetative (VB), and dead (DB) according to the presence or absence of inflorescence primordia using a stereo microscope (binocular magnifier) with 30 × zoom (Fig. 10).

Visualization of 'Arra15' grapevine buds via cross-sections. (A) Fertile bud—FB, healthy bud with two inflorescence primordia, which are fragments of rounded shapes surrounding a central axis. (B) Vegetative bud—VB, healthy bud without inflorescence primordia. (C) Dead bud—DB, total loss of bud by necrosis. 30 × zoom.
For potential fertility, the position of the FB along the branch was identified in basal buds (1st to 3rd), middle buds (4th to 6th), and apical buds (7th to 10th), and the potential fertility of the basal branch region (PFBB), middle branch region (PFMB), and apical branch region (PFAB) were obtained. At the end of the evaluation period, the number of potential bunches per plant (NPB) was estimated at 50 branches per plant with ten buds per branch using the equation: \(\frac{\mathrm{number \,\,of\,\, branches }\times \mathrm{number \,\,of \,\,buds }\times \mathrm{accumulated\,\, fertility}}{100}\).
Statistical analyses
Data were submitted to analysis of variance by the F test. The effect of irrigation rate was considered fixed and the effect of cycle was considered random. In cases of significance, regression analysis was performed, testing linear and quadratic models for irrigation rates. In all cases, a 5% significance level was adopted.
Subsequently, canonical variables analysis was performed with the "candisc" package29 to verify the interrelationship between the treatments and variables evaluated. Finally, Pearson correlations were performed between the evaluated traits, where positive correlations were presented in green, while negative correlations were presented in red scale. The "qgraph" package30 was used to express the results graphically. All statistical analyses were performed using the R software31.
Data availability
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request32.
References
Vilar, P. F. I., Souza, E. I. D., Santos, L. D. S., Martinez, E. A. & Ribeiro, V. G. Phytoregulators on bud fertility and cluster quality of ‘Thompson seedless’ grapes grafted onto ‘Ramsey’rootstock. Rev. Caatinga 30, 97–108 (2017).
Keller, M. et al. Deficit irrigation alters grapevine growth, physiology, and fruit microclimate. Am. J. Enol. Vitic. 67, 426–435 (2016).
Baeza, P. et al. Effects of vine water status on yield components, vegetative response and must and wine composition. Adv. Grape Wine Biotechnol. 1, 73–94 (2019).
Li-Mallet, A., Rabot, A. & Geny, L. Factors controlling inflorescence primordia formation of grapevine: Their role in latent bud fruitfulness? A review. Botany 94, 147–163 (2016).
Monteiro, A. I., Malheiro, A. C. & Bacelar, E. A. Morphology, physiology and analysis techniques of grapevine bud fruitfulness: A review. Agriculture 11, 127 (2021).
Guilpart, N., Metay, A. & Gary, C. Grapevine bud fertility and number of berries per bunch are determined by water and nitrogen stress around flowering in the previous year. Eur. J. Agron. 54, 9–20 (2014).
Taiz, L., Zeiger, E., Moller, I. M. & Murphy, A. Fisiologia e Desenvolvimento Vegetal (Artmed, 2017).
Cabral, I. L., Carneiro, A., Nogueira, T. & Queiroz, J. Regulated deficit irrigation and its effects on yield and quality of Vitis vinifera L., touriga francesa in a hot climate area (Douro Region, Portugal). Agriculture 11, 774 (2021).
Williams, L. E., Grimes, D. W. & Phene, C. J. The effects of applied water at various fractions of measured evapotranspiration on reproductive growth and water productivity of Thompson seedless grapevines. Irrig. Sci. 28, 233–243 (2010).
Walker, R. P. et al. Sucrose metabolism and transport in grapevines, with emphasis on berries and leaves, and insights gained from a cross-species comparison. Int. J. Mol. Sci. 22, 7794 (2021).
Wohlfahrt, Y., Collins, C. & Stoll, M. Grapevine bud fertility under conditions of elevated carbon dioxide. Oeno One 53, 277–288 (2019).
El-Morsy, F. M., Gaser, A. S. & Mohamed, M. N. Morphological description and evaluation of six newly-introduced grape cultivars under Egyptian conditions. J. Plant Prod. 8, 1059–1070 (2017).
Balint, G. & Reynolds, A. G. Irrigation level and time of imposition impact vine physiology, yield components, fruit composition and wine quality of Ontario Chardonnay. Sci. Hortic. 214, 252–272 (2017).
Dayer, S., Prieto, J. A., Galat, E. & Peña, J. P. Leaf carbohydrate metabolism in M albec grapevines: Combined effects of regulated deficit irrigation and crop load. Aust. J. Grape Wine Res. 22, 115–123 (2016).
Oliveira, C. P. M. et al. Physiological and biochemical responses of apple trees to irrigation water depth in a semiarid region of Brazil. Ciênc. Agrotecnol. 44, e015620 (2020).
Fahim, S. et al. Multivariate discrimination of some grapevine cultivars under drought stress in Iran. Horticulturae 8, 871 (2022).
Bassoi, L. H., Chaves, A. R. M. & Teixeira, R. P. Responses of ’Syrah’ grapevine to deficit irrigation in the Brazilian semi-arid region. Agric. Water Manag. 258, 107–186 (2021).
Chaves, M. M. et al. Controlling stomatal aperture in semi-arid regions-the dilemma of saving water or being cool?. Plant Sci. 251, 54–64 (2016).
Tombesi, S., Frioni, T., Poni, S. & Palliotti, A. Effect of water stress “memory” on plant behavior during subsequent drought stress. Environ. Exp. Bot. 150, 106–114 (2018).
Martinazzo, E. G., Anese, S., Wandscheer, A. C. D. & Pastorini, H. Efeito do Sombreamento sobre o Crescimento Inicial e Teor de Clorofila Foliar de Eugenia uniflora Linn (Pitanga) – Família. Rev. Bras. Biociênc. 5, 162–164 (2007).
Karniel, ES, Giumarra, ES. Grape Plant Named ‟Arrafifteen”. US Pat. 2011/0219502 P1, 8 sets, 2011. 4p. Disponível em: https://patents.google.com/patent/US20110219502P1/en. Acesso em: 12 dez (2022).
Lopes, I. et al. Balanço hídrico em função de regimes pluviométricos na região de Petrolina-PE. Irriga 22, 443–457 (2017).
Allen, R. G., Pereira, L. S., Raes, D. & Smith, M. Crop evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56. Fao Rome 300, D05109 (1998).
Albuquerque PEP. Requerimento de água das culturas para fins de manejo e dimensionamento de sistemas de irrigação localizada (Embrapa Milho e Sorgo, Sete Lagoas, Circular Técnica) (2000).
Leão, PCS. Cultivo da videira. (Embrapa Semiárido, Petrolina, Sistemas de Produção, 1 (2004).
Botelho, R. V., Pires, E. J. P. & Terra, M. M. Fertilidade de gemas em videiras: Fisiologia e fatores envolvidos. Ambiência Guarapuava 2, 129–144 (2006).
Miller, G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31, 426–428 (1959).
Yemn, E. W. & Willis, A. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 57, 508 (1954).
Friendly, M., Fox, J., & Friendly, M. M. Package ‘candisc’. R package (2017).
Epskamp, S., Cramer, A. O., Waldorp, L. J., Schmittmann, V. D. & Borsboom, D. qgraph: Network visualizations of relationships in psychometric data. J. Stat. Softw. 48, 1–18 (2012).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria (2021).
Krishnan, S. et al. Bioethanol production from lignocellulosic biomass (water hyacinth): A biofuel alternative. In Bioreactors (eds Krishnan, S. et al.) 123–143 (Elsevier, 2020).
Acknowledgements
The authors would like to thank the Federal University of Mato Grosso do Sul (UFMS), Support Foundation for the Development of Education, Science and Technology of the State of Mato Grosso do Sul (FUNDECT), and National Council for Scientific and Technological Development (CNPq) – Grant number 334 303767/2020-0. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brazil (CAPES), Finance Code 001.
Author information
Authors and Affiliations
Contributions
Conceptualization, C.P.M.O. and P.E.T.; Data curation, C.P.M.O. and W.L.S.; Formal analysis, C.P.M.O. and P.E.T.; Funding acquisition, W.L.S., A.R.M.C., J.S.S.; Visualization, L.L.C. and R.G.; Writing – original draft, C.P.M.O. and P.E.T. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
de Oliveira, C.P.M., Simões, W.L., de Melo Chaves, A.R. et al. Water strategy improves the inflorescence primordia formation of 'Arra 15' grapevine in the Brazilian semiarid region. Sci Rep 14, 6678 (2024). https://doi.org/10.1038/s41598-024-57215-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-57215-7
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
