
Abstract
Hypoxic responses have been implicated in critical pathologies, including inflammation, immunity, and tumorigenesis. Recently, efforts to identify effective natural remedies and health supplements are increasing. Previous studies have reported that the cell lysates and the cell wall-bound lipoteichoic acids of Lactiplantibacillus plantarum K8 (K8) exert anti-inflammatory and immunomodulative effects. However, the effect of K8 on cellular hypoxic responses remains unknown. In this study, we found that K8 lysates had a potent suppressive effect on gene expression under hypoxia. K8 lysates markedly downregulated hypoxia-induced HIF1α accumulation in the human bone marrow and lung cancer cell lines, SH-SY5Y and H460. Consequently, the transcription of known HIF1α target genes, such as p21, GLUT1, and ALDOC, was notably suppressed in the K8 lysate supplement and purified lipoteichoic acids of K8, upon hypoxic induction. Intriguingly, K8 lysates decreased the expression of PHD2 and VHL proteins, which are responsible for HIF1α destabilization under normoxic conditions, suggesting that K8 may regulate HIF1α stability in a non-canonical pathway. Overall, our results suggest that K8 lysates desensitize the cells to hypoxic stresses and suppress HIF1α-mediated hypoxic gene activation.
Similar content being viewed by others


Introduction
Gene regulation is a fundamental and essential process through which living organisms develop, grow, maintain, and perish. Cells that constitute a living organism respond to the environmental and organismal signals and ensure proper gene expression based on the received signals. Hypoxia is an urgent condition that triggers rapid gene regulation in cells to manage low-oxygen stress1. DNA microarray and transcriptome profiling analyses have revealed that several hundred genes are differentially expressed upon exposure to hypoxic conditions2,3. These hypoxia-responsive genes are involved in diverse pathways including energy/glucose metabolisms, cell proliferation, and apoptosis4,5,6. Hypoxic stress is not only caused by an oxygen-deficient environment such as a high-altitude setting or anemia, but is also accompanied by certain physiological microenvironments, including tumors and inflammation7. For instance, hypoxic response is important for tumor progression and proliferation, and dozens of anticancer drugs that suppress hypoxic gene activation have been developed and preclinically administered8,9.
Hypoxia-inducible factor 1 (HIF1) consists of heterodimeric subunits with isoforms6,9. HIF1α is the major subunit and its expression is regulated by oxygen levels in the cell. Under normoxia, HIF1α in the cytoplasm is proline-hydroxylated and ubiquitinated for proteasomal degradation by prolyl hydroxylase (PHD) and von Hippel Lindau (VHL) proteins, respectively10,11,12. On the other hand, under hypoxic conditions, HIF1α is stabilized, translocated into the nucleus, and dimerized with constitutively expressed HIF1β12. The HIF heterodimer binds to the hypoxia response elements on its target genes to activate them. Therefore, the regulation of the HIF1α protein levels is critical to determine whether the cell suppresses or triggers hypoxic responses13. Notably, the over-expression of PHD proteins inhibits HIF1α accumulation, whereas HIF1α also reportedly induces PHD gene expression14,15,16, suggesting a possible feedback inhibition between the two enzymes. The VHL protein is known as a tumor suppressor and E3 ligase that ubiquitinates HIF1α for degradation17,18.
Lactiplantibacillus plantarum K8 (K8) is a Lactobacillus strain isolated from a spicy, fermented cabbage dish called kimchi19. Several research groups have identified interesting health benefits of K8. K8 lysates (K8-L) or K8 lipoteichoic acid (LTA; K8-A) associated with the cell wall can modulate immunity and suppress inflammation and obesity20,21,22,23. The lipoprotein and LTA components of K8 appear to interact with toll-like receptor 2 (TLR2) in host cells to suppress inflammatory responses24,25. K8-L and K8-A appear to regulate the gene expression of the critical genes in these pathways21,23,26. For example, K8-L reportedly inhibits the expression of PPARγ and C/EBPα, which are important for adipogenesis, and the pre-inflammatory factors, interleukin (IL)-1β and -6 and TNF-α at the transcriptional level21,25,27. K8-A supplement effectively suppresses Shigella flexneri peptidoglycans-stimulated cytokine expression by dampening the activation of MAPK pathway including ERK and JNK in human monocyte-like cell line, THP-120. It is not surprising that environmental substances, such as K8-L or K8-A, when treated on the cells or injected into the extracellular matrix, are capable of switching gene expression directly or indirectly through signal-transduction. Accumulating evidence has shown transcriptomic regulation by environmental and natural chemicals28,29,30,31. Understanding their physiological effects and gene regulatory mechanisms has been of interest because the applications of these natural chemicals, which are likely to have fewer adverse health effects, might replace those of synthetic chemicals.
Although K8-L and K8-A have been reported to modulate inflammation and immunity21,25,32,33, their effects on hypoxia, which is closely related to the inflammatory and immune responses, remain unknown. In addition, recent studies indicate that the chemicals released from gut bacteria can affect brain function and neurological health34 and lung function and infection35. We hypothesized that K8 cellular compounds might regulate cellular hypoxic responses through controlling HIF1α activation, with relatively low toxicity, and attempted to understand the effects of heat-treated K8-L (hK8-L) and K8-A on hypoxic gene expression in the human bone marrow and lung cancer cell lines, SH-SY5Y and H460, respectively. We hypothesized that HIF1α stabilization under hypoxic stress might be compromised by K8-L and if so, the expression of HIF1α target genes would be interfered to dampen hypoxic responses. We utilized cytotoxicity and molecular cell biology-based analyses to address these hypotheses. Our data suggest that hK8-L and K8-A are rarely cytotoxic and have potent suppressive activities against the stability of HIF1α and the activation of representative HIF1α-target genes under hypoxic stresses.
Materials and methods
K8 sample preparation
L. plantarum K8 (KCTC10887BP) was cultured in 1 L of MRS broth (BD Bioscience, USA) at 37 °C overnight and then cells were harvested by centrifugation at 8000 × g for 8 min. Bacterial cells were washed with deionized water (DIW) and re-suspended in DIW and disrupted by a microfluidizer five times at 27,000 psi. Disrupted L. plantarum K8 was freeze-dried to make L. plantarum K8 lysates. They were resuspended in PBS and the mixture was boiled at 100 °C for 20 min to inactivate live bacteria and proteins. Inactivation of live bacteria was confirmed by plating on MRS plates after heat treatment and no colonies were formed. PBS and PBS-soluble substances were removed by centrifugation. The PBS-insoluble pellet was dissolved in DMSO and designated hK8-L. The concentration of hK8-L was calculated as the weight of wet pellet divided by the volume of DMSO. Furthermore, hK8-L was centrifuged to remove the insoluble substances and debris, and the supernatant was filtered using 0.2 μm syringe filter to generate hK8-FL. Highly purified LTA was isolated from L. plantarum K8 (KCTC10887BP) by n-butanol extraction, as previously described36. Briefly, L. plantarum K8 was cultured in 8L MRS broth for 16 h at 37 °C. The cells were harvested by cengtrifugation at 8000×g for 8 min, suspended in 0.1 M sodium citrate buffer (pH 4.7), and disintegrated by ltrasonication. The disrupted bacterial cells were then mixed with an equal volume of n-butanol by stirring them for 30 min at RT. After centrifugation at 13,000×g for 20 min, the aqueous phase was evaporated, dialyzed against pyrogen free water, and equilibrated with 0.1 M sodium acetate buffer containing 15% n-propanol (pH 4.7). The LTA was purified by hydrophobic interaction chromatography on an octyl-Sepharose CL-4B (Sigma) column (2.5 cm by 20 cm). The column was eluted with a stepwise n-propanol gradient (100 mL of 20% n-propanol, 200 ml of 35% n-propanol, and 100 mL of 45% n-propanol). Then, the column fractions containing LTA were pooled after an inorganic phosphate assay, and the pool was dialyzed against water. The purity of the purified LTA was determined by measuring the protein and endotoxin contents through the conventional silver staining after polyacrylamide gel electrophoresis and being through the Limulus amebocyte lysate (LAL) assay (pLTA < 0.031 EU/ml) (BioWhittaker, U.S.A.). Nucleic acid contamination was assessed by measuring UV absorption at 260 and 280 nm.
List of L. plantarum K8-derived materials with abbreviations.
L. plantarum K8-derivatives | Description |
|---|---|
PBS-K8 | Heat-inactivated, PBS-soluble K8 |
hK8-L | Heat-inactivated, PBS-insoluble K8 pellet was dissolved in DMSO |
hK8-FL | hK8-L was centrifuged and the supernatant was filtered using 0.2 μm syringe filter |
K8-A | LTA purified from K8 lysates |
Cell culture and conditions
SH-SY5Y and H460 cells (American Type Culture Collection, USA) were grown in high glucose-DMEM (Cytiva, USA) and RPMI-1640 media (Gibco, USA), respectively, supplemented with 10% fetal bovine serum (FBS, Gibco, USA) and 1% penicillin/streptomycin (P/S, Thermo Fisher, USA) solution at 5% CO2 incubator at 37 °C. The cells were grown to 70–80% confluence in a 10 cm dish before splitting into a 6 or 96 well plate. K8 lysates (DMSO, control) were applied according to indicated concentrations for 24 h. CoCl2 (Cat. 60818, Sigma, USA) or deferoxamine methanesulfonate (DFO; D9533, Sigma, USA) salt, was treated to indicated final concentrations (H2O, control) in the halfway, 12 h (CoCl2) or 24 h (DFO) after K8 supplement, for 12 h before collecting the cells for the assays.
Cytotoxicity test
SH-SY5Y and H460 cells were grown to approximately 50–60% confluence in a 96-well plate in complete media. The media were exchanged with the fresh one including K8 lysates in DMSO, 1% of the total media volume. The K8 lysates-treated cells were incubated for 12 h before 100–200 μM CoCl2 in H2O or an equal amount of H2O, 1% of the total media volume. After an additional f12 h incubation, 10% of water-soluble tetrazolium salt (WST, DoGenBio, South Korea) was added to each well following the manufacturer’s instruction. The reaction was developed for 45 min to 1 h and the absorbance was measured at 450 nm using a microplate reader (Tecan, Switzerland).
Immunoblotting
SH-SY5Y cells were grown to approximately 70–80% confluence in 6-well plates. The media were exchanged with the fresh one including K8 lysates in DMSO, to 1% total media volume. The K8- or DMSO-treated cells were incubated for 12 h before CoCl2 supplement to the designated concentrations. After an additional 12 h incubation, the cells were washed with cold PBS and collected with RIPA buffer (Cell Signaling Technology, USA). The protein concentrations of cell lysates were measured using the Bradford assay (Bio-Rad, USA). From the measured protein concentrations, equal amounts of proteins were loaded and ran on a SDS–polyacrylamide gel. The separated proteins were transferred to the nitrocellulose membrane (GE Healthcare, USA). Primary antibodies were used for probing HIF1α (#14179, Cell Signaling Technology, USA), HIF2α (NB100-122, Novus biologicals, USA), p21 (sc-6246, SantaCruz Technology, USA), PHD2 (sc-271835, SantaCruz Technology, USA), β-ACTIN (MA5-15739, Invitrogen, USA), and α-tubulin (sc-8035, SantaCruz Technology, USA). Each antibody was diluted in a 5% skim milk (MBcell, South Korea or Bio-Rad, USA) solution or TBST solution [20 mM Tris-HCl, 137 mM NaCl, 0.1% (v/v) Tween-20, 5% (w/v) BSA, 0.025% (v/v) sodium azide and pH 7.6]. After the primary antibody incubation, the membranes were rinsed twice and washed four times, each for 10 min, with PBST solution [137 mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4, 1.5 mM KH2PO4, 0.1% (v/v) Tween-20 and pH 7.4]. The secondary antibodies, HRP-conjugated rabbit (#7074, Cell Signaling Technology) or mouse (#7076, Cell Signaling Technology), were diluted in 5% skim milk and incubated with the membranes. After incubation, the membranes were rinsed twice and washed with PBST solution four times, each for 10 min. Western Blotting Luminol Reagent (sc-2048, SantaCruz Technology, USA) or SuperSignal West Atto Ultimate Sensitivity Substrate (A38554, Thermo Fisher, USA) was used to develop signals. When indicated, immunoblotting images were quantified using Image J (NIH, USA).
Chromatin immunoprecipitation (ChIP)
ChIP was performed as described in our previous studies 37,38. Briefly, the cell lysis buffer including 5 mM PIPES (pH 8.0), 85 mM KCl, and 0.5% NP-40 was used to resuspend the crosslinked cells. The cells were resuspended with nuclei lysis buffer including 50 mM Tris-HCl (pH 8.0), 10 mM EDTA, and 1% SDS immediately before sonication. Sonication was performed on ice at 22% amplitude for 30 s with 2 min intervals between pulses (Vibra-Cell Processor VCX130, Sonics, USA). The number of pulses was optimized to produce DNA segments ranging between 100 and 1000 bp on a DNA gel. Protease inhibitors were freshly added to the cell and nuclei lysis buffers including 1 mM benzamidine (Sigma, USA), 0.25 mM PMSF (Sigma, USA), aprotinin (1:1000, A6279, Sigma, USA), 1 mM Na-metabisulfite (Sigma, USA), and 1 mM DTT. The ChIP-grade magnetic beads coated with protein G were purchased from Cell Signaling Technology (USA). The antibodies used in immunoprecipitation were Pol II (#39097, Thermo Fisher, USA) and HIF1α (#14179, Cell Signaling Technology, USA). After IP and reverse cross-linking, DNA was purified through Qiagen PCR purification kit (Qiagen, Germany). The ChIP products were analyzed by real-time PCR using SYBR Green Realtime PCR Master Mix (Applied Biosystems, USA) under thermal cycling as 1 min at 95 °C followed by 45 cycles of 15 s at 95 °C, 15 s at 55 °C, and 1 min at 72 °C. The results were analyzed as percent inputs. The primer sets amplifying the transcription start site and the gene body of the HIF1α target genes are listed in Table S1.
Quantitative real-time PCR (qRT-PCR)
cDNAs were constructed from 600 ng of the collected RNAs by reverse transcription using ReverTra Ace qPCR RT Master Mix (Toyobo, Japan). Equal amounts of resultant cDNAs were analyzed through Real-time qPCR using Applied Biosystems PowerUp SYBR Green Master Mix and according to the manufacturer’s instruction (Applied Biosystems, USA). StepOnePlus Real-Time PCR System was used (Thermo Fisher Scientific, USA). The thermal cycle used was 10 min for pre-denaturation, followed by 45 cycles of 95 °C for 15 s, 55 °C for 15 s, and 72 °C for 45 s. The primers used for the experiments were purchased from Integrated DNA technology (IDT, USA) and were summarized in Supplementary Table 1.
Fluorescence-activated cell sorting (FACS)
Flow cytometric analysis was performed to determine the cell cycle status. The cells were harvested by trypsinization and fixed with ice-cold 70% ethanol for 15 min at –20 °C. The cells were washed with PBS twice and suspended in the cold propidium iodide solution. Then the cells were incubated at room temperature for 15 min and analyzed by a flow cytometer (Attune NxT Flow Cytometry, Thermo Fisher).
Statistical analysis
Standard deviation was calculated and used to generate error bars. The Student’s t-test (unpaired, one-way) and Analysis of variance (ANOVA, two- or one-way) with Tukey post-hoc test were conducted to determine statistical significance. Graphs were generated using the Prism 8 software (GraphPad, Inc.).
Results
hK8-L suppresses HIF1α stabilization under hypoxia
K8 lysates and PBS-soluble and DMSO-soluble K8 (hK8-L) were generated as described in the method section. CoCl2 has been widely used to induce hypoxia-mimicking conditions in the in vitro cell-based experiments39,40. In previous studies, CoCl2 treatment has been validated to stabilize HIF1α39,41,42. A bone marrow cancer cell line, SH-SY5Y or a lung cancer cell line, H460, were treated with hK8-L at the indicated concentrations or DMSO (negative control) for 12 h before treatment with 100 μM CoCl2 or H2O (negative control) for an additional 12 h incubation (Fig. 1A). We investigated whether these chemicals affected cell viability and proliferation at the applied concentrations. Using the water-soluble tetrazolium salt (WST) assay, the viability of SH-SY5Y and H460 cells treated with up to 100 μg/mL hK8-L, with or without 100 μM CoCl2, was monitored. As shown in Fig. 1B and Supplementary Figure 1, hK8-L and CoCl2 rarely affected cell growth and proliferation, suggesting that hK8-L was innocuous and that CoCl2-induced hypoxic stress rarely caused cell death at the applied concentrations.

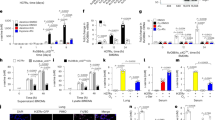
hK8-L inhibits HIF1α stabilization and its target gene expression under CoCl2-induced hypoxic stress. (A) A schematic representation of cell treatment with K8 lysates (0 – 100 μg/mL) and CoCl2 (0 – 200 μM) in this study. (B) Results of cell viability assay using the WST assay in SH-SY5Y cells. Error bars (n = 3), standard deviation (SD) throughout the figures. (C) Immunoblotting results showing the inhibitory effect of hK8-L on HIF1α (HIF1) stabilization/accumulation and p21 expression in CoCl2-mediated hypoxic stresses. hK8-L was supplemented to final concentrations of 0, 10, and 100 μg/mL. CoCl2 was treated to 0, 50, 100, and 200 μM of final concentrations. Tubulin was used as a loading control. (D) qRT-PCR results showing decreased mRNA expression of representative HIF1α-target genes, p21, GLUT1, ALDOC, and IL1β upon hypoxic stresses (0, 50, and 100 μM CoCl2) in the presence of 0 and 100 μg/mL hK8-L. n ≥ 3. Two-way ANOVA, Tukey multiple comparisons test. ns, not significant, **P < 0.01, ***P < 0.005, ****P < 0.0005.
Next, we investigated whether hK8-L affected hypoxic responses in SH-SY5Y cells. To answer this question, HIF1α protein levels at different doses of CoCl2 (0, 50, 100, and 200 μM) were quantified by immunoblotting, with 0, 10, and 100 μg/mL hK8-L (Fig. 1A, C). As expected, HIF1α protein was stabilized by CoCl2 treatment in a dose-dependent manner (Fig. 1C). Remarkably, hK8-L dramatically reduced HIF1α protein levels at 100 μg/mL and also mildly at 10 μg/mL, compared to the DMSO control. In contrast, neither the PBS-soluble K8 lysates nor 3-hydroxypropionic acid have HIF1α-suppressive effects (Supplementary Figure 2), suggesting that the DMSO-soluble fraction of K8 lysates is uniquely effective in alleviating HIF1α stabilization under hypoxic stresses.
hK8-L inhibits HIF1α-target gene activation under hypoxic stresses
HIF1α is a potent transcription activator that stimulates the transcription of a number of genes in the cell and is crucial for cellular metabolic homeostasis and fate decisions. One of the HIF1α-target genes is p21, a cell cycle regulator43. Transcriptional activation at p21 gene is well-known to be mediated by p53, an upstream transcription factor that senses genomic instability44,45. On the other hand, p21 can also be activated in a p53-independent manner, and HIF1α has been reported to directly turn on the transcription of p21 by displacing MYC43. Therefore, the expression of p21 with or without hK8-L and CoCl2 was investigated. Immunoblotting results indicated that CoCl2 supplementation increased p21 expression, which was positively correlated with the HIF1α levels (Fig. 1C). Importantly, hK8-L not only reduced HIF1α protein levels but also p21 levels in 200 μM CoCl2 in a dose-dependent manner (Fig. 1C). These results suggested that HIF1α accumulation and its transactivation at p21 gene under hypoxic stresses are strongly inhibited by hK8-L.
Because HIF1α is a transcription factor, the transcription status of its known target genes was monitored. We selected GLUT1 and ALDOC, important genes involved in glucose metabolism; IL1β, a critical cytokine for inflammation and immunity; and p21, a potent cell cycle modulator, as these pathways are mainly upregulated by HIF1α activation46,47,48,49. When SH-SY5Y cells were treated with 0, 50, and 100 μM CoCl2 in SH-SY5Y cells, the mRNA expression of p21, GLUT1, ALDOC, and IL1β notably increased in a dose-dependent manner (Fig. 1D, white bars). In contrast, hK8-L supplementation at 100 μg/mL significantly interfered with the increase in all tested genes, except for GLUT1 at the highest CoCl2 concentration (Fig. 1D, colored bars). Overall, these results suggest that hK8-L suppresses the gene expression of HIF1α target genes under hypoxic stresses.
In the consideration that K8-L might directly interfere with CoCl2, rather than interfering with the HIF1α stabilization, we also tested the effect of hK8-L on hypoxic responses using a different hypoxia-inducing chemical, deferoxamine methanesulfonate (DFO). hK8-L was supplemented to SH-SY5Y cells at final concentrations of 0 and 100 μg/mL and hypoxic stress was induced to the cells by the addition of DFO to final concentrations of 0, 60, and 120 μM. The protein levels of HIF1α, HIF2α, and p21 were quantified through immunoblotting (Fig. 2A). The results were consistent with the CoCl2 data (Fig. 1C), showing that HIF1α was stabilized at 60 and 120 μM DFO, while hK8-L markedly suppressed the stabilization. Although HIF2α was dramatically reduced by hK8-L in 60 μM DFO, HIF2α appeared to be somewhat stabilized in 120 μM DFO, showing only a slight reduction in comparison to the DMSO control (Fig. 2A). The expression of p21 level was increased under DFO treatment, which was notably decreased in the presence of hK8-L (Fig. 2A). In addition, we measured the mRNA expression levels of HIF1α target genes, GLUT1, ALDOC, p21, and IL1β in the same DFO conditions, with or without hK8-L (Fig. 2B). The qRT-PCR data indicated the gene activation of these HIF1α target genes under DFO-induced hypoxic stresses and an overall suppression of the activation in the cells that were pre-supplemented with hK8-L, compared to the DMSO controls. This compromised HIF1α target gene expression in the presence of hK8-L in both CoCl2- and DFO-induced hypoxic conditions suggest that hK8-L alleviates HIF1α stabilization under cellular hypoxia, rather than interfering with CoCl2 or DFO.

hK8-L inhibits HIF1α stabilization and its target gene expression under DFO-induced hypoxic stress. (A) Immunoblotting results showing the inhibitory effect of hK8-L on HIF1α (HIF1) and HIF2α (HIF2) stabilization/accumulation and p21 expression in SH-SY5Y cells upon DFO-induced hypoxic stress. hK8-L was supplemented to final concentrations of 0, 10, and 100 μg/mL for 12 h before DFO addition. DFO was supplemented to 0, 60, and 120 μM of final concentrations for 24 h before collecting the cells. (B) qRT-PCR results showing decreased mRNA expression of representative HIF1α-target genes, p21, GLUT1, ALDOC, and IL1β upon hypoxic stresses (0, 60, and 120 μM DFO) in the presence of 0 and 100 μg/mL hK8-L. n ≥ 3. Unpaired Student’s one-tailed t-test was applied. ns, not significant, *P < 0.05, **P < 0.01, ***P < 0.005, ****P < 0.0005.
hK8-L interferes with HIF1α and Pol II recruitment on HIF1α-target genes under hypoxia
To verify whether HIF1α destabilization is the major cause of the reduced gene expression of p21, GLUT1, ALDOC, and IL1β in the presence of hK8-L, the enrichment level of HIF1α in the transcription start sites of these genes was quantified using ChIP-qPCR. The data showed that hK8-L treatment alone either barely changed or even slightly increased HIF1α enrichment under the normoxic conditions (–CoCl2; Fig. 3A), whereas hK8-L under the hypoxic conditions (+ CoCl2) significantly and consistently prevented HIF1α from increasing in all four genes (Fig. 3A). A recent study reported that hypoxia-inducible genes harbor promoter-proximally paused Pol II50. Therefore, Pol II occupancy was monitored in the transcription start site (TSS < ± 300 nt from + 1) and the gene body (GB > + 301 nt from + 1) of these HIF1α target genes using the target site-specific primer sets (Table S1) by ChIP-qPCR. Consistently with the changes in HIF1α occupancy, the results showed that the increase in Pol II occupancy in both TSS and GB under hypoxia was attenuated in the presence of hK8-L and CoCl2, compared to that in controls, DMSO and CoCl2 supplementation (Fig. 3B, C). Notably, Pol II occupancy levels were positively correlated with the levels of HIF1α enrichment levels and transcriptional availability, confirming HIF1α function as a transcriptional activator for these genes (Fig. 3B, C). These results also demonstrate the dependence of HIF1α recruitment on the transcriptional activation of p21, GLUT1, ALDOC, and IL1β, and the potent ability of hK8-L to reduce HIF1α and the transcription of these genes under the hypoxic conditions.

hK8-L interferes with HIF1α and Pol II recruitment under hypoxic stress. (A) ChIP-qPCR results showing HIF1α occupancy changes in the transcription start sites (TSS) of p21, GLUT1, ALDOC, and IL1β gene. SH-SY5Y cells were treated with 0 or 100 μg/mL hK8-L for 12 h before supplementing 0 or 200 μM CoCl2 for additional 12 h (Fig. 1A). The cells were cross-linked and sonicated for immunoprecipitation using HIF1α antibody. n = 3. Unpaired Student’s one-tailed t-test was applied. ns, non-significant throughout the figures, *P < 0.05, **P < 0.01, ***P < 0.005, ****P < 0.0005. (B) ChIP-qPCR results showing the Pol II occupancies in the transcription start sites (TSS) of p21, GLUT1, ALDOC, and IL1β gene. n = 3. Unpaired Student’s one-tailed t-test was applied. *P < 0.05, **P < 0.01, ***P < 0.005, ****P < 0.0005. (C) ChIP-qPCR results showing the Pol II occupancies in the gene bodies (GB) of p21, GLUT1, ALDOC, and IL1β gene. n ≥ 3. Unpaired Student’s one-tailed t-test was applied. *P < 0.05, **P < 0.01, ***P < 0.005.
hK8-FL possesses a HIF1α-inhibitory effect similar to hK8-L
As shown Fig. 1B, hK8-L, the heat-inactivated K8 lysates in DMSO, excluding the PBS-soluble substances, was nontoxic to the tested cell lines. However, it is not a homogenized mixture, which includes cell debris and particles in irregular masses that are visible under a light microscope. To remove these masses and obtain a finer, completely DMSO-soluble fraction, hK8-L was centrifugated and the resulting supernatant was filtered by passing through a 0.2-μm filter. The filtered hK8-L (hK8-FL, hereafter) was tested for cytotoxicity using the WST assay. SH-SY5Y cells were treated with hK8-FL at 0, 10, 50, 75, 100, and 500 μg/mL for 12 h before supplementation with 100 μM CoCl2 for an additional 12 h (Fig. 1A). The results showed that hK8-FL was innocuous and did not affect cell viability at the tested concentrations regardless of the presence of CoCl2 (Fig. 4A). Similar results were obtained for H460 cells (Fig. 4B). We questioned whether hK8-FL could suppress the increase in HIF1α under hypoxic condition and if so, the inhibitory effects for the two were comparable. The level of HIF1α increase, with 200 μM CoCl2 treatment, was quantified by immunoblotting, comparing the cells pre-treated with DMSO, hK8-L and hK8-FL (Fig. 4B). The immunoblotting results indicated that both hK8-L and hK8-FL significantly reduced HIF1α protein levels in CoCl2-treated H460 cells and the levels of reduction with the two compounds were comparable (Fig. 4C). Consistently with the HIF1α results, p21 protein levels also decreased in hK8-L- and hK8-FL-treated SH-SY5Y cells, both to similar levels (Fig. 4D).

hK8-L and hK8-FL have a comparable inhibitory effect on HIF1α stabilization. (A) Results of cell viability assay using WST in SH-SY5Y cells. hK8-FL (0, 10, 50, 75, 100, and 500 μg/mL) was pre-incubated with the cells for 12 h before subjecting to the hypoxic stress induced by 100 μM CoCl2 (or equivalent volume of H2O as the control) for 12 h. n = 4. (B) Results of cell viability assay using WST in H460 cells. Same K8 and CoCl2 conditions were applied as (A). n = 3. (C) Immunoblotting results showing the comparable inhibitory effects of hK8-L (100 μg/mL) and hK8-FL (100 μg/mL), in comparison to the DMSO control, on HIF1α accumulation during CoCl2 (200 μM)-mediated hypoxic stresses in H460 cells. One-way ANOVA, Tukey multiple comparisons test. n ≥ 3. *P < 0.05, **P < 0.01. (D) Immunoblotting results showing the comparable inhibitory effects of hK8-L (100 μg/mL) and hK8-FL (100 μg/mL) on p21 gene activation during CoCl2 (200 μM)-mediated hypoxic stresses in SH-SY5Y cells. n ≥ 3. One-way ANOVA, Tukey multiple comparisons test. *P < 0.05, **P < 0.01.
Purified K8 LTA suppresses HIF1α-target gene expression
Next, K8-A, LTA purified from K8 lysates, was tested to determine whether it could suppress the hypoxic responses. Similar to hK8-L and hK8-FL, K8-A at 10, 50, and 100 μg/mL did not display cytotoxicity in SH-SY5Y cells (Fig. 5A). The protein level of HIF1α was measured using immunoblotting. The results indicated that unlike hK8-L and hK8-FL, the level of HIF1α after a treatment of 200 μM CoCl2 appeared less affected by K8-A treatment (Fig. 5B). However, we noticed that the HIF1α species with higher molecular weights (marked with a bracket in Fig. 5B) showed a decreasing trend in K8-A-treated cells, which may suggest certain changes in the post-translational modifications or the dimerization status of HIF1α by K8-A. The mRNA expression of p21, ALDOC, and GLUT1 significantly decreased under hypoxic stresses in K8-A-treated cells, as well as under normoxic conditions, except for p21 at 50 μg/mL K8-A (Fig. 5C–E).

K8-A inhibits HIF1α-target gene activation. (A) Results of cell viability assay using WST in SH-SY5Y cells. K8-A was supplemented to final concentrations 0, 10, 50, and 100 μg/mL and the cells were incubated for 24 h. n = 3. (B) Immunoblotting results showing the effect of K8-A on HIF1α protein in SH-SY5Y cells. K8-A was treated to final concentrations of 0, 50, and 100 μg/mL. CoCl2 was treated to a final concentration of 200 μM. K8-A was pre-incubated for 12 h before applying CoCl2-mediated hypoxic stress for 12 h. β-ACTIN was used as a loading control. NE, HeLa NE as a technical reference. HIF1α species with higher molecular weights marked with a bracket. (C) qRT-PCR results showing the effect of K8-A on the mRNA expression of p21, a HIF1α-target gene during normoxic and CoCl2 (200 μM)-mediated hypoxic conditions in SH-SY5Y cells. n ≥ 3. Unpaired Student’s one-tailed t-test was applied. **P < 0.01, ***P < 0.005, **** < 0.0005. (D) qRT-PCR results showing the effect of K8-A on the mRNA expression of ALDOC, a HIF1α-target gene during normoxic and CoCl2 (200 μM)-mediated hypoxic conditions in SH-SY5Y cells. n ≥ 3. Unpaired Student’s one-tailed t-test was applied. *P < 0.05, ***P < 0.005. (E) qRT-PCR results showing the effect of K8-A on the mRNA expression of GLUT1, a HIF1α-target gene during normoxic and CoCl2 (200 μM)-mediated hypoxic conditions in SH-SY5Y cells. n ≥ 3. Unpaired Student’s one-tailed t-test was applied. ***P < 0.005.
hK8-L inhibits the gene expression of HIF1α regulators, PHD2 and VHL
Lastly, we analyzed the mRNA expression levels of prolyl hydroxylase domain protein 2 (PHD2, also known as EGLN1) and VHL, two proteins that regulate HIF1α stability and hypoxic responses, using qRT-PCR. In the control (DMSO) group, mRNA levels of PHD2 and VHL genes were increased and decreased upon 200 μM CoCl2 treatment in SH-SY5Y cells, respectively (Fig. 6A). The cells were supplemented with 100 μg/mL hK8-L or hK8-FL prior to CoCl2 treatment. The expression of PHD2 and VHL mRNAs decreased, compared to that in the control (Fig. 6A), suggesting that hK8 down-regulates HIF1α probably through a different pathway from the classical PHD2/VHL-mediated degradation pathway. It is noted that PHD2 destabilizes HIF1α and also the transcription of PHD2 is reportedly dependent on HIF1α16. The PHD2 protein was quantified using immunoblotting. Consistently with the qRT-PCR results, hK8-FL treatment decreased the PHD2 protein levels (Fig. 6B), probably due to HIF1α destabilization. Overall, these data suggested that DMSO-soluble K8 lysates have the effect to inhibit HIF1α stability, while decreasing the expression of PHD2 and VHL genes in SH-SY5Y cells.

hK8-L inhibits HIF1α-regulators, PHD2 and VHL. (A) qRT-PCR results showing the effect of hK8-L (100 μg/mL) and hK8-FL (100 μg/mL) on the mRNA expression of PHD2, a HIF1α-regulator and -target gene and VHL during normoxic (- CoCl2) and CoCl2 (200 μM, + CoCl2)-mediated hypoxic conditions in SH-SY5Y cells. n ≥ 3. Unpaired Student’s one-tailed t-test was applied. **P < 0.01, ***P < 0.005, ****P < 0.0005. (B) Immunoblotting results showing the effect of hK8-L on PHD2 protein levels in SH-SY5Y cells. hK8-L was treated to final concentrations of 0 (DMSO only) and 100 μg/mL. CoCl2 was treated to final concentrations of 0, 50, 100, and 200 μM. α-Tubulin was used as a loading control.
Discussion
It is important to note that hypoxic responses occur in inflammation and cancer growth. Chronic inflammation often develops into cancers. For example, people with chronic inflammatory bowel diseases such as Crohn disease are exposed to a higher risk to colon cancers51,52. It is well-known that HIF1α is stabilized also in normoxia during inflammation53. While chronic inflammation shows the increased level of HIF1α, a study reported that HIF1α is required for the resolution of bowel inflammation54. Another study reported that HIF1α aggravates psoriasis by inhibiting BMP6 expression55. As the physiological roles of HIF1α for hypoxic responses, inflammation, and immunity are important, molecules to control the HIF1α level have been developing and clinically tried56. The identification of the novel function of lactobacillus K8 extracts to modulate HIF1α stability in this study suggests a way to develop a potential natural remedy to treat HIF1α deregulation, which is implicated in different disease pathologies.
In this study, we tested Lactiplantibacillus plantarum K8 lysates on the human hypoxic responses in vitro. This property of K8 lysates is pertained to the DMSO-soluble fractions of the freeze-powdered K8 lysates (hK8-L and hK8-FL). Both hK8-L and hK8-FL significantly inhibit HIF1α stabilization, thereby interfering with the HIF1α-mediated hypoxic gene expression (Figs. 1, 2, 3, 4). Representative HIF1α-target genes, including p21, ALDOC, GLUT1, and IL1β, were shown to be downregulated under hypoxic stresses in the presence of hK8-L (Figs. 1, 2, 3). The word “modulation” seems appropriate to describe the K8 results because hK8-L slightly increases the mRNA synthesis and HIF1α/Pol II occupancy of some genes in normoxia, while it inhibits the increase in hypoxia (Figs. 1D, 3A–C). These data suggest that hK8-L may mildly stimulate hypoxic responses including glucose uptake (GLUT1) and metabolism (ALDOC) under normoxic conditions, while it functions as a break to prevent these genes from being acutely transcribed in hypoxia.
In addition, hK8-FL and K8-A exhibited an inhibitory effect on hypoxia-induced gene expression (Figs. 4, 5). Filter cleared fraction of hK8-L (hK8-FL) is void of DMSO-insoluble substances larger than 0.2 μm. Under microscopic examination, we noted that centrifugally cleared hK8-L included a number of irregular sized, visible substances, thus not homogenized. The experimental results with homogenzed hK8-FL, showing the suppressive effect of hK8-FL on HIF1α, is meaningful in that it confirms the effects to be contributable to the DMSO-soluble, small compounds in K8 lysates. In addition, we qualitatively and quantitatively analyzed hK8-L and PBS-K8 using gas chromatography and mass spectrometry (GC/MS) (Table S2–4; Supplementary Data 1–3). Overall, in the analysis of volatile components, K8 powder (52 compounds), hK8-L (24 compounds), and PBS-K8 (33 compounds) don’t share compounds with one another. Between hK8-L and PBS-K8, only two compounds were commonly identified. Because there are no chemical libraries for K8 to validate and compare with our GC/MS data, it is difficult to pinpoint certain chemicals that might contribute to the suppressive effect on HIF1α stabilization among the compounds. However, our GC/MS data verified chemically-differentiated K8 fractions by two solvents, PBS and DMSO, and our experiments showed DMSO to efficiently extract the effect molecule(s) from K8 freeze-dried powder.
Initially, we hypothesized that VHL may be increased by hK8-L pretreatment before hypoxic stress. This hypothesis was based on the fact that VHL is an E3 ligase for HIF1α for degradation5,13,18, which could explain how HIF1α is decreased in the presence of hK8-L in spite of hypoxic stresses. Instead, qRT-PCR results showed a decrease in VHL expression in the presence of hK8-L and hypoxic stress (Fig. 5A). These data suggest that the K8-mediated HIF1α protein reduction under hypoxia is probably not through the classical PHD–VHL degradation pathway but through other mechanisms. The decrease of VHL may explain a mild increase of HIF1α-target gene expression under normoxia (Figs. 1D, 2B, 3A–C). In addition, it is noted that VHL has HIF-dependent and -independent functions such as the regulation of apoptosis and senescence and the inhibition of transcription57,58, suggesting that K8 could affect these biological pathways through VHL down-regulation. Further studies are required to identify the exact underlying mechanisms: how K8 lysates inhibit HIF1α protein accumulation under hypoxic stresses and how K8 down-regulates VHL transcription. Besides, transcriptome analysis is necessary for identifying all hypoxic genes regulated by hK8-L/FL and for understanding the impact of these compounds on hypoxic and overall gene regulation.
Although our current study solely focused on the effect of hK8 on HIF1α stability and its target genes, hK8-L appeared to inhibit HIF2α stabilization under hypoxia as well (Fig. 2A). HIF1α and HIF2α, while both are important for cellular hypoxic responses, do not have redundant functions and the same target genes57,59,60. Studies have reported that HIF1α regulates metabolic pathways including glycolysis, while HIF2α regulates more diverse pathways such as tumor immunity and ribosomal biosynthesis59,60. To obtain a complete picture of the hypoxia-inducible gene regulation of hK8 compounds, further studies are required to understand hK8 effects on HIF2α and the expression of its target genes.
Together, our data support our hypothesis that K8 cellular compounds, hK8-L/FL and K8-A regulate hypoxic responses though controlling HIF1α in SH-SY5Y and H460 cells. Although mechanistic understanding of how hK8 destabilizes HIF1α is currently limited, this study suggests that hK8-L/FL and K8-A inhibit the accumulation of HIF1α and repress the expression of representative HIF1α target genes under hypoxic stresses. Our data provide the first pieces of evidence that natural compounds in lactobacillus K8 lysates have the potential to be developed into a hypoxic regulator. Previous studies have reported quite a few specific natural compounds, mostly from herbs, to inhibit HIF18,61. Our hK8 is unique in that it is undefined, heat-inactivated lysates of bacteria, originated from popular fermented cabbage dish, Kimchi. In addition, it is notable that our extraction methods showed solvent-specific, differential effects of hK8 lysates derivatives on HIF1α stabilization. We propose that hK8-L/FL may provide health benefits to humans as a natural modulator/suppressor of hypoxic responses, which are critical for cancer cell proliferation and inflammation.
Data availability
All data are available in the manuscript or as supplementary information.
References
Lee, P., Chandel, N. S. & Simon, M. C. Cellular adaptation to hypoxia through hypoxia inducible factors and beyond. Nat. Rev. Mol. Cell Biol. 21, 268–283. https://doi.org/10.1038/s41580-020-0227-y (2020).
Mazzatti, D., Lim, F. L., O’Hara, A., Wood, I. S. & Trayhurn, P. A microarray analysis of the hypoxia-induced modulation of gene expression in human adipocytes. Arch. Physiol. Biochem. 118, 112–120. https://doi.org/10.3109/13813455.2012.654611 (2012).
Nakayama, K. et al. Large-scale mapping of positional changes of hypoxia-responsive genes upon activation. Mol. Biol. Cell 33, ar72. https://doi.org/10.1091/mbc.E21-11-0593 (2022).
Kierans, S. J. & Taylor, C. T. Regulation of glycolysis by the hypoxia-inducible factor (HIF): implications for cellular physiology. J. Physiol. 599, 23–37. https://doi.org/10.1113/JP280572 (2021).
Hubbi, M. E. & Semenza, G. L. Regulation of cell proliferation by hypoxia-inducible factors. Am. J. Physiol. Cell Physiol. 309, C775-782. https://doi.org/10.1152/ajpcell.00279.2015 (2015).
Greijer, A. E. & van der Wall, E. The role of hypoxia inducible factor 1 (HIF-1) in hypoxia induced apoptosis. J. Clin. Pathol. 57, 1009–1014. https://doi.org/10.1136/jcp.2003.015032 (2004).
Triner, D. & Shah, Y. M. Hypoxia-inducible factors: a central link between inflammation and cancer. J. Clin. Investig. 126, 3689–3698. https://doi.org/10.1172/JCI84430 (2016).
Ma, Z. et al. Targeting hypoxia-inducible factor-1, for cancer treatment: Recent advances in developing small-molecule inhibitors from natural compounds. Semin. Cancer Biol. 80, 379–390. https://doi.org/10.1016/j.semcancer.2020.09.011 (2022).
Wicks, E. E. & Semenza, G. L. Hypoxia-inducible factors: cancer progression and clinical translation. J. Clin. Investig. https://doi.org/10.1172/JCI159839 (2022).
Tanimoto, K., Makino, Y., Pereira, T. & Poellinger, L. Mechanism of regulation of the hypoxia-inducible factor-1 alpha by the von Hippel-Lindau tumor suppressor protein. EMBO J. 19, 4298–4309. https://doi.org/10.1093/emboj/19.16.4298 (2000).
Appelhoff, R. J. et al. Differential function of the prolyl hydroxylases PHD1, PHD2, and PHD3 in the regulation of hypoxia-inducible factor. J. Biol. Chem. 279, 38458–38465. https://doi.org/10.1074/jbc.M406026200 (2004).
Koyasu, S., Kobayashi, M., Goto, Y., Hiraoka, M. & Harada, H. Regulatory mechanisms of hypoxia-inducible factor 1 activity: Two decades of knowledge. Cancer Sci. 109, 560–571. https://doi.org/10.1111/cas.13483 (2018).
Yu, F., White, S. B., Zhao, Q. & Lee, F. S. HIF-1alpha binding to VHL is regulated by stimulus-sensitive proline hydroxylation. Proc. Natl. Acad. Sci. USA 98, 9630–9635. https://doi.org/10.1073/pnas.181341498 (2001).
Erez, N. et al. Expression of prolyl-hydroxylase-1 (PHD1/EGLN2) suppresses hypoxia inducible factor-1alpha activation and inhibits tumor growth. Cancer Res. 63, 8777–8783 (2003).
Marxsen, J. H. et al. Hypoxia-inducible factor-1 (HIF-1) promotes its degradation by induction of HIF-alpha-prolyl-4-hydroxylases. Biochem. J. 381, 761–767. https://doi.org/10.1042/BJ20040620 (2004).
Fujita, N. et al. Expression of prolyl hydroxylases (PHDs) is selectively controlled by HIF-1 and HIF-2 proteins in nucleus pulposus cells of the intervertebral disc: distinct roles of PHD2 and PHD3 proteins in controlling HIF-1alpha activity in hypoxia. J. Biol. Chem. 287, 16975–16986. https://doi.org/10.1074/jbc.M111.334466 (2012).
Chitrakar, A., Budda, S. A., Henderson, J. G., Axtell, R. C. & Zenewicz, L. A. E3 ubiquitin ligase von Hippel-Lindau protein promotes Th17 differentiation. J. Immunol. 205, 1009–1023. https://doi.org/10.4049/jimmunol.2000243 (2020).
Kamura, T. et al. Activation of HIF1alpha ubiquitination by a reconstituted von Hippel-Lindau (VHL) tumor suppressor complex. Proc. Natl. Acad. Sci. USA 97, 10430–10435. https://doi.org/10.1073/pnas.190332597 (2000).
Kim, H. et al. Effects of oral intake of kimchi-derived Lactobacillus plantarum K8 lysates on skin moisturizing. J. Microbiol. Biotechnol. 25, 74–80. https://doi.org/10.4014/jmb.1407.07078 (2015).
Kim, H. G. et al. Lactobacillus plantarum lipoteichoic acid down-regulated Shigella flexneri peptidoglycan-induced inflammation. Mol. Immunol. 48, 382–391. https://doi.org/10.1016/j.molimm.2010.07.011 (2011).
Kim, H. et al. Lactobacillus plantarum lipoteichoic acid alleviates TNF-alpha-induced inflammation in the HT-29 intestinal epithelial cell line. Mol. Cells 33, 479–486. https://doi.org/10.1007/s10059-012-2266-5 (2012).
Ahn, K. B., Baik, J. E., Park, O. J., Yun, C. H. & Han, S. H. Lactobacillus plantarum lipoteichoic acid inhibits biofilm formation of Streptococcus mutans. PLoS ONE 13, e0192694. https://doi.org/10.1371/journal.pone.0192694 (2018).
Ma, Y., Fei, Y., Han, X., Liu, G. & Fang, J. Lactobacillus plantarum alleviates obesity by altering the composition of the gut microbiota in high-fat diet-fed mice. Front. Nutr. 9, 947367. https://doi.org/10.3389/fnut.2022.947367 (2022).
Lee, I. C. et al. Lipoproteins contribute to the anti-inflammatory capacity of Lactobacillus plantarum WCFS1. Front. Microbiol. 11, 1822. https://doi.org/10.3389/fmicb.2020.01822 (2020).
Le, B. & Yang, S. H. Efficacy of Lactobacillus plantarum in prevention of inflammatory bowel disease. Toxicol. Rep. 5, 314–317. https://doi.org/10.1016/j.toxrep.2018.02.007 (2018).
Kim, J. Y. et al. Lipoteichoic acid isolated from Lactobacillus plantarum suppresses LPS-mediated atherosclerotic plaque inflammation. Mol. Cells 35, 115–124. https://doi.org/10.1007/s10059-013-2190-3 (2013).
Kim, H., Lim, J.-J., Shin, H. Y., Suh, H. J. & Choi, H.-S. Lactobacillus plantarum K8-based paraprobiotics suppress lipid accumulation during adipogenesis by the regulation of JAK/STAT and AMPK signaling pathways. J. Funct. Foods 87, 104824. https://doi.org/10.1016/j.jff.2021.104824 (2021).
Kim, D. et al. Arsenic hexoxide has differential effects on cell proliferation and genome-wide gene expression in human primary mammary epithelial and MCF7 cells. Sci. Rep. 11, 3761. https://doi.org/10.1038/s41598-021-82551-3 (2021).
Jeong, J. et al. Tetraarsenic oxide affects non-coding RNA transcriptome through deregulating polycomb complexes in MCF7 cells. Adv. Biol. Regul. 80, 100809. https://doi.org/10.1016/j.jbior.2021.100809 (2021).
Bunch, H. et al. Evaluating cytotoxicity of methyl benzoate in vitro. Heliyon 6, e03351. https://doi.org/10.1016/j.heliyon.2020.e03351 (2020).
Jeon, S., Lee, S.-H., Roh, J., Kim, J.-E. & Bunch, H. Glyphosate influences cell proliferation in vitro. All Life 13, 54–65. https://doi.org/10.1080/26895293.2019.1702108 (2020).
Kim, H., Jung, B. J., Jeong, J., Chun, H. & Chung, D. K. Lipoteichoic acid from Lactobacillus plantarum inhibits the expression of platelet-activating factor receptor induced by Staphylococcus aureus lipoteichoic acid or Escherichia coli lipopolysaccharide in human monocyte-like cells. J. Microbiol. Biotechnol. 24, 1051–1058. https://doi.org/10.4014/jmb.1403.03012 (2014).
Kim, H. G. et al. Lipoteichoic acid from Lactobacillus plantarum elicits both the production of interleukin-23p19 and suppression of pathogen-mediated interleukin-10 in THP-1 cells. FEMS Immunol. Med. Microbiol. 49, 205–214. https://doi.org/10.1111/j.1574-695X.2006.00175.x (2007).
Sittipo, P., Choi, J., Lee, S. & Lee, Y. K. The function of gut microbiota in immune-related neurological disorders: A review. J. Neuroinflamm. 19, 154. https://doi.org/10.1186/s12974-022-02510-1 (2022).
Chiu, Y. C., Lee, S. W., Liu, C. W., Lan, T. Y. & Wu, L. S. Relationship between gut microbiota and lung function decline in patients with chronic obstructive pulmonary disease: A 1-year follow-up study. Respir. Res. 23, 10. https://doi.org/10.1186/s12931-022-01928-8 (2022).
Kim, H. G. et al. Lipoteichoic acid isolated from Lactobacillus plantarum inhibits lipopolysaccharide-induced TNF-alpha production in THP-1 cells and endotoxin shock in mice. J. Immunol. 180, 2553–2561. https://doi.org/10.4049/jimmunol.180.4.2553 (2008).
Bunch, H. et al. BRCA1-BARD1 regulates transcription through modulating topoisomerase IIbeta. Open Biol. 11, 210221. https://doi.org/10.1098/rsob.210221 (2021).
Bunch, H. et al. Transcriptional elongation requires DNA break-induced signalling. Nat. Commun. 6, 10191. https://doi.org/10.1038/ncomms10191 (2015).
Rana, N. K., Singh, P. & Koch, B. CoCl(2) simulated hypoxia induce cell proliferation and alter the expression pattern of hypoxia associated genes involved in angiogenesis and apoptosis. Biol. Res. 52, 12. https://doi.org/10.1186/s40659-019-0221-z (2019).
Tripathi, V. K., Subramaniyan, S. A. & Hwang, I. Molecular and cellular response of co-cultured cells toward cobalt chloride (CoCl(2))-induced hypoxia. ACS Omega 4, 20882–20893. https://doi.org/10.1021/acsomega.9b01474 (2019).
Triantafyllou, A. et al. Cobalt induces hypoxia-inducible factor-1alpha (HIF-1alpha) in HeLa cells by an iron-independent, but ROS-, PI-3K- and MAPK-dependent mechanism. Free Radic. Res. 40, 847–856. https://doi.org/10.1080/10715760600730810 (2006).
Li, Q., Ma, R. & Zhang, M. CoCl(2) increases the expression of hypoxic markers HIF-1alpha, VEGF and CXCR4 in breast cancer MCF-7 cells. Oncol. Lett. 15, 1119–1124. https://doi.org/10.3892/ol.2017.7369 (2018).
Koshiji, M. et al. HIF-1alpha induces cell cycle arrest by functionally counteracting Myc. EMBO J. 23, 1949–1956. https://doi.org/10.1038/sj.emboj.7600196 (2004).
Engeland, K. Cell cycle regulation: p53–p21-RB signaling. Cell Death Differ. 29, 946–960. https://doi.org/10.1038/s41418-022-00988-z (2022).
Madan, E. et al. HIF-transcribed p53 chaperones HIF-1alpha. Nucleic Acids Res. 47, 10212–10234. https://doi.org/10.1093/nar/gkz766 (2019).
Chen, C., Pore, N., Behrooz, A., Ismail-Beigi, F. & Maity, A. Regulation of glut1 mRNA by hypoxia-inducible factor-1. Interaction between H-ras and hypoxia. J. Biol. Chem. 276, 9519–9525. https://doi.org/10.1074/jbc.M010144200 (2001).
Hayashi, M. et al. Induction of glucose transporter 1 expression through hypoxia-inducible factor 1alpha under hypoxic conditions in trophoblast-derived cells. J. Endocrinol. 183, 145–154. https://doi.org/10.1677/joe.1.05599 (2004).
Leiherer, A., Geiger, K., Muendlein, A. & Drexel, H. Hypoxia induces a HIF-1alpha dependent signaling cascade to make a complex metabolic switch in SGBS-adipocytes. Mol. Cell Endocrinol. 383, 21–31. https://doi.org/10.1016/j.mce.2013.11.009 (2014).
Zhang, W. et al. Evidence that hypoxia-inducible factor-1 (HIF-1) mediates transcriptional activation of interleukin-1beta (IL-1beta) in astrocyte cultures. J. Neuroimmunol. 174, 63–73. https://doi.org/10.1016/j.jneuroim.2006.01.014 (2006).
Yang, Y. et al. HIF-1 Interacts with TRIM28 and DNA-PK to release paused RNA polymerase II and activate target gene transcription in response to hypoxia. Nat. Commun. 13, 316. https://doi.org/10.1038/s41467-021-27944-8 (2022).
Multhoff, G., Molls, M. & Radons, J. Chronic inflammation in cancer development. Front. Immunol. 2, 98. https://doi.org/10.3389/fimmu.2011.00098 (2011).
Freeman, H. J. Colorectal cancer risk in Crohn’s disease. World J. Gastroenterol. 14, 1810–1811. https://doi.org/10.3748/wjg.14.1810 (2008).
McGettrick, A. F. & O’Neill, L. A. J. The role of HIF in immunity and inflammation. Cell Metab. 32, 524–536. https://doi.org/10.1016/j.cmet.2020.08.002 (2020).
Lin, N. et al. Myeloid cell hypoxia-inducible factors promote resolution of inflammation in experimental colitis. Front. Immunol. 9, 2565. https://doi.org/10.3389/fimmu.2018.02565 (2018).
Li, Y., Su, J., Li, F., Chen, X. & Zhang, G. MiR-150 regulates human keratinocyte proliferation in hypoxic conditions through targeting HIF-1alpha and VEGFA: Implications for psoriasis treatment. PLoS ONE 12, e0175459. https://doi.org/10.1371/journal.pone.0175459 (2017).
Hsu, C. W. et al. Identification of approved and investigational drugs that inhibit hypoxia-inducible factor-1 signaling. Oncotarget 7, 8172–8183. https://doi.org/10.18632/oncotarget.6995 (2016).
Li, M. & Kim, W. Y. Two sides to every story: The HIF-dependent and HIF-independent functions of pVHL. J. Cell Mol. Med. 15, 187–195. https://doi.org/10.1111/j.1582-4934.2010.01238.x (2011).
Duan, D. R. et al. Inhibition of transcription elongation by the VHL tumor suppressor protein. Science 269, 1402–1406. https://doi.org/10.1126/science.7660122 (1995).
Hoefflin, R. et al. HIF-1alpha and HIF-2alpha differently regulate tumour development and inflammation of clear cell renal cell carcinoma in mice. Nat. Commun. 11, 4111. https://doi.org/10.1038/s41467-020-17873-3 (2020).
Wu, Q. et al. Hypoxia-inducible factors: Master regulators of hypoxic tumor immune escape. J. Hematol. Oncol. 15, 77. https://doi.org/10.1186/s13045-022-01292-6 (2022).
Li, R. L. et al. HIF-1alpha is a potential molecular target for herbal medicine to treat diseases. Drug Des. Dev. Ther. 14, 4915–4949. https://doi.org/10.2147/DDDT.S274980 (2020).
Acknowledgements
We thank C. Jung, M. Seu, and the current members of the Bunch laboratory members at Kyungpook National University (KNU) for their technical assistance and K. Kang at Dankook University for his advice on statistical analyses. H.B. thanks J. Christ, J. Park, and John for their loving encouragement and support throughout the course of this work.
Funding
This research was supported by grants from the National Research Foundation (NRF) of the Republic of Korea (2022R1A21003569) to H.B.
Author information
Authors and Affiliations
Contributions
S.J., N.D., J.L., J.J., D.K., C.R., and H.B. performed qRT-PCR. B.K., J.J., and H.B. carried out immunoblotting. J.J., D.K., and H.B. performed ChIP-qPCR. H.B. and J.J. generated PBS-K8, hK8-L and hK8-FL. N.P. and D.-H.C. carried out FACS. H.K. and D.K.C. provided freeze-dried K8 powder and research materials. H.B. created the hypothesis, designed the experiments, analyzed and curated the data, and wrote and revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jeong, J., Kang, BH., Ju, S. et al. Lactiplantibacillus plantarum K8 lysates regulate hypoxia-induced gene expression. Sci Rep 14, 6275 (2024). https://doi.org/10.1038/s41598-024-56958-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-56958-7
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
