
Abstract
The relapsing fever agent Borrelia hermsii is transmitted by the tick Ornithodoros hermsi. To study the B. hermsii-tick interactions required for pathogen acquisition and transmission we developed an artificial membrane feeding system for O. hermsi nymphs and adults that results in a high percentage of engorgement. This system provides the nutritional requirements necessary for the tick to develop, mate, and produce viable eggs. By inoculating the blood with B. hermsii, we were able to obtain infected ticks for quantitative studies on pathogen acquisition and persistence. These ticks subsequently transmitted the spirochetes to mice, validating this system for both acquisition and transmission studies. Using this feeding method, a mutant of the antigenic variation locus of B. hermsii (Vmp–) that is incapable of persisting in mice was acquired by ticks at equivalent densities as the wild-type. Furthermore, Vmp is not required for persistence in the tick, as the mutant and wild-type strains are maintained at similar numbers after ecdysis and subsequent feeding. These results support the theory that Vmp is an adaptation for mammalian infection but unnecessary for survival within the tick. Interestingly, B. hermsii numbers severely declined after acquisition, though these ticks still transmitted the infection to mice. This procedure reduces animal use and provides a safe, highly controlled and well-contained alternative method for feeding and maintaining O. hermsi colonies. Importantly, this system permits quantitative studies with B. hermsii strains through ingestion during the blood meal, and thus more closely recapitulates pathogen acquisition in nature than other artificial systems.
Similar content being viewed by others
Introduction
Hindle and Merriman, in their 1912 study “The sensory perceptions of Argas persicus (Oken)” observed, “Whilst conducting the experiments on the effect of odorous vapours on both normal ticks and those in which the forelegs had been amputated, we also tried the effect of placing a vessel of hot water beneath the filter paper on which the ticks were walking … many inserted their mouth parts and endeavoured to feed through the filter paper … These results suggested to us a method by means of which ticks might be made to feed on any desired liquid…”1. This manuscript presents the first description and use of artificial membrane feeding of ticks that we are aware of. Subsequently, such feeding systems have been developed for both hard ixodid ticks (reviewed by Gonzalez et al.2) and soft argasid ticks including Ornithodoros species3,4,5,6,7,8,9,10,11,12. Use of artificial membrane feeding chambers has permitted studies on the nutritional requirements of ticks13, the effectiveness of acaracides14,15,16, and the elimination or introduction of microbes to ticks9,17,18,19,20,21.
Other methods have successfully introduced bacteria or viruses into ticks, such as immersion, capillary feeding, and injection into the hemocoel or midgut. Although these techniques have their advantages, they also possess some inherent limitations. In the immersion technique, which submerges ticks in a solution of microbes, Ornithodoros hermsi infection with B. hermsii was relatively low (~ 30%) and the route by which spirochetes enter the arthropod was undetermined22. The authors, however, speculated that the microbes were likely taken up through the oral cavity. Capillary feeding is labor- and time-intensive and often yields limited numbers of infected ticks. The microcapillary injection method can produce wounds that may result in an altered physiological state and this approach requires a technical proficiency by the researcher to reduce tick mortality. A significant issue with all three techniques is that they require subsequent feeding on a vertebrate host for the ticks to proceed through the developmental cycle and to assess microbial persistence within and transmission by the vector.
Artificial membrane feeding systems are another alternative that provides numerous advantages. The need for onsite animal usage is alleviated because commercial sources of inexpensive blood are readily available. The method also recreates the natural route of acquisition of many tick-borne pathogens via blood-feeding and entry into the midgut. Additionally, this process offers safe containment of ticks during the feeding process. Specifically, confining the ticks in a feeding chamber avoids the placement of ticks on animals and minimizes the potential of escape, while simultaneously eliminating the possibility of incurring animal bites. Therefore, the use of artificial membrane feeding chambers for vector studies closely mimics the natural progression of pathogen infection and dissemination throughout the tick.
We are interested in pathogen-tick interactions that occur between Borrelia species that cause relapsing fever and the soft ticks of the Ornithodoros genus. In the U.S., the main agent of relapsing fever is Borrelia hermsii (https://www.cdc.gov/relapsing-fever/distribution/index.html) and is transmitted by the tick Ornithodoros hermsi. Numerous advances in the genetic manipulation and cultivation have made B. hermsii a model organism to study the pathogenesis and ecology of relapsing fever23,24,25,26,27.
The relapsing fever Borrelia have several fascinating biological characteristics, including a vector specificity that permits only a specific tick species to transmit a cognate Borrelia species28. This vector specificity resembles that observed for Leishmania-sandfly interactions that occurs through recognition of specific lectins in the sandfly midgut by unique lipophosphoglycans produced by the appropriate protozoan species29,30. The mechanism for Borrelia-Ornithodoros specificity remains unknown but relates to the ability of the spirochete species to persistently infect the salivary glands of its cognate vector31, in contrast to the midgut interactions that occur between Leishmania and the sandfly. Another intriguing characteristic of the relapsing fever Borrelia is the antigenic variation system for evading the vertebrate immune system and that permits the reoccurring spirochetemias that have earned the disease its name. When B. hermsii is acquired by the tick, the mammalian antigenic system no longer produces a variable major protein (Vmp) and is replaced by the variable tick protein (Vtp)32. When the expression locus for the antigenic variation system was genetically disabled producing a mutant strain designated Vmp–, the mutant was unable to persist in mice25. The regulation of the switching between the Vmp-Vtp loci is not fully understood.
To further our understanding of these and other aspects of B. hermsii–O. hermsi interactions, we developed an artificial membrane feeding system to provide an alternative method for O. hermsi colony maintenance and to introduce the pathogen into the tick. Optimization of the method permitted feeding of adults and later-stage nymphs to repletion, providing all the nutrients necessary for tick development and reproduction. This system was used to introduce both wild-type and the Vmp– mutant strains into O. hermsi to further evaluate the contribution of Vmp during the tick infectious cycle.
Methods
Bacterial strains and growth conditions
Borrelia hermsii WT strain DAH 2E7 passage 3 and the Vmp– mutant25 were cultured in BSKII medium33 at 35 °C under an atmosphere of 5% CO2 and 3% O2 in a Forma Series II Water Jacket CO2 incubator (Thermo Fisher Scientific, Inc., Waltham, MA) with caps loosely attached on the culture tubes.
Ticks and artificial feeding systems
Ticks were obtained from an O. hermsi SIS colony maintained at Rocky Mountain Laboratories that originated from individuals collected from Siskiyou County, CA. Except when being fed, ticks were held in bell jars containing a saturated KCl solution to maintain high humidity levels (~ 85%).
Various membranes and environmental criteria were tested to obtain conditions for efficient engorgement of O. hermsi. The optimized artificial feeding system (Fig. 1) consists of a polycarbonate tube (approximately 2.5 cm inner diameter, and 4.5 cm tall) with Parafilm M stretched tightly across the bottom to act as a feeding membrane and mosquito netting covering the top of the chamber to confine the ticks. The netting was held in place by a tight-fitting rubber O-ring. As a phagostimulant, mouse hair was lightly distributed over the interior surface of the parafilm. Defibrinated bovine or rabbit blood was purchased from HemoStat Laboratories (Dixon, CA, USA) and supplemented with ATP (4 µM) (Sigma-Aldrich, St. Louis, MO, USA) and glucose (2 mg/mL) (Sigma-Aldrich). Blood (4 mL) was aliquoted into each well of a 6-well plate and the chamber, membrane-side down, was set into the blood. A second tight-fitting rubber O-ring on the chamber permitted adjusting the chamber height to position the membrane just below the surface of the blood but above the floor of the plate (Fig. 1c). The feeding system was placed in a dark incubator at 35 °C with 2.5–3.0% CO2 for 1–3 h. After feeding, ticks were returned to the humidity-controlled bell jars until needed.

Artificial feeding system for O. hermsi. (a) The chamber consists of a polycarbonate tube sealed at the bottom with parafilm (that serves as the membrane) and enclosed on the top with mosquito netting. (b) Ticks are safely contained within the chamber and the parafilm membrane is lightly covered with mouse hair that may act as a phagostimulant. (c) The upper rubber O-ring holds the mosquito netting in place while the lower O-ring can be adjusted to maintain the chamber at the proper height in the blood. (d) After incubation, most ticks have fed to repletion.
Infecting ticks with B. hermsii was accomplished by inoculating the blood reservoir prior to insertion of the chambers. In vitro-cultivated B. hermsii were enumerated by darkfield microscopy using a Petroff-Hausser counting chamber (Electron Microscopy Sciences, Hatfield, PA, USA). An actively growing culture of B. hermsii was pelleted by centrifugation and resuspended in blood to achieve a final concentration of 1 × 106–6 × 107 spirochetes/mL.
Quantification of B. hermsii in ticks
Immediately after chamber feeding, B. hermsii densities in individual ticks and from the blood reservoir were determined by colony enumeration in solid medium, as previously described26. Briefly, engorged, infected ticks were surfaced sterilized by washing sequentially in 3% H2O2, 70% ethanol, and sterile water. Ticks were then pulverized using a sterile pestle in a 1.5 mL Eppendorf tube with 0.1 mL BSKII, then raised to 1 mL final volume. Aliquots of this suspension were added to plating medium26 that was allowed to solidify at room temperature prior to incubation at 35 °C in 5% CO2 and 3% O2 using a Forma Series II Water Jacket CO2/O2 incubator (Thermo Fisher Scientific, Inc., Waltham, MA, USA). Plates were incubated for 10–14 days after which colony forming units were counted and total spirochete densities were calculated for each tick and the blood reservoir. Statistical analysis was performed using GraphPad Prism software version 9.1.1 for macOS (GraphPad Software, San Diego, CA, USA).
Tick-mouse transmission studies
Ticks initially infected via the artificial feeding membrane system were held in bell jars containing a saturated solution of potassium chloride. After molting, infected ticks were allowed to feed on female C3SnSmn.Cg-Prkdcscid/J mice (The Jackson Laboratory, Bar Harbor, Maine) to assess the competence of the ticks to transmit B. hermsii to a vertebrate host. Mice were anesthetized with pentobarbital (0.5 mg/10 g body weight) (Abbott Laboratories, North Chicago, IL) via intraperitoneal injection. Three O. hermsi ticks were fed on each of five SCID mice. Mice were monitored on days 4–7 post-infestation for spirochetemia by examining blood samples by thin-smear darkfield microscopy for the presence of motile B. hermsii among the red blood cells. When a spirochetemia was detected, mice were euthanized.
Ethics statement
All animal experiments were conducted at the Rocky Mountain Laboratories, in accordance with the guidelines of the The Rocky Mountain Laboratories’ Institutional Animal Care and Use Committee (RML ACUC) and with the ARRIVE guidelines (https://arriveguidelines.org). Personnel were trained and certified in accordance with the guidelines of the Association for Assessment and Accreditation of Laboratory Animal Care, International and the Office of Laboratory Animal Welfare. Mice were cohoused in HEPA-filtered cages with nesting material, and commercial food and water were available ad libitum. Humane endpoint criteria were specified in the RML ACUC-approved animal study protocols (ASP 2018-061 and 2021-059) to determine when mice should be humanely euthanized.
Results
Optimization of an artificial membrane feeding system for O. hermsi
Multiple variables were tested in the development of an efficient feeding chamber for O. hermsi ticks, including membrane composition, environmental conditions, and kairomones and phagostimulants (Table 1). In all, 92 of 134 ticks (69%) in 11 trials fed to repletion on the artificial membrane (Table 1). The most efficient conditions for tick engorgement were obtained using parafilm M membranes with the addition of mouse hair, glucose and ATP in the blood, and incubation at 35 °C and 5% CO2. Although we did not assess the individual requirement of each component, higher feeding efficiencies were obtained with these additions. ATP is a phagostimulant for inducing tick feeding34,35, while hair6 and CO2 may act as kairomones. Ticks fed through both silicone-based and parafilm M membranes, but the latter is easier and cheaper to use and produced consistently high proportions of engorgement. When used with these conditions, the artificial membrane feeding chambers (Fig. 1) routinely induced 80–92% of the ticks to feed to repletion (Table 1). Usually, 100% of the engorged ticks survived through ecdysis.
Nymphs that fed through the artificial membranes molted and developed to the next life stage, while adults that fed in this manner mated and produced viable eggs (Table 1). Therefore, this system provides all the necessary nutritional requirements for ticks to progress through the lifecycle from a nymph to the adult and to reproduce. However, we were unsuccessful at inducing the larvae and first-stage nymphs to feed under any conditions tested (data not shown).
O. hermsi can acquire B. hermsii via artificial membrane feeding
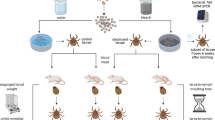
The mouse-tick infection cycle of B. hermsii can be accurately modeled with laboratory mouse strains and tick colonies. However, the ability to study tick-specific phenotypes using this lab model is compromised when studying mutants incapable of infecting mice25,36. Artificial membrane feeding chambers provide a useful method of producing pathogen-infected ticks via the route of blood-feeding9,17,18,19,21. Figure 2 shows the experimental strategy used in this study. The genetically engineered Vmp– mutant, is unable to undergo antigenic variation and does not cause relapses in mice, making it difficult to obtain infected ticks by feeding on inoculated mice25.

Overview of experimental strategy. (a) O. hermsi ticks were placed in the artificial membrane chambers and (b) allowed to feed on infected blood. (c) A portion of the fed ticks were pulverized in liquid medium and diluted into solid medium to calculate spirochete densities. (d) The remaining ticks were allowed to molt and then to feed on naïve mice, after which B. hermsii transmission was assessed by examining mouse blood for spirochetemias (e).
Using the artificial membrane feeding chambers we infected O. hermsi ticks with both the WT and Vmp– strains of B. hermsii. In vitro cultivated spirochetes were inoculated into the blood reservoir and ticks were allowed to feed to engorgement. After feeding was completed, aliquots from the blood and from 2 to 3 crushed ticks from each chamber were added to plating medium to confirm spirochete concentrations. Spirochete concentrations remaining in the blood varied between 1 × 106 and 6 × 107 B. hermsii/mL, depending on the experiment, a concentration range observed in the blood of spirochetemic mice25,37,38. These results demonstrated that the Vmp– mutant was acquired by ticks at comparable densities to that of WT (Fig. 3), thus confirming that Vmp does not contribute to acquisition of B. hermsii by O. hermsi ticks25.

Numbers of B. hermsii spirochetes in O. hermsi ticks post-feeding. Immediately after feeding through artificial membranes on infected blood, individual ticks were crushed and spirochete densities determined by colony count on solid medium. No significant difference was observed between acquisition of the WT strain and the Vmp– mutant as calculated by an unpaired, 2-tailed T-test. Each symbol represents B. hermsii densities within an individual tick, and differences in shading indicates independent feeding experiments (3 total). Bars represent the mean ± the standard error of the mean.
Membrane-fed ticks maintain and transmit B. hermsii
After molting to late-stage nymphs or adults, we used O. hermsi previously infected by blood-feeding through the artificial membrane chambers to assess their ability to transmit B. hermsii to mice. Because the Vmp– mutant does not relapse or reach the densities of WT B. hermsii in immunocompetent mice25, we used C3SnSmn.Cg-Prkdcscid/J (SCID) mice. The lack of a humoral immune response in SCID mice permits relapsing fever spirochetes to attain persistently high densities, making infection easier to detect by darkfield microscopy for B. hermsii in the blood.
In a typical trial, three ticks were placed on individual SCID mice and allowed to feed to repletion; infection was then affirmed by darkfield microscopy for the presence of spirochetes in the blood (Fig. 4). All six mice fed upon by WT-infected ticks became spirochetemic, but only two of four SCID mice total became infected with the Vmp– strain. The two mice that did not become infected were fed upon by one or two ticks only. Hence, consistent infection of SCID mice required 3 ticks per animal, and was less successful when 1–2 infected ticks fed per mouse. Both B. hermsii strains were transstadially maintained by O. hermsi ticks infected by feeding through artificial membrane chambers and transmitted the infection to SCID mice. Therefore, this model system mimics the natural tick-mouse infectious cycle.

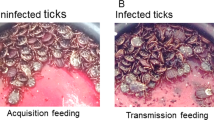
Transmission of B. hermsii by tick bite. (a) Three infected ticks (yellow arrows) were allowed to feed to repletion on individual SCID mice. The boxed area is enlarged (b) to show a feeding O. hermsi tick. (c) Spirochete transmission to the mice was confirmed by visualizing the spirochetes among the red blood cells by thin smear. To enhance visualization of B. hermsii in blood, an inverse image was used and spirochetes were highlighted in red using Adobe Photoshop. Representative images are shown.
Interestingly, B. hermsii densities within ticks were very low following the molt and subsequent feeding (Fig. 5), whether the ticks were fed or unfed, compared to initial densities of spirochetes detected in ticks after acquisition (Fig. 3). This indicates that the reduction in spirochete numbers was not due to effects from the subsequent bloodmeal but to changes occurring either during digestion of the initial bloodmeal or during the molt. Both the WT and Vmp– mutant strains had similar declines and were not significantly different from each other.

B. hermsii densities within the tick decrease following acquisition. Ticks that had acquired the infection by feeding through an artificial membrane were sampled after molting (WT Unfed), or after a subsequent bloodmeal on uninfected SCID mice (WT Fed and Vmp– Fed). Individual ticks were crushed and spirochete densities determined by colony count on solid medium. Each symbol represents B. hermsii densities within an individual tick. No significant difference was observed between WT Fed and the other two samples, as calculated by an unpaired, 2-tailed T-test. Results from two independent feeding experiments are shown. Bars represent the mean ± the standard error of the mean.
Discussion
Ornithodoros ticks are widespread globally, native to both the old world and the new. Some of these tick species have evolved a unique symbioses with specific members of the relapsing fever clade of Borrelia, and new symbiotic species are still being identified39. Occurring in Africa, Asia, Europe and the Americas, relapsing fever produces flu-like symptoms including fever, nausea, chills, aches, and fatigue40. More severe clinical features include diarrhea, respiratory issues, neurological complications that can result in death, and in utero infections leading to abortions or still births.
The ability of relapsing fever Borrelia to evade the host immune system and cause relapses is due to the antigenic variation of the Vmp proteins, although in most cases the host eventually clears the pathogen41. A mutant incapable of expressing Vmp is attenuated for mammalian infection and rapidly cleared by the host25. Identification of mutants attenuated for host infection (e.g. Vmp–) are critical to understanding the mechanisms of disease. However, since they are rapidly cleared and do not persist in vertebrate hosts, it is intrinsically problematic to study the phenotype within O. hermsi ticks. Consequently, artificial feeding chambers would provide a useful method to obtain infected ticks. Although these mutants can be introduced to the tick by multiple methods, as discussed in the Introduction, none recapitulate the natural acquisition route of the pathogen by ingestion with a blood meal except artificial membrane chambers. Mammalian hosts develop relapsing fever by transmission from an infected arthropod. Therefore, an artificial feeding system for O. hermsi provides the ability to characterize relapsing fever strains at the critical points of acquisition and transmission by the tick. A widespread effort has generated artificial feeding systems for a variety of Ornithodoros species, but surprisingly not for O. hermsi.
A comparison of artificial feeding systems described for other Ornithodoros ticks identifies subtle but perhaps significant differences in protocols. For example, O. coriaceus were only induced to feed when mammal hair was placed on the surface of the membrane5, while the addition of rabbit ear wax to the membrane produced better results for O. moubata and O. tholozani3. In a separate study O. moubata ticks had a higher feeding efficiency with elevated temperatures (40° vs. 27 °C degrees)10. Other differences in feeding chambers included the membrane material (parafilm vs. silicone) and the addition of phagostimulants such as glucose, ATP, and glutathione34,35. These requirements may relate to differences in the biology of the Ornithodoros species being studied, or, alternatively, may reflect the peculiarities of lab-to-lab variation in technique. By adopting some of these phagostimulants and optimizing conditions we were able to induce later-stage nymphs and adult O. hermsi ticks to feed to engorgement in an artificial membrane chamber at a high efficiency (Table 1 and Fig. 1). The critical features for enticing the later stages of O. hermsi to feed appear to be the use of parafilm as the membrane material and incubation at 35 °C in a CO2 incubator (Fig. 2). Although adults fed well in a single experiment in a 37 °C water bath (Table 1), the results were inconsistent with nymphs, which only reached reproducibly high proportions of feeding under CO2 at 35 °C. The source of the blood (rabbit or bovine) did not seem to matter and adult ticks that fed on either produced viable eggs. Other kairomones and phagostimulants, such as the addition of mouse hair to the membrane and the inclusion of ATP and glucose in the blood, were not assessed individually for their necessity, but these conditions routinely yielded high repletion percentages (80–92%). The efficiency of tick feeding by this method is equivalent to that of ticks feeding on mice reported by McCoy et al. for nymphs and adults (79–92%)38.
One limitation to this method is the failure of larvae or 1st stage nymphs to feed. Possibly their mouthparts are too small to pierce the parafilm membrane, though the measured thickness of the stretched parafilm in our system is approximately 39 µm (standard deviation ± 9.4, N = 7 independent measurements). The reported length of O. hermsi larval hypostomes ranges from 62 to ~ 80 µm42,43 indicating that the mouthparts of these immature stages should be long enough to cross the parafilm. Alternatively, O. hermsi are relatively small ticks, even compared to other Ornithodoros species, and perhaps the tensile strength of the parafilm poses to great of a resistance to the forces that can be generated by the cheliceral digits of these small ticks. Of the Ornithodoros artificial feeding systems surveyed in the literature, only two described limited success with feeding larvae of O. turicatae and O. puertoricensis4,7. Neither paper noted significant changes to their protocol to stimulate larval feeding compared to that of the nymph or adult, but both had reduced success with larval feeding. Possibly the longer hypostomes of these species compared to that of O. hermsi42,43 permit them to generate the force necessary to penetrate the artificial membrane. We have been unable to definitively determine why the early stages of O. hermsi do not feed efficiently through artificial membranes.
The success with feeding of O. hermsi ticks encouraged us to assess pathogen acquisition using the feeding chambers. Of particular interest was whether we could use this system to infect ticks with mutant strains of B. hermsii attenuated for mammalian infection. Raffel et al. induced an artificial spirochetemia in a single mouse by injecting 1.5 × 108 Vmp– spirochetes and permitting ticks to feed before the mutant strain was cleared, demonstrating that Vmp was not necessary for either acquisition or persistence in the tick25. Using artificial membrane chambers we confirmed this finding by feeding O. hermsi on blood inoculated with either WT or Vmp– B. hermsii strains. The densities of spirochetes detected in the blood meal ranged from 1 × 106 to 6 × 107 B. hermsii/mL, comparable to densities observed in spirochetemic mice25,37,38. We quantified spirochete numbers from the engorged ticks after feeding and demonstrated that both strains were acquired at equivalent densities by ticks (Fig. 3). McCoy et al. reported the blood volume ingested by O. hermsi 3rd stage nymphs and females averaged approximately 8 µL and 16 µL of blood, respectively38. Using these figures, we would predict that a female engorged on blood at a concentration of 1 × 107 bacteria/mL would, on average, imbibe 1.6 × 105 spirochetes (1 × 107 bacteria/mL × 0.016 ml). The mean densities of B. hermsii WT and Vmp– strains in ticks infected through artificial membrane feeding determined in this study were 1.2 × 105 and 1.6 × 105, respectively (Fig. 3), as predicted based on the measurements of blood imbibed by B. hermsii38.
Ticks infected with either B. hermsii strain successfully transmitted the infection to SCID mice and produced a spirochetemia (Fig. 4). Thus, ticks that had acquired B. hermsii through feeding on artificial membranes maintained the spirochetal infection through the molt and successfully transmitted the spirochetes to mice. When three infected ticks successfully fed on a mouse, all mice became infected. However, when a mouse was fed on by one or two ticks the mouse failed to become infected. These results are in agreement with those of Boyle et al., who demonstrated that three O. turicata ticks were sufficient to transmit B. turicatae to most mice in their study, whereas one or two ticks per mouse rarely transmitted the infection44. Davis and Walker reported slightly higher success with 65% of individual O. hermsi ticks transmitting B. hermsii45.
Alternatively, the lower number of infected SCID mice by the Vmp– strain may result from the leaky B cell response that can occur in some SCID mice, which may impact the mutant strain more than the WT46. Regardless of the cause, our results indicate that three ticks per mouse were sufficient to transmit B. hermsii to all mice in this study. However, the spirochete densities in these ticks were substantially lower after the molt than at acquisition (Fig. 5 compared to Fig. 3). The reduction in spirochete numbers from the acquisition stage to the transmission stage was > 99%. As noted above, despite the decline in spirochete numbers, three ticks per mouse was still sufficient to ensure transmission.
The low densities of B. hermsii in ticks was a surprising result, although McCoy et al. noted an approximate 86% reduction of B. hermsii numbers in the five days following acquisition feeding38. In the Lyme disease spirochete that is vectored by hard ticks of the Ixodes genus, B. burgdorferi populations reach their highest densities shortly after the bloodmeal, but decline tenfold when assessed after the molt47. Our observation of spirochete densities decreasing over the lifespan of membrane-fed ticks dictates that future population dynamics studies be conducted comparing B. hermsii numbers within ticks that have fed upon mice versus those fed in membrane chambers.
In summary, the artificial membrane feeding chambers provide a new method for O. hermsi maintenance and insights into tick-microbe interactions. Adult ticks infected with WT or the Vmp– mutant using this technique went on to produce viable egg clutches (Table 1), indicating that this artificial feeding system provides all necessary requirements for O. hermsi to develop through its life cycle. The observations of the B. hermsii Vmp– mutant reinforces the evidence25 that Vmp is not required for survival in the tick but as an adaptation for mammalian infection. The artificial membrane feeding system for O. hermsi can be used for studies on pathogen transmission (e.g. number of microbes transmitted during feeding) and also for transstadial and transovarial transmission experiments. These chambers provide an efficient and inexpensive method for rearing O. hermsi ticks from second stage nymphs through adults that produce viable larvae, thereby significantly reducing the requirement for live animals. Additionally, this system affords a simple method to infect ticks with B. hermsii mutants that are incapable of persisting in mammals and opens new avenues for investigating the mechanism of vector specificity displayed by tick-borne relapsing fever spirochetes.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Hindle, E. & Merriman, G. The sensory perceptions of Argas persicus (Oken). Parasitology 5, 203–216 (1912).
Gonzalez, J., Bickerton, M. & Toledo, A. Applications of artificial membrane feeding for ixodid ticks. Acta Trop. 215, 105818. https://doi.org/10.1016/j.actatropica.2020.105818 (2021).
Ben-Yakir, D. & Galun, R. Comparative study of two argasid tick species: feeding response to phagostimulants. Israel J. Ecol. Evol. 39, 169–176 (1993).
Butler, J. F., Hess, W. R., Endris, R. G. & Holscher, K. H. In vitro feeding of Ornithodoros ticks for rearing and assessment of disease transmission. In Acarology VI Vol. 2 (eds Griffiths, D. A. & Bowman, C. D.) 1075–1081 (Ellis Horwood Ltd., 1984).
Hokama, Y., Lane, R. S. & Howarth, J. A. Maintenance of Adult and Nymphal Ornithodoros coriaceus (Acari: Argasidae) by artificial feeding through a parafilm membrane. J. Med. Entomol. 24, 319–323 (1987).
Howarth, J. A. & Hokama, Y. Studies with Ornithodoros coriaceus Koch (Acarina: Argasidae), the suspected vector of Epizootic Bovine Abortion. In Tick-Borne Diseases and Their Vectors (ed. Wilde, J. K. H.) 169–174 (Centre for Tropical Veterinary Medicine, University of Edinburgh, 1978).
Kim, H. J., Filatov, S., Lopez, J. E., Pérezdeleón, A. A. & Teel, P. D. Blood feeding of Ornithodoros turicata larvae using an artificial membrane system. Med. Vet. Entomol. 31, 230–233. https://doi.org/10.1111/mve.12223 (2017).
Lewis, C. E., Bartholomay, L. C. & Blanchong, J. A. Feeding of adult Ornithodoros tartakovskyi ticks using a modified artificial membrane feeding system. Med. Vet. Entomol. 34, 123–126. https://doi.org/10.1111/mve.12398 (2020).
Neelakanta, G., Sultana, H., Sonenshine, D. E. & Marconi, R. T. An in vitro blood-feeding method revealed differential Borrelia turicatae (Spirochaetales: Spirochaetaceae) gene expression after spirochete acquisition and colonization in the soft tick Ornithodoros turicata (Acari: Argasidae). J. Med. Entomol. 54, 441–449. https://doi.org/10.1093/jme/tjw171 (2017).
Osborne, R. W. & Mellor, P. S. Use of a silicone membrane feeding technique in laboratory maintenance of a colony of Ornithodoros moubata. Trop. Anim. Health Prod. 17, 31–38 (1985).
Tarshis, B. A preliminary study on feeding Ornithodoros savignyi (Audouin) on human blood through animal-derived membranes (Acarina: Argasidae). J. Exp. Biol. 24, 41–61. https://doi.org/10.1242/jeb.24.1-2.41 (1947).
Zheng, H. et al. Biological and physiological characterization of in vitro blood feeding in nymph and adult stages of Ornithodoros turicata (Acari: Argasidae). J. Insect Physiol. 75, 73–79. https://doi.org/10.1016/j.jinsphys.2015.03.005 (2015).
Perner, J. et al. Acquisition of exogenous haem is essential for tick reproduction. eLife https://doi.org/10.7554/eLife.12318 (2016).
Kuhnert, F. Feeding of hard ticks in vitro: new perspectives for rearing and for the identification of systemic acaricides. Altex 13, 76–87 (1996).
Krober, T. & Guerin, P. M. An in vitro feeding assay to test acaricides for control of hard ticks. Pest Manag. Sci. 63, 17–22. https://doi.org/10.1002/ps.1293 (2007).
Krober, T. & Guerin, P. The tick blood meal: from a living animal or from a silicone membrane? ALTEX 24 Spec No, 39–41 (2007).
Korner, S. et al. Uptake and fecal excretion of Coxiella burnetii by Ixodes ricinus and Dermacentor marginatus ticks. Parasit. Vectors 13, 75. https://doi.org/10.1186/s13071-020-3956-z (2020).
Migne, C. V. et al. Evaluation of two artificial infection methods of live ticks as tools for studying interactions between tick-borne viruses and their tick vectors. Sci. Rep. 12, 491. https://doi.org/10.1038/s41598-021-04498-9 (2022).
Oliver, J. D. et al. Infection of immature Ixodes scapularis (Acari: Ixodidae) by membrane feeding. J. Med. Entomol. 53, 409–415. https://doi.org/10.1093/jme/tjv241 (2016).
Oliver, J. D. et al. Growth dynamics and antibiotic elimination of symbiotic Rickettsia buchneri in the tick Ixodes scapularis (Acari: Ixodidae). Appl. Environ. Microbiol. https://doi.org/10.1128/AEM.01672-20 (2021).
Vimonish, R. et al. Quantitative analysis of Anaplasma marginale acquisition and transmission by Dermacentor andersoni fed in vitro. Sci. Rep. 10, 470. https://doi.org/10.1038/s41598-019-57390-y (2020).
Policastro, P. F., Raffel, S. J. & Schwan, T. G. Cotransmission of divergent relapsing fever spirochetes by artificially infected Ornithodoros hermsi. Appl. Environ. Microbiol. 77, 8494–8499. https://doi.org/10.1128/AEM.05830-11 (2011).
Fine, L. M., Earnhart, C. G. & Marconi, R. T. Genetic transformation of the relapsing fever spirochete Borrelia hermsii: stable integration and expression of green fluorescent protein from linear plasmid 200. J. Bacteriol. 193, 3241–3245. https://doi.org/10.1128/JB.05037-11 (2011).
Policastro, P. F. & Schwan, T. G. Experimental infection of Ixodes scapularis larvae (Acari: Ixodidae) by immersion in low passage cultures of Borrelia burgdorferi. J. Med. Entomol. 40, 364–370 (2003).
Raffel, S. J., Battisti, J. M., Fischer, R. J. & Schwan, T. G. Inactivation of genes for antigenic variation in the relapsing fever spirochete Borrelia hermsii reduces infectivity in mice and transmission by ticks. PLoS Pathog. 10, e1004056. https://doi.org/10.1371/journal.ppat.1004056 (2014).
Raffel, S. J., Williamson, B. N., Schwan, T. G. & Gherardini, F. C. Colony formation in solid medium by the relapsing fever spirochetes Borrelia hermsii and Borrelia turicatae. Ticks Tick Borne Dis. 9, 281–287. https://doi.org/10.1016/j.ttbdis.2017.11.001 (2018).
Battisti, J. M., Raffel, S. J. & Schwan, T. G. A system for site-specific genetic manipulation of the relapsing fever spirochete Borrelia hermsii. in Bacterial Pathogenesis Methods and Protocols Vol. 431 (eds F. R. DeLeo & M. Otto) 69–84 (Humana Press, 2008).
Davis, G. E. Species unity or plurality of the relapsing fever spirochetes. Am. Assoc. Adv. Sci. Monogr. 18, 41–47 (1942).
Pimenta, P. F. et al. Evidence that the vectorial competence of phlebotomine sand flies for different species of Leishmania is controlled by structural polymorphisms in the surface lipophosphoglycan. Proc. Natl. Acad. Sci. U.S.A. 91, 9155–9159 (1994).
Sacks, D. L., Pimenta, P. F., McConville, M. J., Schneider, P. & Turco, S. J. Stage-specific binding of Leishmania donovani to the sand fly vector midgut is regulated by conformational changes in the abundant surface lipophosphoglycan. J. Exp. Med. 181, 685–697 (1995).
Schwan, T. G. Vector specificity of the relapsing fever spirochete Borrelia hermsii (Spirochaetales: Borreliaceae) for the tick Ornithodoros hermsi (Acari: Argasidae) involves persistent infection of the salivary glands. J. Med. Entomol. 58, 1926–1930. https://doi.org/10.1093/jme/tjab060 (2021).
Schwan, T. G. & Hinnebusch, B. J. Bloodstream- versus tick-associated variants of a relapsing fever bacterium. Science 280, 1938–1940 (1998).
Barbour, A. G. Isolation and cultivation of Lyme disease spirochetes. Yale J. Biol. Med. 57, 521–525 (1984).
Galun, R. & Kindler, S. H. Glutathione as an inducer of feeding in ticks. Science 147, 166–167. https://doi.org/10.1126/science.147.3654.166 (1965).
Galun, R. & Kindler, S. H. Chemical basis of feeding in the tick Omithodoros tholozani. J. Insect Physiol. 14, 1409–1421 (1968).
Guyard, C. et al. Periplasmic flagellar export apparatus protein, FliH, is involved in post-transcriptional regulation of FlaB, motility and virulence of the relapsing fever spirochete Borrelia hermsii. PLoS ONE 8, e72550. https://doi.org/10.1371/journal.pone.0072550 (2013).
Crowder, C. D. et al. Pathogen and host response dynamics in a mouse model of Borrelia hermsii relapsing fever. Vet. Sci. https://doi.org/10.3390/vetsci3030019 (2016).
McCoy, B. N., Raffel, S. J., Lopez, J. E. & Schwan, T. G. Bloodmeal size and spirochete acquisition of Ornithodoros hermsi (Acari: Argasidae) during feeding. J. Med. Entomol. 47, 1164–1172. https://doi.org/10.1603/me10175 (2010).
Bermudez, S. E. et al. Isolation and genetic characterization of a relapsing fever spirochete isolated from Ornithodoros puertoricensis collected in central Panama. PLoS Negl. Trop. Dis. 15, e0009642. https://doi.org/10.1371/journal.pntd.0009642 (2021).
Warrell, D. A. Relapsing fevers. In Infectious diseases (eds Cohen, J. et al.) 1105–1109 (Elsevier, 2017).
Barbour, A. G., Dai, Q., Restrepo, B. I., Stoenner, H. G. & Frank, S. A. Pathogen escape from host immunity by a genome program for antigenic variation. Proc. Natl. Acad. Sci. U.S.A. 103, 18290–18295. https://doi.org/10.1073/pnas.0605302103 (2006).
Cooley, R. A. & Kohls, G. M. The Argasidae of North America, Central America and Cuba. The American Midland Naturalist, Monograph No. 1 (1944).
Kohls, G. M., Sonenshine, D. E. & Clifford, C. M. The systematics of the subfamily Ornithodorinae (Acarina: Argasidae). II. Identification of the larvae of the Western Hemisphere and descriptions of three new species. Ann. Entomol. Soc. Am. 58, 331–364. https://doi.org/10.1093/aesa/58.3.331 (1965).
Boyle, W. K., Wilder, H. K., Lawrence, A. M. & Lopez, J. E. Transmission dynamics of Borrelia turicatae from the arthropod vector. PLoS Negl. Trop. Dis. 8, e2767. https://doi.org/10.1371/journal.pntd.0002767 (2014).
Davis, G. E. & Walker, M. E. Ornithodorus hermsi: feeding and molting habits in relation to the acquisition and transmission of relapsing fever spirochetes. Publ. Health Rep. 55, 492–504 (1940).
Bosma, G. C. et al. Evidence of functional lymphocytes in some (leaky) scid mice. J. Exp. Med. 167, 1016–1033. https://doi.org/10.1084/jem.167.3.1016 (1988).
Piesman, J., Oliver, J. R. & Sinsky, R. J. Growth kinetics of the Lyme disease spirochete (Borrelia burgdorferi) in vector ticks (Ixodes dammini). Am. J. Trop. Med. Hyg. 42, 352–357 (1990).
Acknowledgements
We thank Tom Schwan for sharing his expertise on Ornithodoros hermsi rearing and biology, and critical reading of the manuscript. We also thank Dan Sonenshine for advice and insight on artificial membrane feeding of ticks. We are grateful for the graphical expertise of Austin Athman, Ryan Kissinger, Anita Mora and Rose Perry-Gottschalk for figure preparation. This work was supported by the Division of Intramural Research, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, MD, USA.
Funding
Open Access funding provided by the National Institutes of Health (NIH).
Author information
Authors and Affiliations
Contributions
P.E.S. and S.J.R. performed experiments and analyzed data. P.E.S. prepared figures and wrote the manuscript. S.J.R., F.C.G., and M.E.B. edited the manuscript and figures. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Stewart, P.E., Raffel, S.J., Gherardini, F.C. et al. Kinetics of tick infection by the relapsing fever spirochete Borrelia hermsii acquired through artificial membrane feeding chambers. Sci Rep 12, 13479 (2022). https://doi.org/10.1038/s41598-022-17500-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-17500-9
This article is cited by
-
Experimental transmission of a novel relapsing fever group Borrelia harbored by Ornithodoros octodontus (Ixodida: Argasidae) in Chile
Experimental and Applied Acarology (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
