
Abstract
Self-organizing three-dimensional cellular models derived from human pluripotent stem cells or primary tissue have great potential to provide insights into how the human nervous system develops, what makes it unique and how disorders of the nervous system arise, progress and could be treated. Here, to facilitate progress and improve communication with the scientific community and the public, we clarify and provide a basic framework for the nomenclature of human multicellular models of nervous system development and disease, including organoids, assembloids and transplants.
Similar content being viewed by others

Main
One of the most striking scientific advancements in the stem cell field has been the uncovering of the remarkable potential for self-organization during human pluripotent stem cell differentiation in a laboratory setting1,2,3,4,5. This generally involves creating three-dimensional (3D) aggregates of pluripotent stem cells and growing them in extracellular matrices and/or free-floating conditions to differentiate into self-organizing, millimetre-sized, cellular aggregates that recapitulate some features of a tissue of interest. These advances have been particularly pertinent in neuroscience, a field of study in which access to functional brain tissue is restricted and the need to examine, probe and manipulate human neurons and glial cells from human individuals is essential for revealing the complex biology underlying nervous system development and diseases6,7,8. With the enormous interest in human cellular models of neural development and the rapid pace of progress, many different names and classifications have been used to designate the various 3D tissue-mimicking constructs. The lack of a clear consensus to name these new structures makes it challenging for researchers (even for those active in the field and for trainees) and the public to follow and to clearly delineate technological advances and needs.
We gathered a group of researchers who have participated in the Cold Spring Harbor Laboratory meeting on 3D modelling of human brain to clarify the nomenclature and provide a working set of guidelines for defining 3D cellular models of the nervous system. Following a series of virtual discussions integrating the work performed in many laboratories, we propose the following broad and flexible classification (Fig. 1 and Box 1), as well as a simple set of guidelines for naming that is rooted in developmental neuroanatomy. We hope this effort will facilitate communication and scientific advancement, promote collaborations, and initiate the development of quality control measures and benchmarking in this exciting field.

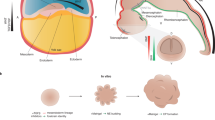
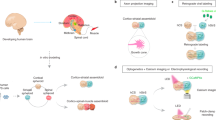
Human pluripotent stem cells (hPSCs; induced or embryonic) can be differentiated in monolayer (2D) conditions to derive the main lineages of the nervous system, and those can be used in isolation or combined as co-cultures, including as part of on-a-chip preparations. Alternatively, human pluripotent stem cells can be differentiated in self-organizing 3D cultures to derive unguided neural organoids or regionalized neural organoids resembling regions or domains of the nervous system. These can be combined to generate assembloids or can be transplanted into animals (grafted or transplanted organoids). Assembloids can be generated from organoids derived from different individuals, which create inter-individual assembloids, or different species, to create inter-species assembloids.
Defining key concepts
We focus our discussion on three main classes of 3D models that are used in the field at present: organoids, assembloids and grafted organoids. We define organoids as in vitro-generated cellular systems that emerge by self-organization, include multiple cell types, and exhibit some cytoarchitectural and functional features reminiscent of an organ or organ region. Organoids can be generated as 3D cultures or by a combination of 3D and 2D approaches (also known as 2.5D) that can develop and mature over long periods of time (months to years). For the nervous system, they are generally constructed from pluripotent stem cells but can also be derived from donor tissues with growth potential (such as glioblastoma organoids). The term organoids encompasses previously used terms such as SFEBq, neuroloids and neural spheroids, if the structures are derived from pluripotent stem cells or from primary tissue stem cells and they include self-organized structures reminiscent of an organ or organ region. The term spheroids can be used to describe a cellular system obtained by combining in 3D culture one or more separately patterned cell types that have limited self-organization properties. By contrast, in organoids, the cells co-develop through self-organizing and interaction, resulting in cells that differ from mixing separately generated cells. Engineered 3D cultures obtained by bioprinting that enable and/or sustain self-organization would fall under the umbrella term organoids. However, if postmitotic neurons are put together in 3D and there is minimal re-organization afterwards, we would not consider this 3D culture an organoid.
We refer to nervous system organoids by the major anatomic region they model, such as cortical organoids, retinal organoids, hypothalamic organoids or spinal cord organoids; the terms neural organoids and nervous system organoids each encompass a broader spectrum of cell types, including those from the central and the peripheral, autonomic and enteric nervous systems, and are appropriate as blanket terms or to describe organoids encompassing several regionally diverse cell types. We define assembloids as self-organizing cellular systems resulting from the combination of a type of organoids with another type of organoids (for example, dorsal forebrain with ventral forebrain) or with different specialized cell types (for example, cortical organoid with endothelial cells) that result in integration9. We refer to organoids or assembloids that have been transplanted10 into animals as grafted organoids or grafted assembloids.
A working classification
We recognize two main categories of neural organoids on the basis of the level of guidance provided during the differentiation of pluripotent stem cells into organoids. By guidance we mean the addition of small molecules or factors intended to generate a particular region or collection of cell types. The term guided is preferred over directed, patterned or specified because exogenous factors are used to guide self-organization and self-patterning in 3D cultures towards developing into a region of the nervous system, and the terms patterned or specified have been used in the context of 2D cultures in which factors are used to direct pluripotent cells towards a specific set of cell fates. If, however, differentiation is unguided, which leads to organoids with a high diversity of neural cells typically representing different locations in the neural axis as well as non-neural derivatives, we refer to them as unguided neural organoids. If organoids are obtained by selecting parts of organoids by using genetic reporters, cell sorting or mechanical dissection (for example, germinal-like zones or optic cup-like structures), we refer to these as selected neural organoids. By contrast, when instructive signals are used to guide the generation of organoids composed of more regionally specific cell types, we call them regionalized neural organoids, which can recapitulate cellular, molecular or anatomical features of particular domains of the nervous system. For instance, hypothalamic organoids resemble part of the hypothalamus, retinal organoids recapitulate features of the retina, telencephalic organoids encompass the telencephalon or forebrain, and cerebral cortical organoids resemble part of the dorsal forebrain.
Organoids can be used to form assembloids (assembled organoids), enabling functional modelling of processes such as neuro-immune interactions, cell migration and circuit assembly. The combination of organoids with other organoids or with different specialized cell types in self-organizing 3D cultures results in emergent features of the system, and the term assembloids is preferred over fused organoids or integrated organoids. Assembloids are obtained by: incorporating different organoids into multi-region assembloids, such as when combining dorsal with ventral forebrain, cerebral cortex with striatum, or cerebral cortex with thalamus; integrating neural or non-neural cells, such as microglia, immune cells and endothelial cells, into organoids to create multi-lineage assembloids; or combining organizer-like cells with organoids (for example, cells producing morphogens such as SHH). Assembloids can be generated from organoids derived from different individuals, to create inter-individual assembloids, or different species, to create inter-species assembloids. Similarly, individual organoids can be generated from mixtures of cell lines derived from different species to create inter-species organoids. Organoids composed of cells from different individuals or that combine mutant and wild-type cell lines are mosaic organoids. We propose that organoids transplanted into animals are referred to as grafted organoids. By contrast, the term chimaeric animals should be reserved for integration of human cells into early embryonic stages of animal development, including following the depletion of a part of the host’s nervous system. These early-stage manipulations may result in broader integration of human cells into the host animal compared to a grafted organoid.
It was felt strongly by all authors that the names mini-brain, brain-in-a-dish and whole-brain organoid to describe neural organoids should be avoided. These terms do not accurately inform the type of organoid produced, and they have proved to be confusing to the layperson and to the scientific community11. Similarly, the term humanized animals to describe animal models with neural organoid-derived grafts should be avoided as it raises concerns about conferring advanced cognitive and behavioural abilities.
General recommendations
To facilitate the current and upcoming nomenclature framework, we propose the following guidelines to promote clarity and rigour. Neural organoids should be named as broadly as possible unless strong evidence for recapitulating a specific domain is provided (for example, spinal, hindbrain or arcuate nucleus). The term cerebral organoid has sometimes been used broadly in the field, but it should be restricted to organoids that predominantly contain cells of the cerebrum. Claiming bona fide identity of in vitro-derived organoids or assembloids requires rigorous benchmarking to primary human tissue12,13. This is often challenging as a comprehensive atlas of the developing human nervous system does not yet exist, so more conservative nomenclature should be adopted when in vivo benchmarking proves difficult. Moreover, specific neuronal or glial cell types are characterized by distinct morphology, location, physiological properties or connectivity. Therefore, a more descriptive naming convention is preferred in the absence of demonstrated features of those cells or regions. For instance, if such evidence is lacking, organoids should be defined by region along axes (for example, ventral–dorsal or anterior–posterior) and by specifying their cell composition (for example, the proportion of GABAergic or glutamatergic neurons). We recommend thoughtfulness in labelling cell types, taking into account their gene expression profiles, electrophysiological properties, activity-dependent states, areal identities and developmental trajectories. Culture conditions that may affect cell states and differentiation capacity (for example, use of serum, commercial kits of unknown composition and uncharacterized cellular substrates), the cell type of origin, the use of exogenous extracellular matrices and the defining features and cellular components used to generate the organoid or assembloid should be provided in detail in scientific publications. If possible, commercial components of unknown composition should be avoided to enable open scientific enquiry.
In comparing organoids to in vivo neural development in the current state of the field, we do not endorse the use of terms such as fetal-like or the unsubstantiated use of terms describing complex, emergent processes of the nervous system. Regarding the use of terms commonly used to define complex mental, cognitive and behavioural processes, such as sentience, intelligence, learning and pain, we encourage authors to explicitly define cellular activities in the isolated system, rather than using them to infer that organoids are learning or sentient bodies. For example, cells in retinal organoids respond to light and exhibit aspects of phototransduction, but we should not suggest that these organoids have vision or sight.
Final remarks
We agree to implement this nomenclature system in our publications and invite others to align with this framework and provide input as the field evolves. We believe nomenclature should remain flexible to accommodate new developments and information and should be updated as needed. Moving forwards, there may be a need for new names, but for now we recommend avoiding excessive renaming.
Importantly, we believe that this type of work with human cells must be performed in close conversation with ethicists and the broader scientific and non-scientific community. Public support depends on trust that all involved in disseminating findings, including scientists, institutional public relations departments and scientific reporters, are accurately reporting and describing these structures and their use. Descriptive terms that are emotive can cause considerable concern and could damage scientific progress. Overstating or hyping findings is detrimental to the credibility of the field and ultimately undermines efforts to find treatments that are desperately needed for nervous system diseases and disorders.
We are enthusiastic about the potential of human neural multicellular models to inform our understanding of development, evolution and disorders of the nervous system. We are hopeful that this framework will facilitate communication within and across fields, catalyse further discussions and accelerate progress.
References
Sasai, Y. Cytosystems dynamics in self-organization of tissue architecture. Nature 493, 318–326 (2013).
Eiraku, M. et al. Self-organized formation of polarized cortical tissues from ESCs and its active manipulation by extrinsic signals. Cell Stem Cell 3, 519–532 (2008).
Ardhanareeswaran, K., Mariani, J., Coppola, G., Abyzov, A. & Vaccarino, F. M. Human induced pluripotent stem cells for modelling neurodevelopmental disorders. Nat. Rev. Neurol. 13, 265–278 (2017).
Lancaster, M. A. & Knoblich, J. A. Organogenesis in a dish: modeling development and disease using organoid technologies. Science 345, 1247125 (2014).
Pasca, S. P. The rise of three-dimensional human brain cultures. Nature 553, 437–445 (2018).
Kelley, K. W. & Pasca, S. P. Human brain organogenesis: toward a cellular understanding of development and disease. Cell 185, 42–61 (2022).
Quadrato, G., Brown, J. & Arlotta, P. The promises and challenges of human brain organoids as models of neuropsychiatric disease. Nat. Med. 22, 1220–1228 (2016).
Qian, X., Song, H. & Ming, G. L. Brain organoids: advances, applications and challenges. Development 146, dev166074 (2019).
Pasca, S. P. Assembling human brain organoids. Science 363, 126–127 (2019).
Chen, H. I. et al. Transplantation of human brain organoids: revisiting the science and ethics of brain chimeras. Cell Stem Cell 25, 462–472 (2019).
Farahany, N. A. et al. The ethics of experimenting with human brain tissue. Nature 556, 429–432 (2018).
Di Lullo, E. & Kriegstein, A. R. The use of brain organoids to investigate neural development and disease. Nat. Rev. Neurosci. 18, 573–584 (2017).
Brazovskaja, A., Treutlein, B. & Camp, J. G. High-throughput single-cell transcriptomics on organoids. Curr. Opin. Biotechnol. 55, 167–171 (2018).
Author information
Authors and Affiliations
Contributions
All authors discussed the content of the manuscript, and reviewed and edited the entire manuscript. S.P.P. led the writing of the manuscript following extensive discussions and with input from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Simon Hippenmeyer and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Pașca, S.P., Arlotta, P., Bateup, H.S. et al. A nomenclature consensus for nervous system organoids and assembloids. Nature 609, 907–910 (2022). https://doi.org/10.1038/s41586-022-05219-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-05219-6
This article is cited by
-
Modelling the complex nature of the tumor microenvironment: 3D tumor spheroids as an evolving tool
Journal of Biomedical Science (2024)
-
Parechovirus infection in human brain organoids: host innate inflammatory response and not neuro-infectivity correlates to neurologic disease
Nature Communications (2024)
-
Induced pluripotent stem cells (iPSCs): molecular mechanisms of induction and applications
Signal Transduction and Targeted Therapy (2024)
-
Recent advances and current challenges of new approach methodologies in developmental and adult neurotoxicity testing
Archives of Toxicology (2024)
-
Humanized brain organoids-on-chip integrated with sensors for screening neuronal activity and neurotoxicity
Microchimica Acta (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
