
Abstract
Molecular catch bonds are ubiquitous in biology and essential for processes like leucocyte extravasion1 and cellular mechanosensing2. Unlike normal (slip) bonds, catch bonds strengthen under tension. The current paradigm is that this feature provides ‘strength on demand3’, thus enabling cells to increase rigidity under stress1,4,5,6. However, catch bonds are often weaker than slip bonds because they have cryptic binding sites that are usually buried7,8. Here we show that catch bonds render reconstituted cytoskeletal actin networks stronger than slip bonds, even though the individual bonds are weaker. Simulations show that slip bonds remain trapped in stress-free areas, whereas weak binding allows catch bonds to mitigate crack initiation by moving to high-tension areas. This ‘dissociation on demand’ explains how cells combine mechanical strength with the adaptability required for shape change, and is relevant to diseases where catch bonding is compromised7,9, including focal segmental glomerulosclerosis10 caused by the α-actinin-4 mutant studied here. We surmise that catch bonds are the key to create life-like materials.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


Data availability
Source data are provided with this paper.
Code availability
The Python script to simulate the minimal 1D crosslinker model can be found at GitHub via https://github.com/YuvalMulla/Minimal_1D_Crosslinker_Model.
References
Marshall, B. T. et al. Direct observation of catch bonds involving cell-adhesion molecules. Nature 423, 190–193 (2003).
Liu, B., Chen, W., Evavold, B. D. & Zhu, C. Accumulation of dynamic catch bonds between TCR and agonist peptide-MHC triggers T cell signaling. Cell 157, 357–368 (2014).
Mbanga, B. L., Iyer, B. V. S., Yashin, V. V. & Balazs, A. C. Tuning the mechanical properties of polymer-grafted nanoparticle networks through the use of biomimetic catch bonds. Macromolecules 49, 1353–1361 (2016).
Thomas, W. E., Trintchina, E., Forero, M., Vogel, V. & Sokurenko, E. V. Bacterial adhesion to target cells enhanced by shear force. Cell 109, 913–923 (2002).
Huang, D. L., Bax, N. A., Buckley, C. D., Weis, W. I. & Dunn, A. R. Vinculin forms a directionally asymmetric catch bond with F-actin. Science 357, 703–706 (2017).
Petrie, R. J., Koo, H. & Yamada, K. M. Generation of compartmentalized pressure by a nuclear piston governs cell motility in a 3D matrix. Science 345, 1062–1065 (2014).
Yago, T. et al. Platelet glycoprotein Ibα forms catch bonds with human WT vWF but not with type 2B von Willebrand disease vWF. J. Clin. Invest. 118, 3195–3207 (2008).
Luo, T., Mohan, K., Iglesias, P. A. & Robinson, D. N. Molecular mechanisms of cellular mechanosensing. Nat. Mater. 12, 1064–1071 (2013).
Kim, J., Zhang, C.-Z., Zhang, X. & Springer, T. A. A mechanically stabilized receptor–ligand flex-bond important in the vasculature. Nature 466, 992–995 (2010).
Feng, D., DuMontier, C. & Pollak, M. R. Mechanical challenges and cytoskeletal impairments in focal segmental glomerulosclerosis. Am. J. Physiol. Renal Physiol. 314, F921–F925 (2018).
Ehrlicher, A. J. et al. Alpha-actinin binding kinetics modulate cellular dynamics and force generation. Proc. Natl Acad. Sci. USA 112, 6619–6624 (2015).
Ribeiro, E. D. A. et al. The structure and regulation of human muscle α-actinin. Cell 159, 1447–1460 (2014).
Yao, N. Y. et al. Stress-enhanced gelation: a dynamic nonlinearity of elasticity. Phys. Rev. Lett. 110, 018103 (2013).
Mulla, Y., Oliveri, G., Overvelde, J. T. B. & Koenderink, G. H. Crack initiation in viscoelastic materials. Phys. Rev. Lett. 120, 268002 (2018).
Mulla, Y. & Koenderink, G. H. Crosslinker mobility weakens transient polymer networks. Phys. Rev. E 98, 062503 (2018).
Mulla, Y., Wierenga, H., Alkemade, C., ten Wolde, P. R. & Koenderink, G. H. Frustrated binding of biopolymer crosslinkers. Soft Matter 15, 3036–3042 (2019).
Mulla, Y., Mackintosh, F. C. & Koenderink, G. H. Origin of slow stress relaxation in the cytoskeleton. Phys. Rev. Lett. 122, 218102 (2019).
Jung, W., Murrell, M. P. & Kim, T. F-actin cross-linking enhances the stability of force generation in disordered actomyosin networks. Comp. Part. Mech. 2, 317–327 (2015).
Kim, T. Determinants of contractile forces generated in disorganized actomyosin bundles. Biomech. Model. Mechanobiol. 14, 345–355 (2014).
Kim, T., Hwang, W., Lee, H. & Kamm, R. D. Computational analysis of viscoelastic properties of crosslinked actin networks. PLoS Comput. Biol. 5, e1000439 (2009).
Mak, M., Zaman, M. H., Kamm, R. D. & Kim, T. Interplay of active processes modulates tension and drives phase transition in self-renewing, motor-driven cytoskeletal networks. Nat. Commun. 7, 10323 (2016).
Hosseini, K., Sbosny, L., Poser, I. & Fischer-Friedrich, E. Binding dynamics of à-actinin-4 in dependence of actin cortex tension. Biophys. J. 119, P1091–1107 (2020).
Grashoff, C. et al. Measuring mechanical tension across vinculin reveals regulation of focal adhesion dynamics. Nature 466, 263–266 (2010).
Feng, D. et al. Disease-causing mutation in α-actinin-4 promotes podocyte detachment through maladaptation to periodic stretch. Proc. Natl Acad. Sci. USA 115, 1517–1522 (2018).
Rosales, A. M. & Anseth, K. S. The design of reversible hydrogels to capture extracellular matrix dynamics. Nat. Rev. Mater. 1, 15012 (2016).
Rivera, J. et al. Toughening mechanisms of the elytra of the diabolical ironclad beetle. Nature 586, 543–548 (2020).
Wang, C. et al. Visible-light-assisted multimechanism design for one-step engineering tough hydrogels in seconds. Nat. Commun. 11, 4694 (2020).
Zhang, L., Bailey, J. B., Subramanian, R. H., Groisman, A. & Tezcan, F. A. Hyperexpandable, self-healing macromolecular crystals with integrated polymer networks. Nature 557, 86–91 (2018).
Garcia-Manyes, S., Liang, J., Szoszkiewicz, R., Kuo, T.-L. & Fernández, J. M. Force-activated reactivity switch in a bimolecular chemical reaction. Nat. Chem. 1, 236–242 (2009).
Dansuk, K. C. & Keten, S. Self-strengthening biphasic nanoparticle assemblies with intrinsic catch bonds. Nat. Commun. 12, 85 (2021).
Bell, G. Models for the specific adhesion of cells to cells. Science 200, 618–627 (1978).
Pollard, T. D., Blanchoin, L. & Mullins, R. D. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annu. Rev. Biophys. Biomol. Struct. 29, 545–576 (2000).
Skrzeszewska, P. J. et al. Fracture and self-healing in a well-defined self-assembled polymer network. Macromolecules 43, 3542–3548 (2010).
Jung, W., Murrell, M. P. & Kim, T. F-actin cross-linking enhances the stability of force generation in disordered actomyosin networks. Comp. Part. Mech. 2, 317–327 (2015).
Kim, T. Determinants of contractile forces generated in disorganized actomyosin bundles. Biomech. Model. Mechanobiol. (2014).
Kim, T., Hwang, W., Lee, H. & Kamm, R. D. Computational analysis of viscoelastic properties of crosslinked actin networks. PLoS Comput. Biol. 5, e1000439 (2009).
Mak, M., Zaman, M. H., Kamm, R. D. & Kim, T. Interplay of active processes modulates tension and drives phase transition in self-renewing, motor-driven cytoskeletal networks. Nat. Commun. 7, 10323 (2016).
Tang, V. W. & Brieher, W. M. α-Actinin-4/FSGS1 is required for Arp2/3-dependent actin assembly at the adherens junction. J. Cell Biol. 196, 115–130 (2012).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Broedersz, C. P. et al. Cross-link-governed dynamics of biopolymer networks. Phys. Rev. Lett. 105, 238101 (2010).
Yin, J. et al. Genetically encoded short peptide tag for versatile protein labeling by Sfp phosphopantetheinyl transferase. Proc. Natl Acad. Sci. USA 102, 15815–15820 (2005).
Avellaneda, M. J., Koers, E. J., Minde, D. P., Sunderlikova, V. & Tans, S. J. Simultaneous sensing and imaging of individual biomolecular complexes enabled by modular DNA–protein coupling. Commun. Chem. 3, 20 (2020).
Berg-Sørensen, K. & Flyvbjerg, H. Power spectrum analysis for optical tweezers. Rev. Sci. Instrum. 75, 594–612 (2004).
Acknowledgements
We thank M. van Hecke and C. Alkemade for critical reading of the manuscript. We thank P. R. ten Wolde, K. Storm, W. Ellenbroek, C. Broedersz, D. Brueckner and M. Berger for fruitful discussions. We thank W. Brieher and V. Tang from the University of Illinois for the kind gift of purified α-actinin-4 (WT and the K255E point mutant) and their plasmids; M. Kuit-Vinkenoog and J. den Haan for actin and further purification of α-actinin-4; A. Goutou and I. Isturiz-Petitjean for co-sedimentation measurements and V. Sunderlíková for the design, mutagenesis, cloning and purifying of the α-actinin-4 constructs used in the single-molecule experiments. We gratefully acknowledge financial support from the following sources: research program of the Netherlands Organization for Scientific Research (NWO) (S.J.T., A.R. and M.J.A.); ERC Starting Grant (335672-MINICELL) (G.H.K. and Y.M.). ‘BaSyC—Building a Synthetic Cell’ Gravitation grant (024.003.019) of the Netherlands Ministry of Education, Culture and Science (OCW) and the Netherlands Organisation for Scientific Research (G.H.K. and L.B.); and support from the National Institutes of Health (1R01GM126256) (T.K. and W.J.).
Author information
Authors and Affiliations
Contributions
Y.M. and G.H.K. conceived and designed the study. M.J.A. and S.J.T. designed the optical tweezer experiments. M.J.A., A.R. and L.B. performed the optical tweezer experiments. M.J.A. and A.R. analysed the optical tweezer experiments. Y.M. performed and analysed all the other experiments and designed and simulated the 1D model. W.J. and T.K. designed, performed and analysed the actin network simulations. Y.M., M.J.A., T.K., S.J.T. and G.H.K. wrote the manuscript with input from W.J., A.R. and L.B. All the authors approved the final version.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Materials thanks Patricia Bassereau, Jean-Francois Joanny and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 High-speed co-sedimentation measurements of the affinity of α-actinin-4 (WT) and K255E crosslinkers for actin filaments.
a, b, supernatant resulting from a high-speed centrifugation of a mixture of actin filaments and crosslinkers was run on an SDS-page gel. The bands on the bottom show the α-actinin-4 (WT) or K255E (resp. a and b, molecular weight ∼ 100 kDa in both cases), while the bands on the top show actin (42 kDa). Each labelled column contained a different actin concentration as indicated. Some lanes were kept empty as spacers. The crosslinker concentration was fixed at 0.1 μM. A single measurement was performed per condition. c, The fraction of bound crosslinkers, as determined from the co-sedimentation assay, as a function of the actin concentration was fit to the equation: \(\varphi _{bound} = \frac{{c_{actin}}}{{K_a}}\), where Ka is the affinity of the crosslinker. d, Consistent with the high affinity of both crosslinkers, SDS-page gels of supernatant resulting from a high-speed centrifugation of a crosslinked actin network at the concentration used in all our experiments (48 μM actin together with 0.48 μM crosslinker) do not show any measurable fraction of unbound crosslinkers. A single measurement was performed per condition.
Extended Data Fig. 2 Fluorescence recovery after photobleaching measurements reveal that α-actinin-4 crosslinkers are more dynamic than the K255E mutant.
Example fluorescence recovery curves of α-actinin-4 (a) and K255E (b) in the presence of 48 μM actin show full recovery of both proteins after photobleaching at time t=0, but with different timescales. The solid lines represent exponential fits to the data (see Methods). c, Average recovery time for α-actinin-4 (red) and for K255E (blue), with the standard error on basis of 5 measurements of different locations within the same sample per condition. Measurements were performed at 25 °C.
Extended Data Fig. 3 Generation and classification of α-actinin-4/actin tethers.
a, DNA was coupled to α-actinin-4 (WT or K255E) using an SFP synthase-mediated reaction. Because α-actinin-4 is a homodimer, the yBBr tag used for coupling is present in both monomers. To favour DNA attachment to only one monomer, we performed coupling reactions with several DNA titrations, and the coupling yields were quantified using SDS-PAGE gel electrophoresis. The DNA:α-actinin-4 molar ratios are indicated above each lane. At a molar ratio of 1:1, most of the α-actinin-4 is uncoupled, that is most dimers will be either not coupled or have only one monomer coupled to DNA. A single measurement was performed per condition. b, Concurrent confocal fluorescence images of a trapped bead coated with α-actinin-4 (left) and a trapped bead coated with actin filaments (right). The bead’s autofluorescence is depicted in green, and the fluorescent emission of Alexa Fluor 647-tagged actin is depicted in orange. In total, 36 images have been taken of 4 independent samples. This image shows representative examples. c, Force-extension curves showing the overstretching regime of a single dsDNA tether (black), and a case where the two beads are linked by multiple tethers, which yields a shorter contour length and higher forces without unzipping (red). Variability in bead radii and actin layer thickness results in force-extension curves that can be shifted along the Extension axis, from the theoretical 850 nm by ±30 nm. Grey area:’single-tether region’. Tethers with a force-extension curve within this area that broke in a single step were regarded as single tethers and hence included in measuring the force-dependent lifetime.
Extended Data Fig. 4 Nonlinear and temperature-dependent rheology of actin networks crosslinked by α-actinin-4 or K255E.
Rheological measurements of wild type (red) and K255E mutant (light blue) α-actinin-4 crosslinking actin networks at 10 °C and of K255E mutant crosslinked actin networks at 25 °C (dark blue). a-c, The storage (triangles) and loss moduli (circles) were measured using small amplitude oscillatory shear. The moduli are shown as a function of frequency (a) and as a function of the frequency normalized by the frequency at which the loss modulus peaks (b). Both curves are measured at 10 °C. Data are presented as mean values + /- the standard error indicated by bars and shaded regions on basis of 4 independent samples per condition. The collapses in b and c show that the crosslinker unbinding kinetics, but not the network structure, is significantly different for the different conditions (see Main Text). d, The stress relaxation frequency was extracted from Extended Data Fig. 4a, c using Methods equations 4–7. Data are presented as mean values + /- the standard error on basis of 4 independent samples per condition. e, Representative example curves of the differential storage modulus at 0.5 Hz (top) and of the strain rate (bottom) are plotted against the applied shear stress. We define the rupture strain as the data point where K’ peaks. f-g, We apply a semiflexible polymer network model to fit the frequency-dependent differential elastic modulus as a function of prestress (see Methods). h, Thus, we extract the crosslinker bound lifetime as a function of stress for both α-actinin-4 (red) and the K255E mutant (blue) at 25 °C. The shaded areas represent the error on basis of the fits. The bound lifetime of the mutant is significantly longer at low stress, but the lifetimes of both types become similar at high stress as the bound lifetime of the wild type increases. The abrupt decay of bound lifetime in the K255E-crosslinked network when the stress reaches 5 Pa is due to network fracturing.
Extended Data Fig. 5 Fracturing in the minimal 1D crosslinker model.
a, Time trace of the bound number of catch bonds (red) and slip bonds (blue) in a network undergoing a linearly increasing stress (see Supplementary Table 1 for parameters). As the catch bonds have faster dynamics than the slip bonds, a larger spread in the bound fraction is observed. After a long time of steady state fluctuations, the networks suddenly fracture as the number of linkers rapidly goes to 0. b, c, Kymographs showing at which positions there are bonds (red for catch bonds, blue for slip bonds) or no bonds (white). At steady state, linkers continuously bind and unbind (−1000 to approximately −300 steps). Cracks can spontaneously initiate and propagate through the network (the last ~300 steps of the simulation) for both catch and slip bonds in a similar manner. d, The fraction of 1D-networks that rupture when a gap of varying ablation length lablate is introduced for both catch (red) and slip bonds (blue). Inset: schematic of the ablation simulation.
Extended Data Fig. 6 Simulations show that catch bonds only provide a mechanical advantage over slip bonds when they are mobile and present in sufficiently large numbers.
The system size dependence of the rupture stress (a) and bond turnover at the point of rupture (b) reveals that catch bonds (red) are only stronger than slip bonds (blue) for networks larger than ~10 bonds, emphasizing that the increased network strength by catch bonding is an emergent property (Supplementary Note 1). Each data point is the average of 10 repeats and the standard errors are smaller than the symbol size. c, Catch bond-induced network strengthening is not observed when crosslinkers are immobile and rebind in the same location from which they unbound. The bond turnover as a function of stress reveals catch bonds (red) cause more dynamic materials (right), but do not enhance strength (top) compared to strong slip bonds (light blue) and are less dynamic than networks consisting of weak slip bonds (dark blue). Data are presented as mean values + /- the standard error on basis of 100 independent simulation runs per condition. d, We also considered a three-state model where linkers are doubly bound, singly bound or unbound (Supplementary Note 1). Similar to the two-state model, the bond turnover as a function of stress reveals that networks of catch bonds (red) are stronger and more deformable than networks of strong slip bonds (light blue) or weak slip bonds (dark blue). Data are presented as mean values + /- the standard error on basis of 5 independent simulation runs per condition.
Extended Data Fig. 7 Catch bonding is only effective when the bond lifetime is high.
a, Simulations of the rupture stress as a function of the bond lifetime \(k_{on}/k_{off,0}^{slip}\), keeping \(k_{off,0}^{slip}/k_{off,0}^{catch}\) fixed (see Methods and Supplementary Table 1), shows that catch bonds (red) are only stronger than slip bonds (blue) when the binding rate is high. b, Consistent with the simulations, enhancing the bond lifetime in experiments by decreasing the temperature from 25 °C (light) to 10 °C (dark) increases the rupture stress more steeply for wild type α-actinin-4 (red) than for K255E (blue). The error bars represent the standard error (N=4 independent samples for each condition).
Extended Data Fig. 8 Actin network simulations.
a, Stress-strain curve of the simulated catch (red) and slip bond (blue) actin networks. The black circles indicate the yielding points of both networks (see Methods). b, Distribution of tension on actin filaments in networks for simulated catch bond (red) and slip bond (blue) with 10 Pa stress. c, The polymer network simulation predicts that the average number of active crosslinkers per filament at 10 Pa is lower for catch bonds than for slip bonds, in line with the 1D simulations (panel a) and the catch bond’s lower bond lifetime. d, The average number of crosslinkers as a function of the filament tension shows that slip bonds are mainly enriched on low-tension filaments. The tension on the x-axis is binned such that each bin contains 10% of the filaments. e-f, Similar plots as Fig. 3d but then for the catch (d, red) and slip bond (e, blue) simulations separately: the active crosslinkers are binned according to tension acting on pairs of filaments (tension on filament 1 on x-axis, tension on filament 2 on y-axis) connected by the crosslinkers. 10×10 bins are used, and the distribution was smoothed using bicubic interpolation. The tension spacing along the x- and y-axis is non-uniform, such that each bin includes 10% of the filaments. These plots show that both catch and slip bonds preferentially connect tense filaments, likely because of geometrical reasons and/or because filament tension resulted from having more crosslinkers bound. However, catch bonds localize more strongly to tense filaments than slip bonds (Fig. 3d) because they bind less to the rest of the network due to their higher off-rate in the absence of force and therefore redistribute to the tense filaments (Fig. 3e).
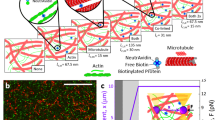
Extended Data Fig. 9 Confocal fluorescence images of crosslinked actin networks.
10% of the actin monomers were labelled with Alexafluor-647. At a 1:100 crosslinker:actin molar ratio, the actin networks studied in this work are isotropic and spatially uniform, for both wild type (a) and K255E α-actinin-4 (b). We do not observe any discernable structure because the mesh size is ~200 nm, which is on the order of the diffraction limit, indicating that filaments are isotropically crosslinked rather than bundled. c, For comparison, actin bundle clusters were observed at a 1:25 α-actinin-4:actin molar ratio. The colour coding was inverted for all images to improve the visual contrast between bundles and background. Scale bars are 20 μm. 10 images were taken of different locations within the same sample per condition and all images had similar results per condition.
Extended Data Fig. 10 Fracturing occurs within the actin network, not at the rheometer-network interface.
The rheology of wild type α-actinin-crosslinked actin networks was compared in the presence (dark green) or absence (light green) of a Polylysine-coated surface on both the bottom and top plate of the rheometer (see Methods). a) a frequency sweep at zero prestress shows that the linear rheology is unaffected by changing the rheometer-network interface. The storage (triangles) and loss moduli (circles) were measured as a function of frequency using small amplitude oscillatory shear. b) the network rupture strain and c) rupture stress (bottom) are not significantly affected by the addition of Polylysine at the rheometer-network interface. The error bars represent the standard error (N=4 independent samples for each condition).
Supplementary information
Supplementary Information
Supplementary Notes 1–3 and Tables 1 and 2.
Supplementary Video 1
Visualization of the top 10% stressed filaments in actin network simulations of catch (top) and slip (bottom) bond networks with actin filaments shown in green and the network borders shown in white. The network is strained over time by moving the top boundary. The x and y axes show the distance in micrometres.
Supplementary Video 2
This video shows the same analysis as that in Fig. 4c, with each frame showing the distribution at a different stress level (top-right corner). Colour scale, percentage relative enrichment of catch bonds over slip bonds bound between a pair of actin filaments for varying tensions on filament 1 (vertical axis) and on filament 2 (horizontal axis). Specifically, the colour scale is defined as (\(\left( {\frac{{n_{\mathrm{catch}}}}{{n_{\mathrm{slip}}}} - 1} \right) \times 100\%\)) when ncatch > nslip (shown in red) and (\(- \left( {\frac{{n_{\mathrm{slip}}}}{{n_{\mathrm{catch}}}} - 1} \right) \times 100\%\)) when nslip > ncatch (shown in blue), where ncatch and nslip are the number of respective catch and slip bond linkers connecting two filaments. The tension spacing is the same for the x and y axes and is updated at every timestep such that each bin includes 10% of the filaments, where the top-right corner represents connections between the tensest filaments.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 1
Unprocessed image.
Source Data Extended Data Fig. 1
Unprocessed image.
Source Data Extended Data Fig. 1
Unprocessed image.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 3a
Unprocessed image.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 10
Statistical source data.
Rights and permissions
About this article

Cite this article
Mulla, Y., Avellaneda, M.J., Roland, A. et al. Weak catch bonds make strong networks. Nat. Mater. 21, 1019–1023 (2022). https://doi.org/10.1038/s41563-022-01288-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41563-022-01288-0
This article is cited by
-
Adherens junctions as molecular regulators of emergent tissue mechanics
Nature Reviews Molecular Cell Biology (2024)
