
Abstract
The development of induced-pluripotent stem cell (iPSC)-derived cell types offers promise for basic science, drug testing, disease modeling, personalized medicine, and translatable cell therapies across many tissue types. However, in practice many iPSC-derived cells have presented as immature in physiological function, and despite efforts to recapitulate adult maturity, most have yet to meet the necessary benchmarks for the intended tissues. Here, we summarize the available state of knowledge surrounding the physiological mechanisms underlying cell maturation in several key tissues. Common signaling consolidators, as well as potential synergies between critical signaling pathways are explored. Finally, current practices in physiologically relevant tissue engineering and experimental design are critically examined, with the goal of integrating greater decision paradigms and frameworks towards achieving efficient maturation strategies, which in turn may produce higher-valued iPSC-derived tissues.
Similar content being viewed by others
Introduction
The scientific community is investing heavily in tissue engineering to produce highly functional models of most tissues in the human body, which both elucidate the processes underlying healthy development and function as well as the mechanisms underlying pathology. As a result, tissue engineering has achieved considerable advancements over the past decade and holds significant potential for application in cell therapy and the generation of artificial organs for implantation. Driven by a long-term vision for physiological recapitulation, a shorter-term goal for the application of engineered tissues lies in preclinical drug testing for both tissue-specific efficacy and toxicity. Another significant driver of pharmaceutical screening is the severe attrition rate of drugs in the current clinical trial paradigm. This challenge carries a significant cost burden, and is inefficient with respect to development time, animal lives, and trial participant outcomes that could be better-invested with drug candidates that were better-filtered through highly reliable preclinical models (e.g., functional human in vitro tissues as opposed to immortalized human cell lines and in vivo animal models, both of which carry certain inherent inaccuracy1). Moreover, post-release drug recalls demonstrate that even successfully marketed drugs may carry significant risk to the population that cannot be detected even in the medium-throughput environment of late-stage clinical trials2,3. By refocusing on failing drug candidates for reasons of toxicity or inefficacy earlier in the process, and passing drug candidates that may be viable in humans but not in animal models, significant patient benefit, as well as cost and time savings, could be realized in the development of dependable drugs. To reach this point, continued progress in engineering tissues that closely replicate function in vitro is required. Widespread investment in the design of organ-on-a-chip models, that allow for standardized high-throughput microphysiological system experimentation, has propelled the field toward commercial and clinical relevancy. However, the functional maturity of most engineered tissues is limited by the extent of that maturation, thereby presenting a new challenge to the field of systems modeling that does not seem to have a simple and generalizable solution.
In tissue engineering to date, special attention has been paid to tissue structure and system throughput, with both reliability and robustness recognized as essential for uptake and widespread use by both industry users and scientists4. Furthermore, the field has recognized the importance of comprehensive and quantitative physiological metrics by which to grade models; these are usually emergent (i.e., quantitative in themselves, but difficult to subdivide into measurable component factors that fully account for the presentation). Examples of such emergence in the form of functional physiological metrics, readouts, or endpoints depending on tissue type may include contractile kinetics and dynamics, barrier function or absorption, connectivity, electrophysiology or Ca2+ handling, or metabolic kinetics (biochemical output or detoxifying flux)4,5. These functional metrics have been ubiquitously correlated with highly specific and differentiated tissue architecture (e.g., vascularized tissues, muscle with physiologically relevant resistance, the blood-brain barrier, intestinal crypts, liver portals, etc.) as well as allowing for the recreation of relevant functional interfaces, physiological fluid flow, and selective permeability. These attempts all aim to recapitulate the tissue niche (i.e., the biochemical and mechanical local environment) as closely as possible. Furthermore, paracrine and autocrine signaling, as well as direct contact and physical signaling phenomena, may represent mechanisms underlying much of this function. Many of the traditional and high-yield aspects of tissue engineering (co-culture, 3D tissue organization) to date have been prioritized to recreate the niche. Although the physical complexity and (bio)fabrication schemes required to create some of these models has been a challenge, there have been significant advancements. As a result, recapitulating the (bio)chemical niche of those tissues is now beginning to be recognized as an active field with high-yield opportunity for iteration.
“Cell maturation” itself is a poorly-defined process, and maturity a poorly defined state. For nearly all cells, maturity combines characteristic and interdependent states of morphology, metabolism, and cell-cycle arrest (quiescence, terminal differentiation, or senescence; depending on cell/tissue6) that cooperatively lend hallmark functionality to a cell for it to contribute to the physiological role of its tissue (Fig. 1). The environment conducive to cell maturation, in general, results from commonly recognized modulators of cell response and differentiation including mechanical signaling, hormone availability, and paracrine and autocrine excretions. In addition, maturing cells also benefit from metabolic substrate availability, cofactors, and other signaling-active small molecules, ionic complement, and osmotic profile, as discussed throughout the text below. As such, tissue or cell maturity in this context may best be defined as a state of signaling and metabolic homeostasis within a tissue, driven by both exogenous and endogenous stimuli, that is enabled by sufficient energetic flux to maintain the range of peak functional outputs characteristic of that tissue when enabling healthy activity, leading to functional activity.

As metabolically provided energy is diverted from proliferative activity to physiological function, cell complexity, and functional parameters are improved. This increasing specialization necessitates an investment of energy to manage cell size, structure, and specialized structures or organelles. The energetic outlay and continued flux for this expenditure is generally maintained by high-yield and efficient oxidative phosphorylation from lipids, short-chain fatty acids, carbohydrates, amino acids, lactate, and/or ketones, depending on the specific tissue and stage of maturation in question. Metabolic supply is provided by increased perfusion and spatial zonation, at which point the tissue can then exert its hallmark function. Image created using BioRender.com.
In this review, we describe the convergence of key cell signaling pathways that regulate cell growth, extracellular matrix (ECM) deposition, terminal differentiation, and maturation of functional tissues. Given the wealth of existing in vivo knowledge associated with metabolic regulation, we refer to many excellent reviews and primary articles that offer additional detail relating to important effectors of the metabolic state. Several key recurring signaling pathways are further contextualized with insight into their potential mechanisms in evoking functional maturation. Finally, we highlight and discuss design strategies that may be well-suited to advance the function of engineered tissues. The discussion is also examined in the context of experimental design strategies developed in other engineering disciplines, in order to screen complex culture conditions using quantitative functional parameters as objective metrics. As a whole, this review defines the insight needed to yield efficient and biologically-inspired tissue engineering that could evoke maximal functional maturation without undue experimental failure or redundancy.
Potential signaling convergences in specialized engineered tissue models: case studies
Myocardium
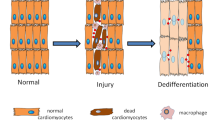
The high burden of both cardiac toxicity and lack of cardiac efficacy in pharmaceutical clinical trials or clinical use, as well as the low regenerative potential of the adult myocardium post-trauma, have led to a significant investment, in both innovation and funding, toward enabling cardiac tissue engineering for screening7,8,9 and therapeutic goals10,11,12. Adult myocardium is a highly specialized tissue in structure, function, and metabolic profile, growing in complexity and metabolic profile through development and postnatal maturation13,14,15. Although many factors contributing to maturation have been identified (Table 1), their precise mechanisms and the full complement of maturation factors necessary to produce adult myocardium remain elusive. Myocardial functional specialization reflects significant changes to cell homeostasis, with extensive mitochondrial biogenesis and turnover16, fueled both by extreme internal energetic demands and exogenous (e.g., endocrine and paracrine) influences on cardiomyocyte (CM) structure and metabolic organization. Fully-matured myocardium retains extensive, millisecond-level regulation on mitochondrial turnover and connectivity to maintain full and efficient oxidative coverage of CM function17. In contrast, the diseased state, with reduced function in at least some aspect by definition, tends to necessitate a lesser degree of structural and metabolic organization and so can revert to a state resembling lower-maturity CMs in certain aspects, with corresponding decreases in function18. Given the significant burden of cardiovascular pathology and the shortcomings of 2D systems to recapitulate adult functional benchmarks18,19,20, 3D culture systems continue to evolve e.g.21,22,23,24, and often make use of concurrent electrical stimulation techniques which have been found to improve CM functionality in vitro25,26,27. As induced-pluripotent stem cell (iPSC)- or embryonic stem cell (ESC)-derived CMs can be highly matured when transplanted into adult hearts, logic would dictate that a high degree of in vitro maturation is possible (i.e., there is not a fundamental limit on the level of functional maturation attainable by PSC-CMs), although the conditions necessary to achieve it remain elusive28.
The myocardium is highly oxygenated, with each CM nominally directly accessible by a capillary to enable the large oxidative flux required to fuel the continuous ion transport and contractility of the myocardium. Indeed, the perinatal switch to oxidative metabolism13,14,15 seems to be mediated in part by the permanent increase in myocardial oxygenation that occurs in the switch from placental to pulmonary respiration; this rapid metabolic refocusing appears coordinated by Hand129, as its overexpression diminishes fatty acid (FAO) oxidation and maintains glycolytic dependency. Hand1 is induced by HIF1α, of which perinatal downregulation during the transition to an oxygen-rich environment is well-known to be critical to the oxidative switch in the heart30,31,32. The DNA damage response to oxidative damage is a key mediator of cell-cycle arrest, and the rate of this arrest is proportional to the extent of oxidative damage33. Standard cell cultures, depending on cell type and density, can be chronically hypoxic or hyperoxic without obvious signs such as cell die-off34. Both hypoxia and hyperoxia can induce oxidative stress, as discussed below. Together, this evidence suggests that oxygen control through medium coverage34, as well as medium antioxidant supplementation, needs to be carefully titrated for the desired results, especially in advanced culture systems. Finally, even without perfusion, the inclusion of endothelial cells in myocardium can improve physiological metrics, both in vitro35 and in vivo36.
A key feature of perinatal CM maturation is the downregulation of proliferative processes, and the concomitant initiation of cellular hypertrophy that translates to whole-heart growth. Cardiac hypertrophy has been linked to AMPK-Sirt1 signaling, which is linked to the NAD+/NADH ratio as discussed in section 3.1 below37, and maturation-associated processes have been linked to PGC-1α/PPAR activity38,39. NAD+ availability also tends to decrease with age40, which therefore suggests negative modulation of cardiac differentiation and general mature phenotype as disease onset pushes CM phenotype to a neonatal-adjacent physiological state18,41, although actual transcriptional and proteomic states may vary. Regulation of AMPK/Akt signaling has been detected in heart tissues, including through miRNA-mediated pathways42. Some of this modulation may include AMPKγ isoform switching, demonstrated to mediate changes in intrinsic cardiac pacing43 and illustrating the importance of energetic signaling on phenotype. Important clues to homeostasis in pathology might be evident by more closely examining the metabolic phenotype. Similarly, the expression of AMPKα2 vs. α1 subunit expression rescues the heart failure phenotype, ostensibly through promoting selective mitophagy44. This activity may prevent energy limitation, excess ROS production, and the release of cytochrome C, the latter of which would trigger apoptosis. The authors attribute this isoform-dependent activity to the ubiquitination kinetics of the α2 vs. α1 subunits. In downstream signaling, ERRγ has been implicated in activating cardiac transcription patterns controlled by GATA, MEF2, and TEAD, while downregulating NR2C2 which itself transcriptionally represses the nuclear PPARα, retinoid receptors, and thyroid receptors45 that were discussed above, indicating synergies in maturation signaling. ERRs also directly activate many contractile, calcium handling, and ion channel genes indicative of maturation state45,46.
Several actionable biochemical effectors of CM function have been studied, and are being introduced into culture practices20,28. The peptide hormone neuregulin may revert to maturity and seems to increase proliferation, but also increases cardiac performance, ostensibly through ErbB signaling47. Additionally, the targeted inhibition of ErbB4 stops the cell cycle and induces maturation48. Neuregulin, a suggested myocardial regenerative agent, has also been implicated in enhancing CM maturity when used in conjunction with insulin-like growth factor-1 (IGF-1)49. Triiodothyronine (T3) is also known to be vital for healthy heart development50. Briefly, T3 increases the expression of natriuretic peptide receptors, adrenergic receptors, cytoskeletal/contractile proteins, and the complement of ion channels responsible for the development of specialized electrophysiological transients of the adult heart. T3 mediates an anabolic switch in CMs from proliferative to hypertrophic processes, likely through modulation of MAPK and Akt/mTOR pathways51,52, and has been widely used for physiological maturation of PSC-CMs in vitro38,53,54,55,56,57. Similarly, glucocorticoids have been used for CM maturation in vitro38,55,57, potentially through the activation of PGC-1α58. Retinoids are deeply implicated in regional development of the embryonic heart59, but can also promote oxidative metabolism and electrophysiological/Ca2+ handling maturation through unelucidated mechanisms60. Given the importance of retinoids in the heart field and CM subtype specification59,61, the continued role of retinoids in evoking higher function would not be unexpected.
Mechanical stimuli and progressive electrical pacing21,24,25,27,62, and Ca2+ homeostasis have been demonstrated to be critical in proper CM maturation. Sarcomerogenesis in particular is reliant on mechanical signaling,63 and sarcomeric complex-mediated mechanical signal transduction may exert significant effects on cellular disposition64,65. Focal adhesion/Z-disc signaling is likely another important effector of the mature CM phenotype, and often implicates Rho signaling66,67,68. In addition to its role in mediating contraction, Ca2+ cycling through a contractile cell may exert complex analog signaling, as described above, through PKC activation (likely tempered by lipid homeostasis and second messengers, depending on the isoforms at play). In parallel, Ca2+-induced calcineurin activity mediates cell-cycle arrest69 and in concert with the HIF activity described above may be a contributor to the switch between hyperplasia and hypertrophy in the young heart. Finally, calcineurin/LIM and cysteine-rich domains 1 (LMCD1) interactions impact cardiac development and growth70, but must be carefully tuned to avoid pathological hypertrophy71.
The heart displays a highly specialized metabolic phenotype, transitioning from a primarily-glycolytic to a high-flux and primarily-oxidative tissue over a short perinatal period15,41,72, as previously discussed. Healthy adult CMs demonstrate a clear preference for lipids and lactate as primary carbon sources, with flexibility toward glucose, ketone bodies, and amino acids73. However, the degree of flexibility and the potential interactions between substrates’ regulation of each others’ usage remains an open question, and are likely sensitive to the model (in vivo vs. in vitro, tissue vs. cell vs. isolated mitochondria) and the timescale41,73,74,75,76 studied. Importantly, this metabolic flexibility and overall flux seems dependent on continued Ca2+ homeostasis in the CM; aberrant Ca2+ flux via ryanodine receptor abrogation results in pathological remodeling in the cell, metabolic quiescence, and autophagy77. In general, there are both long- and short-term effects from signaling, remodeling, and allostery, based on substrate availability and preference with regard to cardiac function and respiration74. The functional effects of specific carbon sources, particularly toward maturation, are active areas of research; in general lipids seem to promote functional maturation23,38,78,79, but can seemingly induce lipotoxicity if not titrated appropriately53. From a mechanistic perspective, the removal of dietary FAs in a mouse model extended CM proliferation80, while PSC-CMs cultured in high vs. no glucose increased nucleotide production, maintained proliferation, and demonstrated lower maturation81. This evidence lends support to a ROS-induced switch from proliferation to maturation, and suggests several interesting routes of investigation to induce greater CM maturity in vitro, as well as potential targets or mechanisms to arrest or reverse pathological progression in vivo.
Neural tissue
Although individual neural function is well-understood, the emergent properties of even small neural assemblies are very difficult to assess in vivo, with many poorly understood or even unrecognized inputs being essential to define a system. The drive to better control conditions for both applied pharmacological toxicity/efficacy screening and for basic science research into both central and peripheral nervous function and pathology have led to interest in neural tissue engineering. However, by definition, the insight garnered from a neural model is a direct function of its complexity, necessitating advanced culture techniques, and so the development of a minimum viable neural organoid is ongoing. The blood-brain barrier (BBB), formed by endothelium and glia, provides a controlled ionic and molecular environment to the neurons of the brain, ensuring proper function, despite potentially large (bio)chemical changes in the rest of the body. This relies on active transport through two layers, comprised of different types of cells (both endothelial and glial), which must transport nutrition and hormones to any neuron. This highly differentiated structure has not been ignored, and key functional metrics (e.g., excitatory and inhibitory activity) have been advanced using simple co-culture in either contact or Transwell models82. 3D models further improve function and give rise by definition to the potential for brain-like structure, including spontaneous tissue layer stratification within an organoid, and potentially cortical folding (gyrification)83, which is a morphological metric enabling greater organizational complexity. As for additional cellular-level structure, Qian et al.83 also review the available literature on endothelial and glial co-culture in 3D organoids. Efforts to include endothelial cells in brain organoids have resulted in gains to physiological function, spontaneous formation of in vivo-resembling developmental architecture, and the ability to engraft in vivo84,85. The current state is such that it is difficult to conceptualize an effective model of higher tissue functionality that does not faithfully recapitulate 3D structure and the BBB/vascular organization in doing so. Efforts to physiologically mature neural tissue are summarized in Table 2.
The brain is a significant energetic sink, consuming up to 25% of total metabolized glucose in the human body86. Both neurons and glial cells have GLUT3 transporters87 that allow for direct glucose transport from the vascular endothelium. However, despite this level of energetic activity, metabolic flow and access to the brain are highly controlled by the BBB to maintain homeostasis. Neurons have high non-obligate glycolytic capacity, commensurate with their level of maturity, but their oxidative rate also appears to increase proportionally to functionality88, suggesting that mature function is reliant on oxidative phosphorylation. In contrast, glia are primarily-glycolytic87,89. Glucose can be directly imported to neurons, but due to their high relative glycolytic rate, glia surrounding blood vessels provide the generally-preferred lactate substrate via their own glycolysis89,90.
Astrocyte morphology is polar, with distinct subcellular compartmentalization to allow for homeostatic support of neurons. Astrocytic perisynaptic processes uptake synaptically-released glutamate using passive Na+/K+ release along their gradients; the gradient is maintained with concurrent NKA activity, and is energetically supported locally by sequestered mitochondria at these perisynaptic regions89. Given the low density of glial mitochondria, whole-cell activity is primarily supported by the glycolytic activity of the astrocyte which is typically elevated relative to that of the more oxidative neuron. The astrocyte then secretes waste lactate via monocarboxylate transporters, which can be used as a preferential oxidative substrate by nearby neurons (i.e., the lactate shuttle). Astrocytes also exhibit Ca2+ transients, ostensibly due to synaptic contact; the mechanisms underlying this physiology are still in question91, but these regular transients could be tied to increased metabolic demand and be, in part, responsible for regulation of the highly specific astrocyte metabolic response to a synaptic event. In a physiologically-similar process, neurons seem to exert spatial control of their mitochondria in response to intracellular glucose gradients, so as to mediate metabolic control of transport92. Direct signaling by metabolic substrates acting via ligands is relatively unexamined in neural tissues. However, short-chain FAs have been shown to activate GPR41/FFAR3, and by extension PLCβ/MAPK signaling, leading to the induction of sympathetic functional processes; in contrast, ketone bodies inhibit this signaling93, similar to in intestinal epithelium (below).
Neural development, both on a cell and tissue level, is known to be heavily mediated by the activity of the Akt pathway94,95 and its interactions with physical (i.e., Yap pathway) signals96. Akt-related signaling is also known to maintain neuronal function97, however, given the metabolic demands outlined above, sustained AMPK activation is also required to maintain a “healthy” neural phenotype98, potentially as a downstream effect of constitutive HIF2α activation99. It is not yet clear whether this AMPK activation is a relatively flat and continuous phenomenon, or an oscillatory pattern on an unknown timescale (see below for the role of oscillatory signals in tissue and system homeostasis). This could depend on intracellular architecture in the context of diffusion-based limitation and the need for intracellular metabolic shuttling. Nevertheless, this underscores the dipolar relationship between metabolism and the gain or maintenance of function. However, AMPK hyperactivation has been implicated in synaptic destruction via its induction of autophagic processes in cell culture and hippocampus in vivo100. Together, this evidence lends itself to the hypothesis that a homeostasis between AMPK- and Akt-consolidated signals provides a set of both antagonistic and synergistic controls toward coordinated maturation in neural tissue.
Hepatic tissue
The liver, as the metabolic center of the body, carries out many functional roles both for its own maintenance, as well as whole-body homeostasis. Although well-known as the primary detoxification tissue in the body due to an array of cytochrome P450 isoforms that oxidize a wide number of small molecules, hepatic tissue also mediates glucose and lipid processing, including significant glycogen storage, FA synthesis and oxidation, and cholesterol and lipoprotein synthesis. Hepatic culture models in common use include neonatal hepatocytes, PSC-derived hepatocyte-like cells, and hepatoma cell lines (hepatocyte-like and cholangiocyte-like cells). Putative markers of hepatic tissue maturity include the formation of functional bile canaliculi featuring tight junctions, cell polarization, and the expression of drug transporter at the apical and sinusoidal poles101,102,103,104; advancements in evoking hepatic tissue maturity are summarized in Table 3. To date, most in vitro modeling studies have focused on the hepatocyte and increasing its functional maturation; the advancement of Kupffer cells, stellate cells, and hepatic endothelium may prove to be essential toward fully recapitulating native tissue in an engineered setting105.
The metabolic role of hepatocytes includes the esterification and circulation of absorbed lipids from diet or hepatic synthesis, as well as the partial oxidation (ketogenesis) of FAs liberated from peripheral adipocytes during low-energy periods; the latter process is substrate-regulated by negative feedback involving malonyl-CoA106 and is therefore limited by equilibrium except in times of extreme energetic demand (i.e., requiring whole-body ketosis). Hepatocytes cannot effectively metabolize these ketones, and so they are reserved for other tissues. Although the liver has extensive glycolytic capacity, these products are largely used for biosynthetic reactions via the pentose phosphate pathway. Instead, the liver largely subsists on oxidation of amino acids via the TCA (e.g., α-ketoacids); indeed, the provision of extra amino acids in culture medium has been shown to significantly enhance the functional output of hepatocytes in vitro107. The liver also produces IGF-1 in response to growth hormone; this activity is linked to hepatic maturation in vivo108. IGF-1 is a near-ubiquitous growth and maturation signaling agent with effects seen in many tissues as covered elsewhere in this review.
Hepatic functional maturation has been linked to several ubiquitous pathways, including PKA102,109,110, AMPK via LKB1110,111, and the MAPK cascade111. Physical signaling has been less-investigated in hepatic differentiation and maturation when compared to other tissues, although 3D or spatial influences have been implicated in tissue structural development109. Formation and maintenance of the bile canalicular network111,112, and its barrier function103, are partially regulated by bile-activated LKB1/AMPK activity. Akt has been implicated in the early phases of hepatic regeneration, in part through its role in mediating the signaling resulting from several hepatic growth factors113; Akt has also been tied to either developing or maintaining the metabolic functions of the liver, especially those intersecting with insulin activity and sensitivity and notably through its regulation of FoxO-controlled gene expression114. Many extracellular signals have been implicated in hepatic maturation. Hepatocyte growth factor (HGF) is a well-known enhancer of hepatocyte functional maturation115,116, whose processes are largely controlled through c-Met activity117. Oncostatin M acts to enhance tissue function, most likely through STAT3115 and in cooperation with glucocorticoid signaling, although the variance between individual glucocorticoids and their respective affinities to specific receptor isoforms makes it difficult to elucidate the underlying mechanisms118. Separately, T3 hormone increases gluconeogenesis and IGF excretion50. Although certain pathways are now well-implicated in liver maturation, it is not clear which transcription factors or direct signaling events are most closely implicated in liver tissue maturation. There is evidence that early development includes a process of gene bookmarking that can enable or enhance functional maturation119; atypical culture or organoid formation could therefore limit the theoretical functional ceiling of the “matured” tissue.
Many studies have suggested the importance of the local metabolic environment in liver development. The liver displays feed-forward effects of primary bile acid production, as taurocholate is implicated in maturation110,111. This sensitivity itself carries implications of intestinal microbiome contributions, as in intestinal cultures (discussed below). Lithocholic acid and vitamin K2 (menaquinone) compounds, which are microbially derived in vivo, induce gap junction formation120 and regulate CYP activity121. These effects in turn would seem to implicate the establishment of polarity (tying into the physical signaling discussed), as well as cell-cell communication. As suggested previously, endothelial co-culture increases maturity, including the spontaneous formation of rudimentary vasculature101. 3D culture improves gene expression and morphology122. 3D heterotypic cell-cell contact and cell-ECM interactions also induce polarization and the development of functional architecture123; cell spatial relations and ECM gradients also allow for metabolic zonation124. Direct contact and paracrine efflux from nonparenchymal cells (endothelial, stellate, Kupffer, and other immune cells) can lead to maturation in both function and physical form, or otherwise contribute to maintaining function in ex vivo primary cultures125.
Despite rapid advancements, disparate findings, and evolving developments in replicating hepatic structure and function in vivo, many questions remain outstanding in developing mature engineered liver in vitro126. These challenges include robust, replicable, and throughput-enabling cell sources for each implicated cell type (e.g., primary vs. PSC-derived vs. immortalized lines, etc.), as well as tissue architecture that would recapitulate the niche microenvironment of the target tissue. Further challenges include establishing a chemical environment conducive to gaining and maintaining tissue-level function. Although models are being established for many different hepatic pathologies using currently-available tools (e.g., hepatitis, diabetes/obesity, and cancer127,128,129), their full potential for identifying pathophysiological mechanisms and targets will not likely be realized until they can be maximally differentiated from matured, functional tissue.
Pancreatic tissue
The ubiquity of types I and II diabetes mellitus, as well as gestational diabetes, has spurred significant scientific and financial investment into pancreatic tissue engineering, for both modeling and therapeutic applications. In developing pancreatic models, two families of function are held as primary goals: ductal or nutritional (exocrine) function, and endocrine function130. Towards these goals, in vitro pancreatic organoid models have been developed from both primary sources131 as well as PSC-derived cells132. Both cell sources seem able to recapitulate tissue-level structure and can demonstrate aspects of structural progression, disease modeling, and regeneration; factors implicated in pancreatic maturation are summarized in Table 4.
As with other tissues, recapitulating native pancreatic architecture in vitro seems crucial toward achieving final mature function, and is regulated by numerous effectors, but especially the transcription factor Pdx1, which is differentially expressed via many protocols reviewed elsewhere133,134,135. Furthermore, sympathetic innervation is required for proper development and function136, likely due to the effects of epinephrine on regulating embryonic islet vascularization, in part by maintaining VEGF release within strict limits, which in turn mediates normal function137. The delivery of growth factors and ECM secretion by ECs have been suggested as key mechanisms for this activity137, but are likely complemented by nutrient and oxygen gradients, given the precise niche and metabolic importance of β-cells.
The pancreas undergoes a perinatal or post-weaning nutrient switch which, as with other organs, seems vital to induce higher pancreatic function138,139. However, the mechanisms underlying these specific adaptations or responses are not fully elucidated and are likely complex given the central signal-consolidating and influence of the pancreas in its endocrine and chemical relationships with other organs and tissues. The signaling pathways implicated in the late stages of pancreatic development and maturation seem to be held in common with other tissues; Akt/mTOR activity is implicated in post-embryonic islet secretory functional development and structure134,140. Furthermore, these processes are negatively correlated or reversed under high c-Myc expression, which maintains proliferative activity, and so would hypothetically oppose post-mitotic maturation processes141. As with many secretory cells, β-cells are relatively oxidative to provide sufficient energy flux to maintain insulinar activity. This metabolic capacity is mediated and maintained by ERRγ signaling and is thus highly implicated in islet development and sustained function142. Upstream of ERRγ, AMPK is implicated in both the development and functional maturation of the pancreas143. The Akt/AMPK balance, previously discussed, seems to hold constant in β-cells, with high mTORC1 activity often opposing metabolic specialization (e.g., the oxidative switch) and islet function (glucose regulation) in vivo144; the precise role of AMPK in insulinar activity remains unclear145, but is likely to be involved in functional maintenance, as well as pathophysiological progression, through its downstream effectors and their transcriptional regulation146. Several circulating hormones are likely to influence pancreatic maturation; hydrocortisone increases the expression of insulin, as well as the glucose-sensing molecules that stimulate its release147. Similarly to many other tissues, T3 has demonstrated an important role in embryonic and postnatal islet development and function, potentially due to its signaling effects on metabolic regulation148. Finally, specific GPCRs with lipid ligands may either stimulate or inhibit insulin release via PLC/PKC and adenylate cyclase pathways, including long-chain- and short-chain-specific receptors of FAs, as well as FA derivative-specific receptors with putatively insulinotropic activities, albeit through mechanisms that are not fully elucidated135.
Importantly, function in pancreatic cells is a product of metabolic (e.g., AMP/ATP or ADP/ATP) and Ca2+ oscillations in acinar149, α-150, and β-cells; Ca2+ transient-triggered insulinar release in the latter is well-studied as the phenomenon of “bursting”151,152. Common between these cell types, these Ca2+ transient waveforms and periodicity are highly dynamic, suggesting well-regulated consolidation of signals of various provenance to focus functional output in a dynamic and highly concerted manner. Given the importance of Ca2+ as a secondary messenger in many signaling processes, these transients could conceivably serve a dual purpose in directly activating maintenance of mature pancreatic structure and function. The use of linked tissue compartments on-chip153,154 may also prove to enhance maturation in both pancreatic and peripheral cultures, as linked tissues benefit from pancreatic secretions, while in turn the pancreas receives metabolic feedback as it seeks to establish systemic homeostasis.
Skeletal muscle
Muscle tissue engineering efforts have advanced rapidly in recent years, constructed both to elucidate healthy function or development, as well as pathogenesis155,156. As surgical applications of engineered skeletal muscle tissue remain limited, the targets of most efforts are in models to better understand the development of genetic conditions such as Duchenne’s muscular dystrophy, as well as to obviate atrophy such as in ICU-acquired weakness. The contractility of skeletal muscle, as with myocardium, has spurred rapid focus on 3D culture systems and yielded many organoid designs, including innervated and vascularized models157,158. Key factors implicated in skeletal muscle maturation are summarized in Table 5.
Despite a seemingly similar structure to other striated muscle, myocardium, skeletal muscle presents several functional differences that necessitate important considerations for culture. Mature skeletal muscle fibers dwarf even the largest CMs, with diameters of 50–100 µm, lengths from hundreds of microns to tens of millimeters, and dozens of nuclei. Furthermore, skeletal muscle retains a high anaerobic capacity in contrast to the obligate oxidative metabolism of myocardium; skeletal muscle couples this catabolic capacity with both high gluconeogenic activity to regenerate glycogen, and a large phosphocreatine (PCr) pool in preparation for burst activity. In times of relatively low activity, skeletal muscle relies on the oxidation of glucose and lipids, using the adenylate kinase shuttle and regenerating the PCr pool for non-diffusion-limited transport159,160.
Mitochondrial density, and thus oxidative capacity in vivo are dependent on a trained workload. The stratification of cultured muscle to fast- or slow-twitch phenotypes has been achieved using high or low levels of glucose in culture, respectively161. Furthermore, ostensibly through effects on Ca2+-dependent signaling and/or force-feedback on maturation, streptomycin, which non-specifically inhibits Ca2+ channels (including force-sensitive channels), abrogates a degree of fast-twitch functionality induced by high glucose availability161.
The signaling effectors of engineered skeletal muscle maturation seem to resemble those of myocardium. PGC-1α upregulation increases mitochondrial density, oxidation, and contractile features162. PI3K, Akt, and DAG kinase activation results in hypertrophy163,164,165,166. The mTOR substrate P70-S6K1 is required for increased force but not hypertrophy167, and is a common effector of numerous steroids and growth factors (e.g., testosterone, clenbuterol, IGF-1, etc.) clinically associated with muscular maturation. Similarly, mTOR phosphorylation associated with hypertrophy and mature contractility is caused by electrical stimulation in culture168. Myostatin signaling opposes overdevelopment via Smad3-repression of mTOR signaling163; Smad3 has since been demonstrated to directly oppose PGC-1α activation and inhibit Akt through the expression of PTEN, which antagonises PI3K activity169. Upstream of Akt signaling, and similarly to the heart, skeletal muscle has a large mechanobiological aspect that is important toward maturation in vivo, as measured through hypertrophy and function, in part mediated by integrins and FAK signaling170,171. Additional mechanical signaling occurs through LMCD1 via a calcineurin-dependent pathway172.
It is likely that further developments in myocardial maturation and culture may benefit those domains of skeletal muscle research and vice-versa, given their similar function. However, tissue-specific energetics, mechanics, and physical form will necessitate dedicated study of both, and will likely uncover specificities in optimal culture techniques.
Intestinal epithelium
The complexity of the gut and its ubiquity in whole-body homeostasis has inspired significant and rapidly growing research attention. Perhaps the hallmark of the gut is its largely mutualistic population of a diverse array of bacteria, which lead to a rich list of nutrients that varies with a dependence on the individual host and location along the length of the tract itself. The nutrient diversity of the intestine in vivo leads to a complex interplay between metabolism and export by the intestinal epithelium. Interest in developing new therapeutic approaches for pathologies including Crohn’s disease and irritable bowel syndrome, as well as enhancing personal nutritional benefit, have spurred considerable interest in modeling intestinal physiology in vitro. Indeed, much of our nutrition is derived from anaerobic fermentation of large molecules (e.g., dietary fiber) to short-chain fatty acids (SCFAs) by the resident microbiome173; recapitulating the niche of these flora will therefore likely be necessary to fully recapitulate intestinal function in vivo. To this end, there is a rich field of research ongoing in intestinal tissue engineering which has tended toward 3D organoids174, although both 3D and functional 2D cultures demonstrate specialized applications175. In the pursuit of functional intestinal synthesis, crypt differentiation has been of particular interest as of late176; key factors implicated in intestinal tissue maturation are summarized in Table 6. The unique niche of the intestinal lumen allowing (re)absorption and stable bacterial colonization has greatly directed the formation of unique structures. The latter lends itself to a unique definition of “maturity”; in contrast to most other tissues, “mature” intestinal epithelial cells live only for days before being shed. As a result, the crypt represents a physical gradient of maturation, with villus tips composed of highly differentiated and functional cells, while the protected crypt bottoms maintain constitutive and physiological stem or feeder (Paneth) cell co-populations allowing lifetime epithelial replenishment for intestinal function177. Significant in vivo differences in function exist between the small and large intestines178 which are out of the scope of this review given that intestinal organoid models currently lack the ability to make such distinctions.
The intestinal epithelium is characterized by a dynamic environment with many gradients and biochemical triggers underlying its development. Intestinal epithelial cells sustain a high oxidative flux to metabolically support rapid proliferation and nutrient absorption. This absorption and maintenance of homeostasis coincides with a dynamic environment in the intestinal lumen, and both processes rely on active transport with high energy demands178. As with other tissues, the perinatal period is of considerable importance to the specialization of intestinal tissue, as the metabolic physiology of the developing intestine rapidly changes in response to diet and microbiome179,180, and with very specific spatial regulation181. Finally, the regulation of oxygen availability through PPARγ-activated beta-oxidation of lipids maintains low oxygen tension in the intestinal lumen, which helps to maintain an anaerobic environment and allows for the selection of beneficial microbiota173, again underscoring the importance of structure and perfusion in mediating environmental control and homeostasis in advanced tissue function. As implied above, an oxidative metabolic switch is required for full villus formation and is mediated at least in part by the mTOR/PGC1α-activated transcription factor YY1. Ablation of YY1 compromises the morphology of intestinal epithelium, as does direct pharmacological inhibition of the ETC182, or loss of the mitochondrial transcription factor TFAM183. STAT3 and mTOR signaling are inducible in intestinal cells by IL-2, and are crucial to the differentiation of several mature functional cells in organoids184,185.
The study of intestinal microbiota and their physiological effects has exploded recently; gut flora comprising trillions of bacteria, viruses, and fungi have been identified for far-reaching effects around the body. Most cogently, within the intestines, host-microbe and microbe-microbe interactions alter gut metabolism, absorption, and immunity, which can promote homeostasis or diverge toward disease186,187. A key nutritional role of the intestinal microbiota is in the partial fermentation of primary nutrients to produce SCFAs (chiefly acetate, propionate, and butyrate), which are more readily uptaken by the intestinal epithelium188 and form a non-trivial source of energy for the body, in addition to the monosaccharides, FAs, cholesterol, and amino acids more commonly attributed to nutrition. These SCFAs exert immediate and notable effects on the local inflammasome and epithelial physiology; dietary fibers processed by the microbiome into SCFAs increases intestinal intracellular [Ca2+], mediates epithelial repair through IL-18, stabilizes HIF1, and induces STAT activity189.
Microbial SCFAs also activate GPR109A/HCAR2 in gut epithelium and, similarly to GPR43/FFAR3, this receptor carries complex and possibly tissue-dependent interactions with ketones190. GPR109A activation influences the inflammasome, ion homeostasis, and gut wall integrity and barrier function; this interaction may be critical in the development of colitis/fiber nutrition models179,188,189,190. Furthermore, GPR43 stimulation by SCFAs may also promote antimicrobial peptide production in IECs via mTOR and STAT3 signaling191, which again seem to represent ubiquitous effectors of maturation.
The amino acid l-glutamate is a key nutrient in intestinal metabolism from birth onwards. There is a strong negative l-glutamate gradient from the gut to the bloodstream, due to local oxidation and consumption as a precursor for the biosynthesis of many amino acids; unabsorbed glutamate from the small intestinal lumen can become a precursor for additional SCFA production in the colon178. Perhaps unsurprisingly, evidence is also accruing that other amino acids carry out key physiological signaling roles as well192, suggesting that as tissue-engineered models further evolve, maintaining a range of available metabolites will be key to evoking and detecting precise functionalities.
Due to the essential and ubiquitous role of the microbiome in intestinal function, intestinal/floral co-culture is a rapidly growing field and at the forefront of establishing steady-state colonization models of culture. Models associated with the study of the leading risk factor of fatal gastrointestinal disease, H. pylori, have been developed, along with other commensal or pathogenic bacteria (e.g., Clostridium, Salmonella, etc.), enteric viruses, and different stromal co-cultures to encapsulate intestine functionality in a larger context193,194,195,196. Additional efforts undertaken with immune co-culture within intestinal organoids197,198 suggest great promise, both at dissecting healthy function as well as modeling autoimmune or pathogen-linked disease.
Kidney
Nephrotoxicity remains a significant issue underlying clinical trial failures and drug recalls, as well as a persistent risk, even in marketed drugs; the need for relevant preclinical screening infrastructure and personalized risk profiling for prospective drug regimens has spurred significant effort in developing iPSC models of renal function. The shortage of transplantable kidneys has also spurred significant effort in lab-grown, artificial, or wearable tissue-engineered kidneys. Current engineered kidney organoids appear to recapitulate much of the cellular heterogeneity and basic architecture of the developing kidney199. Moreover, many of these differentiation and spontaneous structuring processes seem to mirror in vivo developmental processes based on molecular identities and gradients; such identified factors are summarized in Table 7. Despite these similarities, organoids do not approach mature function, and may in fact regress or fibrose before gaining such advanced filtering and barrier functions199.
As in other highly differentiated tissues, the structures of the postnatal kidney in vivo are known to be highly oxidative. In fact, the kidney is efficient enough to be a net positive source of glucose during energy restriction (i.e., highly-developed gluconeogenic capacity supported by oxidative phosphorylation). However, there are established heterogeneity of metabolic activity and substrate preference along the nephron, wherein proximal tubules are the most oxidative, given the ongoing need for active transport for resorptive processes; the development of these spatial energetic specialization domains has been linked to Akt/mTOR and AMPK signaling200, and is seemingly maintained by differential PPAR expression201. There is additional capacity for glycolytic futile cycling, potentially for glucose or temperature regulation, or more likely to provide a nutritive buffer for the constantly changing demands on the kidney due to a number of activities (e.g., exertion, eating, drinking, excretion, etc.) that manifest in fluctuating electrolyte levels202.
The signaling effectors underlying the provenance of these spatially distinct functional regions in the nephron remains unclear. Multiple gradients (e.g., chemical, osmolar, nutritive, and oxygen availability) exist and largely correlate in the nephron, which could therefore benefit from many of the signaling paradigms discussed elsewhere in this review, to aid in maturation and stratification. Furthermore, as iPSC-derived renal organoids can spontaneously self-organize into nephron-like structures with little functional pressure and even in the absence of perfusion199, there is likely a large physical or spatial signaling component to early nephric tissue maturation. This spontaneous architecture can be complemented with vasculature that is perfusable after implantation203. Other potential transcription factors at play carry traditionally significant metabolic effects; ERRγ has been demonstrated in renal epithelial cells to mediate mitochondrial development, fueling collecting duct function and resorptive processes (e.g., glucose, amino acids, and electrolytes) in cooperation with hepatic nuclear factor-1β204. Furthermore, early B-cell factor-1 expression is spatially-organized, mediating glomerular structure, podocyte differentiation, and metabolism205. The induction of these effectors is still poorly understood and will likely benefit from the use of new organoid culture systems in concert with traditional loss-of-function studies and high-throughput screens, so as to rapidly identify the key stimuli that seemingly induce and modulate kidney maturation.
Common central signaling trends underlying functional maturation
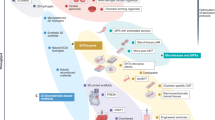
Much of the maturation and developmental progress of tissues have been attributed to Akt and its effectors (often investigated via mTORC1, its subunit Raptor, and its substrates 4E-BP1 and p70S6K) through their enhancement of differentiation, motility, and proliferative cellular processes. This activation has generally been found to be conserved across tissue types, driving direct signaling cascades to regulate cell function through existing infrastructure. In the context of maximizing tissue maturation and functional performance, this supports the existing experimental evidence paradigm that a somatic cell source could be critical to the efficiency and ceiling associated with functional performance in iPSC-derived cells206,207. However, certain roles for Akt (also known as protein kinase B) would seem to preclude it as the sole major driver of maturation, especially as it pertains to metabolic maturation (typically increasing catabolic flux capacity and activity to fuel matured functional energetic demands). This suggests AMP-activated protein kinase (AMPK) as a key player in maturation, due to its role in mitochondrial biogenesis and inducing catabolic pathways to balance and tune the primarily anabolic and anti-quiescent role of Akt (Fig. 2). Throughout this review, common and specific effectors of cell or tissue maturation will be identified, suggesting that the active epigenome and larger patterns of expression are key to specifying the important tissue-specific processes that occur during maturation.

Black arrows represent activation while red hashes represent inhibition; solid lines represent putatively direct interactions while dashed lines represent indirect effects (intermediates not shown or unknown). Image created using BioRender.com.
Consolidators of metabolic signaling: the Akt and AMPK axes
Akt is a central signaling consolidator linking upstream insulin signaling and phosphoinositide signaling (receptor tyrosine kinases, G-protein coupled receptors, focal adhesion kinases, Janus kinases, B-cell receptors, etc.) to control of mTOR, GSK3, MAPK, IKK, nitric oxide, and Bcl signaling, among others; its activation canonically results in cell survival and DNA repair, growth and anabolic metabolism, motility, and proliferation from this wide array of regulators and effectors208 implicated in cell growth, proliferation, differentiation, and alternatively apoptosis. Akt is subject to positive feedback from mTORC2 but potentially negative feedback from mTORC1114,209,210.
In contrast, the heterotrimeric serine/threonine kinase complex forming AMPK is generally perceived to induce widespread catabolic processes that serve to increase intracellular substrate availability and flux. Its catalytic activity is mainly controlled by phosphorylation of Thr172 on the activation loop motif of its α-subunit by liver kinase 1 (LKB1)211 and calcium/calmodulin-dependent protein Kinase (CaMKK)212. The kinetics of this phosphorylation are positively influenced by the binding of AMP to the Bateman domain of the AMPK γ-subunit, which itself is controlled by the AMP/ATP ratio of the cell. This ratio is an inverse indicator of the cells’ energy state. LKB1 itself is activated by PKA (adenylate cyclase produces cAMP), as well as Erk and p90RSK from the MAPK cascade213. CaMKK, in contrast, is activated by increased intracellular [Ca2+] resulting as a second messenger from a number of cell signals. AMPK is also directly inhibited by Akt phosphorylation on the serine-threonine-rich loop at Ser487, but only on AMPKα1 (i.e., the α2 isoform cannot be negatively regulated by Akt)214. This isoform dependency lends itself to state-dependent and tissue-specific mechanisms of controlling crosstalk between pathways. In turn, AMPK both activates TSC1/2 and inhibits Raptor, thus preventing both the propagation of Akt signaling as well as the positive feedback loop of Akt activation through mTORC1/2 cycling215,216,217,218. Unregulated AMPK activation causing continued catabolism leads to autophagy through targets including activation of ULK and inhibition of p70S6K.
By upregulating catabolism, AMPK indirectly regenerates NAD+ by enhancing NADH consumption via the electron transport chain (ETC), which in turn activates the NAD+-dependent deacetylase SIRT1. Important targets of both AMPK and SIRT1 are PPARγ coactivator-1α (PGC-1α), forkhead box O (FOXO), eukaryotic initiation factor-2 (eIF2) and peroxisome proliferator-activated receptor-γ (PPARγ)211,219), which are also control points of phosphoinositide/Akt signaling. Furthermore, AMPK directly activates catabolic pathways (e.g., glucose internalization and glycolysis, FAO), and inhibits anabolic pathways by phosphorylating key control point enzymes220.
AMPK exerts additional significant transcriptional control through the PPARγ-coactivator-1α (PGC1α) signaling cascade, directly or indirectly activating transcription factors including peroxisome proliferator-activated receptors PPARα and PPARβ, nuclear respiratory factors (NRF), and estrogen-related receptors (ERR; see discussion below)16,45; these gene products enhance mitochondrial biogenesis, lipid uptake and beta-oxidation, ketogenesis, gluconeogenesis, and bile production41,73. FOXOs, phosphorylated through AMPK/PGC1α, also upregulate gene products to protect against oxidative stress, both in short-term response to direct oxidative upregulation, as well as the increased oxidative baseline brought on by AMPK-induced transcriptional programs and mitochondrial biogenesis220.
As mentioned previously, SIRT1 is a vital indirect effector of AMPK signaling via catabolic regeneration of its allosteric activator NAD+. SIRT1 increases mitochondrial function via PGC-1α deacetylation, acting through both NRF1/2- mediated mitochondrial biogenesis and CAC/ETC gene upregulation211,221, as well as increasing insulin sensitivity222. SIRT1 is kinetically limited by NAD+ availability223,224, which is dependent on both the energetic and redox states of the cell; stresses associated with both increased SIRT1 activity due to NAD+availability as well as activation of the AMPK/PGC/PPAR axis. PGC1α also enhances MCT expression225, consistent with its role in enhancing catabolic activities. SIRT1 furthermore displays feed-forward expression regulation where it upregulates activity of its own transcriptional activators224. Similarly, SIRT1 deacetylates LKB1, causing nuclear-cytoplasm translocation and allowing it to be directly bound and activated by MO25 and STRAD226, whereupon it can further enhance AMPK activation.
The adiponectin receptor 1 (AdipoR1) is another common means by which cells serve to activate catabolism; AdipoR1 activation increases intracellular [Ca2+], thereby activating CaMKKβ227 which is an effector of the activating phosphorylation event of Thr172 of AMPKα228; AMPK would therefore activate PPARα activity through the PGC1α/SIRT1 axis outlined previously. AdipoR2 in contrast appears to more directly activate PPARα, potentially through increasing its ligand availability, although the precise mechanisms of this action remain unclear227.
While Akt and AMPK are usually portrayed as mutually opposing signalers, with common targets that often provide negative feedback for each other229, there are conceivable instances in which these processes can induce each other230, whether directly or indirectly. For example, under very specific circumstances with exogenous hormones combined with metabolic/hypoxic stress, AMPK initiates Akt activity by inducing ubiquitination231. Perhaps more interestingly, despite their common opposition, both Akt and AMPK activities support glucose uptake and mitochondrial biogenesis232,233,234,235. This is in keeping with their synthetic/anabolic and energy-mobilizing roles, respectively, although direct convergence of these pathways has yet to be thoroughly characterized in the production of robust and efficient mitochondria. Similarly, the complex interplay between Akt and AMPK suggests that the highly regulated physical, biochemical, and functional niches of many developing tissues could in theory benefit from closely regulated crosstalk between these two master regulators of cell function. The convergence of this signaling represents a key control point in the functional maturation of tissues, and one that can be influenced by highly regulated temporal signaling to tune physiology in real time. In the next section, we discuss the cellular digitization of analog signaling, and the role of oscillating signals in the putative balancing of opposing AMPK and Akt signaling processes. The latter are called on to precisely regulate cell maturation and function.
Oscillating signaling in metabolic homeostasis and cell maturation
The absolute ATP content within a cell is low236, meaning that it must be recycled rapidly. Contracting cells, such as muscle, only contain ATP sufficient for a few contractions237, while cells involved in transport, homeostasis, or metabolism for tissue function (e.g., neurons, intestinal epithelium, glomerular cells, pancreatic β-cells, hepatocytes, activated immune cells, etc.) must match a high continuous metabolic drain, generally only sustainable through efficient energetic organization (i.e., oxidative phosphorylation). Such energetic infrastructure would include high capacity for O2 transport to the cell, high/O2-sequestering capacity, high oxidative substrate uptake, high mitochondrial content and substrate import capacity, and direct adenine nucleotide channeling (DANC) or PCr systems to rapidly disperse chemical energy throughout the cell13,238. Cyclic ATP shortages will activate AMPK on a similar oscillating basis, inducing increased [NAD+] and activating SIRT1. Importantly, [NAD+] is de facto tied to the redox state of the cell; changes to redox homeostasis within the cell will necessarily impact AMPK activation. Additionally, there is mounting evidence that oscillations of ERK activation (frequency and amplitude) significantly affect transduction into its physiological effects239,240,241,242.
A key second messenger in many signaling pathways is Ca2+, with direct roles in PI3K/Akt/NFAT pathways, as well as those of certain PKC isoforms. Ca2+ oscillations have been detected in many cell types151,152,243,244, which when compared to modular but flat levels offer notable benefits to these ubiquitous signaling pathways. The functional effect of Ca2+ oscillations is that a binary signal (i.e., [Ca2+]i above or below a threshold for enzyme activation) is evolved into a digital signal modulated by many control points245. Moreover, PKC activity can be sustained well after the dissipation of an initial activating Ca2+ transient due to dynamics of isoform control, diacylglycerol (DAG) availability, and cytosolic trafficking246,247, which increases the information density of a PKC signaling event and which has been studied mostly in the context of neural long-term potentiation. The result of discrete Ca2+ events is very fine control over Ca2+-sensitive transcription or other signaling processes244, with the opportunity for further signaling complexity offered by crosstalk with other signals as discussed above. Modeling efforts have demonstrated that oscillating Ca2+ in the form of cytoplasmic transients will result in very different active patterns of dephosphorylated, DNA-ready NFAT, which will interact differently with second messenger-originating Ca2+ (e.g., from hypertrophic signaling pathways)243. For example, cAMP oscillations can provide filtering to fine-tune oscillating Ca2+ signals248 https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0007189. In the case of endothelium, pulsatile flow in arterial vascular constructs (already known to be associated with endothelial proliferation, function, and patency249,250) induces Ca2+ oscillations of much greater magnitude than steady shear stress251, leading to extensive Ca2+ transient models in endothelial cells (ECs)252 that provide valuable insight into the incorporation of endothelial structures into co-cultured tissue models. Shear forces are well-known to be important in endothelial monolayer proliferation, function, and patency. Furthermore, these Ca2+ oscillations are dependent on VEGF signaling correlated to angiogenesis253. Finally, as Ca2+ channels are well-known to operate with higher-order kinetics with respect to [Ca2+]254,255,256, the ionic environment both within and outside of the cell becomes clear as a dynamic modulator of physiological signaling.
Therefore, interactions of cyclic metabolic signalers could be similarly expected to exert highly specific controls in concert with exogenous growth or maturation signals. For example, increased cytosolic [Ca2+] upstream of the AMPK activator CaMKK should cyclically induce catabolism to fuel contractions in muscle cells, while PKA interacts with AMPK through adenylate kinase257. Similarly, there could conceivably be Ca2+-facilitated PI3K or PKC signaling conflicting with AMPK activation. Considering a muscle cell as an example, PI3K activation can be expected to occur cyclically through integrin/FAK signaling during contractions. These seemingly contrasting signals are likely highly tuned and sensitive in vivo to provide maximum metabolic control, giving further impetus to mirror the (bio)chemical, ionic, and mechanical niches of engineered tissue models to maximize their transferability.
There are obvious difficulties in trying to assess cyclic activation of AMPK, Akt, or other signaling consolidators. Nevertheless, new work has demonstrated a rapid induction of AMPK activation (<30 s) and signaling in newly-activated neurons98, with corresponding long-term effects that presuppose continual AMPK activity. Although many effects of AMPK are direct activations and inhibitions of existing transport and catalytic processes, it is prudent to think that short but periodic upregulations of specific genes will in time change the proteome of the cell appreciably. For example, it could be conceived that short activations of AMPK, above a certain analog threshold, result in a low near-constant digital signal. This signal could manifest in certain long-term AMPK effects resulting from a convergence with Akt activation patterns (e.g., robust mitochondrial biogenesis), while avoiding certain counterproductive AMPK-linked activities in an otherwise maturing or differentiating cell (e.g., low-integrity (leaky or inefficient) mitochondrial biogenesis caused by selective gene expression, or autophagy and excessive nonoxidative catabolism). Oscillations or aggregate effects of cyclic metabolic demand such as exercise could conceivably exert differential effects on AMPK activation patterns258. Similarly, insulin control in the body is a quintessential example of oscillating endocrine signaling, with pancreatic secretions affecting whole-body metabolic homeostasis and carbon sourcing152,259,260; prandial rhythms or nutritional status variation in other hormone profiles would provide additional control points from the same base stimulus (e.g., feeding).
Perhaps one of the most stable and pervasive oscillating signals in vivo is circadian rhythms, which are well-established effectors of maturation in longer timescales. These signals control a wide range of internal cell processes, and greatly influence the secretory processes of different cells, thereby modulating the levels of different hormones and other circulating factors throughout the day, with downstream effects on cellular metabolite and Ca2+ concentrations261. Circadian rhythms and their signals have been implicated in both myocardial262 and β-cell maturation in vivo263. Although little in vitro work has examined the effect due to notable challenges of both controlling and modulating the cellular clock in culture, co-culture models may be able to recapitulate some degree of these interactions, possibly potentiating the impact of advanced culture techniques toward greater maturation.
Physical signaling and cell-cell contact in maturation
The physical environment of cells in their functional development, plasticity, and response to stimuli has been of considerable interest, as differential mechanics are well-known to be both the source and result of a number of pathologies across many tissue types264,265,266,267,268. Cell-mediated physical forces have been extensively identified as critical to cell fate and development from the early embryonic stage269,270, wherein several examples of pathology can be attributed to aberrant processes of mechanotransduction271. To study this, several methods have been developed to deliver specific physical stimuli to organ-on-a-chip models, including fluid shear forces, compression, and stretch to achieve relevant maturation272.
Cell connections and environmental stiffness are well-known to be transduced through integrins and junctional complexes, in some cases dependent on MRTFs and YAP/TAZ signaling through the Hippo pathway268,273, while related Wnt/β-catenin signaling has been well-recognized in the development and maturation in many tissues274,275,276,277,278. Similarly, cadherins and ephrins are implicated in certain specific maturation events279,280,281, and may well be implicated in others as they are targeted for study in the future. Cadherins have specifically been shown to interact with the PI3K/Akt pathway282,283,284, suggesting that exogenous forces can modulate the transduction of other maturative signals that utilize these pathways (Fig. 3). Cell tension and other integrin signaling can be transduced through a focal adhesion kinase (FAK)-assembled complex (containing paxillin, Src, and talin among other proteins) affecting PI3K/Akt, MAPK, Src, and JNK pathways273,285, as well as integrin-linked kinase286,287,288, which can all exert maturative effects.

Black arrows represent activation while red hashes represent inhibition; solid lines represent putatively direct interactions while dashed lines represent indirect effects (intermediates not shown or unknown). Image created using BioRender.com.
Nanotopography and ECM micropatterning have also proven to be a powerful method to exert control on cell fate and function in vitro in several applications288,289,290,291,292,293,294,295, in part by defining boundary conditions and so controlling the directionality of exogenous forces on a cell, as well as by regulating the formation of focal adhesions296. These concepts have been well-illustrated in the model of the intestinal crypt, which features diverse cell types in close proximity and a dynamic environment. Cell distribution and niche is highly organized, with specific gradients along the depth of the villus/crypt297 that likely correlate with a change in stiffness298. In the intestinal epithelium, integrin-linked kinase has been linked to cell motility, differentiation, and function299. Furthermore, an ephrin/Wnt/Notch signaling network has been implicated in both villus/crypt development and intestinal stem cell maintenance/differentiation298. Given the ubiquity of common mechanobiological signaling pathways in developing tissues300, it is likely that analogous processes can be found in other models. Physical signaling in vasculature exerts significant secretory and gradient effects on its environs (as discussed below). Signaling effectors of permeability are generally associated with calcium-dependent pathways (including PKC, Src family kinases, and RhoA)301, as well as the output metrics of polarity and alignment302. Shear forces have been also demonstrated to exert direct effects on function in models of the gastric epithelium303,304 and liver305.
ECM and physical signaling are perhaps most studied in context of fibrosis. Fibrosis occurs in some form across nearly all tissues, and generally represents loss of signaling and mechanical homeostasis followed by the re-establishment of a new equilibrium characterized by myofibroblastic maturation and responses by the other cells present in the tissue306. Although fibrosis has been directly studied in tissue models307,308,309,310, excess ECM deposition or pathological stiffening may need to be monitored in most if not all organoid models as they become more complex due to 3D form, perfusion, and co-culture, as such a positive feedback loop-response to a loss of homeostasis may mask, blunt, or subvert the physiology of models of either otherwise healthy function, or specific pathology.
In addition to cytoskeletal transduction of physical forces via cell-cell and cell-ECM connections, cells can respond to external forces using mechanosensitive ion channels including: Piezo1/2 channels, TREK/TRAAK K2P channels, TMC, and TMEM63. These channels are present and active on a wide variety of tissues including myocardium, vasculature, hepatic sinusoids, bone, lung parenchyma, pancreas, cartilage, multiple glandular epithelia, nervous tissue, and renal tissue and urothelium311,312,313. Additionally, these channels may provide a significant contribution to shear stress mechanotransduction and baroreception311,312,313, of which definitive mechanisms have remained difficult to identify. With specific proven roles in development and maintenance of function (or pathology in their dysfunction), mechanosensitive ion channel313,314 and ROS activity13,315,316 likely exert a role on the maturation of cells and their composite tissues. However, the extent of their signaling activities, and local protein assemblies that mediate their effects, remain an active area of research.
In a tissue, many highly differentiated cells become connected in a functional syncytium via expressions of linking connexins. Connexin isoform expression patterns are tissue-specific, and so provide selective cell-cell interactions based on compatibility. Furthermore, connexin subtype gives control over passage of transjunctional small molecules based on size, shape, and charge; allowing regulated passage of second messengers and metabolites including but not limited to adenosine and its phosphates (cyclic) AMP, ADP, and ATP, and IP3317, as well as Ca2+, ROS, and NO318. Transit of these messengers (and thus oscillatory behavior, see above) between cells is geometry-dependent319, again reinforcing the importance of 3D biomimetic structure and co-culture (for connexons between different cell types), but also the convergence of cues towards holistic maturation (e.g., morphological maturation influences calcium flux between cells, which could have positive feedback toward further maturation, as discussed above). For example, physical pairing of an immature CM to a mature one in vitro can positively enhance the functional maturity of the immature one without further intervention320, suggesting the importance of cell connectivity in signaling.
Molecular effectors of cell and tissue maturation
Although each tissue or cell type may respond more to certain key factors playing outsized roles in development, the realities of cell and tissue maturation (i.e., higher cell functionality) generally requires increased metabolic flux. As mitochondrial oxidation is both higher-yield in terms of ATP flux as well as more efficient in terms of energy extraction, most maturing tissues must activate mitochondrial biosynthesis; however, substrate preference can be more muted between tissues based on nutrient availability in vivo. Not surprisingly, the endocrine, paracrine, and physicochemical signals of the cell niche that induce these metabolic maturation processes are often conserved between tissues.
A postnatal switch in metabolism and the level of function occurs in many organs; postnatal shifts often coincide with weaning in animal models138, and may be associated on a tissue-specific basis with hormonal windows or profiles, nutrient shifts, changes to oxygen availability, and microbial exposure. This early generalization and later specification to an organism’s state may be reconciled on the level of natural selection, namely that organisms must adapt to specific challenges in their environment and specification; full functional commitment to a given nutritional or signaling niche before birth may disadvantage a newborn due to the resources needed to remodel tissues (e.g., based on available diet/nutrition, hormones in milk, etc.). This flexibility enables the high level of postnatal plasticity observed in many tissues, and is encouraging to tissue engineering efforts with the goal of replicating native tissue function; many embryonic windows and specific signaling events may not need to be met if the rich and specific postnatal niche is achieved in vitro to guide tissue-specific maturation. However, a ceiling on cell function, based on early stimuli, may exist. Similar to the phenomenon of epigenetically-enabled somatic cell memory as observed in iPSCs, oxygen availability321 and mechanical environment293, even at the pluripotent stage, can greatly influence functional performance.
Ultimately, despite tuning sensitivity toward many active maturation factors, those signals must be present to exert their influence, whether on an intracellular, autocrine, paracrine, endocrine, interstitial, or otherwise circulating basis. An example of metabolic regulation influencing receptivity to other maturation signals can be found with T3, a common effector of maturation. Thyroid hormone has numerous roles in tissue maturation and continued systemic metabolic regulation,50,322, and has been shown to be essential for the development of virtually all organs, including kidney, liver, neurons and BBB, other vasculature, and heart322,323, with evidence for very specific temporal and regional regulation, at least in brain323. T3 is transported at least in part via monocarboxylate transporters. Relatively little is known about the regulation of T3-active transporters, although a review of the available evidence suggests downregulated expression of transporters in both fasting and inflammation324. Alternatively, the nutritional state of the cell and the general endocrine/paracrine environment would certainly be relevant in T3 homeostasis. Even after the dynamics of T3 internalization, it must exert its effects in another dynamic system; the thyroid hormone receptor (THR) binds other nuclear receptors such as the retinoid X receptor325. However, in the absence of ligand, THR complexes with nuclear receptor co-repressors 1 and 2, among others. These co-repressors, once bound, oppose many transcriptional activities, including those of ERRs326; this suggests that THR activation may potentiate the pro-maturation effects of other pathways that are described in this text.
Glucocorticoid receptor (GR) biology has been noted as contributing to metrics of maturation in multiple tissues; several GR ligands have been identified to regulate downstream signaling by NF-κB, AP-1, STATs, and PPARs, among others118. This makes GR signaling an immediate subject of interest for possible synergistic, conflicting, or potentiating interactions with the other key pathways in maturation discussed above. Additionally, PPARα interacts with GRs; although GRα internalizes and activates genes implicated in lipid and protein metabolism, it also upregulates gluconeogenesis. Simultaneous PPARα activation will silence the transcription of genes regulated by glucocorticoid response elements327. However, at the same time, PPARα activation enhances active GRα binding and blockage of NF-κB and AP-1, and their associated transcriptional effects327, suggesting that the balance of signaling in terms of transcription factor activation is highly context-dependent and likely dependent on crosstalk from concurrent signaling processes. Continuous GRα activation is implicated in insulin insensitivity by decreasing insulin receptor substrate (IRS) expression328; this could be viewed as a positive reinforcing loop, as decreased insulin signaling will then result in even less downstream activity. PPAR binding of lipids329 is thought to stabilize protein-protein interactions and response element binding, although the low affinity may mean that lipid activation of PPAR is not significant under physiological conditions329,330.
The soluble and orphan (i.e. unknown endogenous ligand) ERR also activate PPARα/β response elements16,41. Although the ligands for ERRs are far from settled, ERRα activation by cholesterol has been established331, and other steroids or lipids remain potential ligands. The metabolic response to ERR activation332 is both dependent on receptor subtype and tissue in question, the latter ostensibly results from epigenetic influence or crosstalk with other active regulatory pathways. For example, in the hippocampus, ERRγ is heavily implicated in memory function through its enabling of greater glycolytic and glucose-based oxidative flux333, while in the heart ERRα/γ have been shown to downregulate glycolytic genes and simultaneously upregulating (FA) oxidative function45.
Finally, metabolic and maturation effects can often be attributed to one or more of many PKCs, with different dependences on Ca2+ or DAG (produced via phospholipase C activity) for activation, having downstream effects on motility and adhesion as well as FA storage and metabolism334. These widespread signals include both positive and negative interactions with insulin signaling through multiple control points, including the IRS, Akt, and other poorly elucidated targets. The net result of PKC signaling is, therefore, isoform- and tissue-dependent, and is likely afforded great flexibility due to the range of PKCs whose expression can be induced, which have been reviewed in detail elsewhere335,336. PKA and JAK/STAT signaling intersect with PKC337 and AMPK338,339,340 as well, especially at the level of transcription factor effectors.
Metabolic signatures: substrate signaling fueling differentiated tissue function
During tissue differentiation and maturation, many cells reduce their endogenous biosynthesis of essential substrates and cofactors, relying instead on vascular supply of nutrients, hormones, and other signaling-active molecules from metabolic organs (e.g., the liver, pancreas, GI tract, etc.). Depending on the tissue in question, common carbon sources for energetic usage include glucose, lactate, lipids, ketones, or amino acids. However, substrate use, at steady-state, requires that bioenergetic and biosynthetic intermediates (NAD+ and NADP+, respectively) be regenerated to equilibrium with the cellular redox state, the former via either mitochondrial oxidation or anaerobic fermentation to lactate, and the latter via the completion of reductive anabolic steps. The preferential use of specific carbon sources, and the pathways implicated in this regulation, are highly dependent on the nature of the tissue function. Below we discuss tissue-specific examples of how cell types require a metabolic niche, and how the resulting metabolic niche of a tissue in turn affects signaling and function. Even signaling-inert molecules (i.e. non-ligands) can indirectly modulate relevant pathways by affecting key indices that influence homeostasis, such as pH, AMP/ATP ratio, NAD+/NADH ratio, redox status, etc. In the next sections of the review, we highlight both the central mechanisms underlying maturation, and several convergences with metabolism that can impact the functional phenotype of the tissue.
Similar to initial carbon sources, many intermediate metabolites or biological reductants are signaling-active; including intermediate carbon sources such as acetyl-CoA, succinyl-CoA, AMP/ATP, and glutamine341, likely among many others. Other metabolically-associated small molecules can influence the signaling state through simple equilibrium effects; increased acetyl-CoA concentrations in turn kinetically upregulate acyltransferase activity190. Similarly, changing the NAD+/NADH balance through redox state or nutrition can change sirtuin family deacylase activity342, as discussed above. More directly, many carbon sources can induce direct signaling cascades within targeted cell types. For example, ketone signaling can be extensive both via direct receptors, and via downstream response effects due to the implied metabolic state that influences ketone circulation and metabolism to begin with. Ketone signaling has been observed in neuronal, intestinal, vascular, adipose, and liver tissue190; acetoacetate activation of GPR43 indicates convergences with that receptor’s canonical lipid sensitivity343, and has also demonstrated crosstalk with spatial (Notch) signaling in intestine344. Another common ketone body, β-hydroxybutyrate (β-OHB), inhibits certain histone deacetylase activity and in doing so activates oxidative stress defenses to enable oxidative metabolism345, but conversely can also inhibit lipolysis in adipocytes via activation of GPR109A/HCAR2 (hydroxycarboxylic acid receptor 2, also known as niacin receptor 1)190,346. The antioxidant activity of β-OHB has been attributed to downregulation of NF-κB, ostensibly through IP3 activation and intracellular [Ca2+] increase. These signals will in theory interact with the Ca2+-sensitive processes identified above, and therefore be sensitive to cytosolic Ca2+ oscillations. Furthermore, GPR109A activity leads to both MAPK and PPARγ activation347, exerting additional control points on energetic and exogenous signal-linked pathways, as described previously. Given ketones’ roles as a metabolite of last resort during periods of low supply (e.g., fasting or starvation)346, it is likely that there are many ketone-dependent signaling processes organized per tissue that have not yet been fully characterized in context.
Design strategies for complex processes: future directions for search processes, culture practices, and optimization strategies
To date, the study of in vitro tissue maturation has revealed few shortcuts to high functionality, suggesting that efficient and effective maturation is a multi-system process. As outlined in the early sections of this review, the convergence of tissue-specific cues, circulating nutrients and endocrine signals appear to coordinate progression across tissues to genetically-encoded checkpoints348. However, the available evidence suggests that functional maturation within a specific tissue is largely a holistic process, and that advancing one phenotypic metric is likely to advance others, both via shared transcriptional organization and through the development of cellular microdomains, and due to the self-reinforcing paradigm of cellular energetics. Advanced function requires specialized metabolism, which will only occur through metabolic insufficiency stress and ROS production, while increased energetic capacity will enable biosynthesis, structural complexity, and thus tuned and advanced function.
As a result of our limited understanding of the processes, pathways, and checkpoints underlying tissue maturation, it appears difficult to reach these functional milestones in vitro with current capabilities. Moreover, existing tissue engineering optimization strategies do not necessarily have the predictive power of novel approaches, either in interpolation or extrapolation. Below, three generalizable and actionable strategies for optimization of tissue maturity are outlined and discussed in depth. They include: replication of a tissue’s (bio)chemical niche, perfusable 3D co-culture, and design of experiments to account for biological complexity.
Tissue-matching of the (bio)chemical environment
The (biochemical) balance of an in vitro environment is a longstanding and common method of biological model optimization, pioneered with the introduction of Ringer’s solution in the 1880s, continued with Earle’s salts in the 1930s, and providing a foundation for development of many common cell culture media in the 1950s-60s. Serum (e.g., fetal bovine, fetal porcine, or calf sera) has been traditionally used to provide both complex nutritional supplementation and growth factors, but carries several important limitations. Undefined culture media (e.g., containing serum at 5–10% v/v or greater) are not replicable and introduce batch effects into experiments349. Moreover, high medium serum content induces toxicity, whereas low serum does not provide enough essential nutrients at biomimetic levels350. Efforts to minimize the presence of effective but undefined growth factor content include the use of adult or supplemented calf sera, which are generally less effective for cell growth. In recent years, the rise of defined (i.e., containing isolated or purified xenogeneic components) and chemically-defined (i.e., containing only recombinant products beyond organic small molecules and inorganic components) supplements to ubiquitous basal media have overcome the use of serum-derived complete media for both replicability and translatability concerns. Several such supplements of note include the B-27 supplement, which was developed for neural culture but has been recognized for its efficiency in the maturation of diverse tissues351,352,353. Other supplements include insulin-transferrin-selenium, creatine-carnitine-taurine, serum replacements (such as KnockOut™), lipid-enriched and/or recombinant albumins, and chemically-defined lipid supplements, in a growing field of defined supplements354.
The near-ubiquitous use of media unspecialized to the ionic and nutritional niche of the cell or tissue in question has been recently examined355, warning that cell physiology may be compromised without carefully designed media for the situation at hand. In the context of tissue cultures, this suggests a benefit to the use of a conscious choice of basal media, and/or the use of properly perfused tissues with endothelium or co-cultures that benefit from vascular barrier function and ion-regulation (see below). Similarly, tissue function may be compromised in medium such that they cannot regulate their environment as expected in vivo. For example, astrocyte-mediated regulation of neural tissue extracellular fluid is extremely precise356. Changing the niche and scale, where the mass of cells cannot adequately regulate within said niche, would change the metabolic demands and thus the functionality of the cells.
Traditional culture media are also notorious for physiologically irrelevant levels of glucose in media (commonly 25 mmol L−1). For short-term cancer cell culture these levels may be largely inconsequential; however, the effects on putatively healthy cells with altered homeostasis may currently be underappreciated355, and could elicit magnified effects when used in delicate and interbalanced co-cultures. Included in these concerns are worries of oxidation and interactions with insulin-related signaling and effects, if present. Glycation can occur to functional effect on extracellular proteins within weeks357, which could in theory affect ECM, receptors, channels, or other extracellular-presenting proteins. Little work has been done on glycation or oxidative degradation of media, and most of it has been in the field of bioprocess; as a result, proliferation or recombinant protein production are the functional metrics of interest that show sensitivity to these processes and suggest that other emergent functions could as well358. Although advanced glycation end products can take time to appear, they may have rapid effects on cell function once present359; this is of particular interest not for traditional proliferative cultures, but for quiescent and highly functional tissue models.
The issue of glucose over-availability would not likely be addressed by simple replacement by a large quantity of a different carbon source, as toxic effects of other metabolic substrates may also exist; lipid supplementation can result in the relatively unelucidated phenomenon of lipotoxicity41,360, which generally manifests in ER, mitochondrial, or Golgi stress. The pathways of this toxic response can include JNK/Bcl2 family signaling as well as the well-known ER stress signaler PERK, leading to pro-apoptotic ATF4 activation361. Lipids also interact with cyclooxygenase and cytochrome P450 activities, both of which can elicit ROS production, or may result in unintended metabolite production, such as the ubiquitous FA, arachidonic acid, and its eicosanoid products362. Lipids may however carry additional benefits to complex co-cultures beyond specific functional effects; reports of lipid supplementation enhancing the secretion of angiogenic factors by mesenchymal stem cells363 suggests feedback loops when perfused co-cultures are combined with well-optimized culture media.
The use of carbon sources may need to be considered on a case-by-case basis, such as in personalized medicine screening. Ketone metabolism is unsustainable to certain CoA-transferase deficient patients in vivo346, and can be incompatible with certain metabolic or signaling states even in healthy tissue/patients. The use or avoidance of ketones may be important in eliciting a desired response, or in mitigating an unintended phenotype. These patterns may also hold true with lipid-related CoA-transferase or carboxylase deficiencies.
Tissues benefit from co-culture: 3D structure, cell-mediated culture benefits, and gradient effects of perfusable vasculature
Tissues generally require a selection of cells to maintain utility; a complex mix of endothelial cells for perfusion, fibroblasts for interstitial maintenance, immune cells, and often specialized structural cells support the function of the nominally “functional cells” of a tissue. These cells often provide paracrine or mechanical support in situ, such as endothelial secretion of potent growth factors, and immune-directed tissue remodeling. In this way, the complementary functions of different cell types can drive each other to advanced function. There is compelling evidence that the inclusion of immune cells may be a powerful tool for tissue maturation due to their complex cytokine secretion profiles, and regenerative or pro-maturation programming paradigms125,197,198,364,365. However, there has not been widespread inclusion of immune cells in functional tissue models to date due to several factors, including difficulty in cell sourcing, complexity of phenotypes in tissues of interest, and lack of effective techniques to include immune cells in culture without inducing unintended response patterns. The growing recognition of the role of the immune system in healthy function and development, as well as pathogen defense, is likely to spur significant advances in our understanding of how both to harvest and implement immune cells in co-cultured engineered tissues.
The benefit of perfusable vasculature has been mentioned in several engineered tissue types above in the context of their respective secretomes and nutrient selection for cells outside the vascular lumen. Engineered vasculature may enable nutrient and oxygen supply in larger 3D cultures104, which through mechanical signaling alone can significantly influence cell fate. Subtle variations to nutrient and oxygen gradients can also have outsized effects366,367. Vasculature provides selective barrier function, and a massive secretome of growth factors, cytokines, and other small molecules. Moreover, endothelial function can be modulated by the perfusing chemical environment or the main functional cells of the tissue, such as CMs368, which will then feedback into emergent tissue function; endothelial permeability and smooth muscle tone can be modulated by histamine and platelet-activating factor, radicals such as nitric oxide or superoxide, and even nutrients such as ketones347. Oxygen gradients established in perfusable tissues have been previously discussed, and can differentially activate HIF1/2α signaling, which has been implicated in the differentiation, maturation, and maintenance of healthy tissue function in many cases369.
The inclusion of vasculature or considerations of mechanical replication of tissue features seemingly necessitates 3D culture, which is most typically aided by the use of hydrogels or scaffolds that are largely generated through synthetic means or using decellularized native ECM; polymeric scaffolds or gels will ultimately offer the highest batch replicability and lowest propensity for contamination, toxicity, or unelicited and divergent cell responses. However, 3D tissues also necessitate perfusion, as oxygen gradients to the interior of an engineered tissue quickly become unsustainable. Oxygen availability is not an issue limited to 3D culture; depending on media depth, specific metabolic rate, and cell density, static monolayer cultures can experience significant hyperoxia or hypoxia34. Paradoxically, either condition may induce ROS production34,370. While the source of ROS under hyperoxia is clearly driven by equilibria, hypoxia (a lack of oxygen) may also induce the production of superoxide, as ETC complexes are forced to remain in a reduced (i.e., electron-rich) state371,372; increased ROS may either cause a toxic response in the face of insufficient mitigation, or require a constant and significant metabolic expenditure in the form of biosynthetic reductant (NADPH) to maintain antioxidant defenses depending on the cells or tissue in question. Mature tissues, which rely heavily on oxidative flux to maintain function, will likely have developed significant cellular infrastructure that is then allocated to maintaining ROS within acceptable levels; ROS are not uniformly harmful to cells and are key in homeostatic signaling in healthy tissue373,374. Therefore, the appropriate provision of substrates, cofactors, and antioxidants in the media may need to be optimized for the tissue model in question as insufficient levels of ROS may equally inhibit nominal cell function or maturation processes375.
Experimental design to reflect and harness biological complexity
In recent years, the ability to craft in vitro tissues of functional relevance has increased rapidly, and commensurately with advances in cell and developmental physiology, microfluidic and organoid culture, and biomaterial development. However, recapitulating the dynamic nature and inherent complexity of a living functional system is extraordinarily difficult, and reaching higher levels of mature function has to date required progressively greater investment of time and resources in cell biology and biomaterials science. Moreover, the analysis and benchmarking of further-matured engineered tissues requires greater precision, resolution, and standardization in methods, all of which require greater investment and training in relevant equipment and techniques. As these burdens become greater, the ability to screen cultures by traditional means may present a serious barrier to continued innovation in tissue engineering, or ever fully achieving high-throughput tissue engineering in a non-industrial setting. Furthermore, experimental infeasibility may impose limitations on innovation where progress to tissue maturity may represent more of a sigmoid of function against time, where the upper bound is not adult in vivo function but rather limited or idiosyncratic function. However, this putative limitation may be more likely to be realized under widespread continuation of the prevailing traditional prototypical design strategies (one-at-a-time, single version, or design-of-experiment processes) of tissue engineering, that struggle to model all of the dynamic and interrelated processes of a living cell or tissue (i.e., a complex system, characterized by nonlinear responses, interaction between factors or outputs, or hidden variables). The physiological complexity of any functional tissue likely precludes its engineering by traditional design methodologies, e.g., without the use of significant screening phases for relative cell composition and source, the inclusion of ECM and other biomaterials, perfusion and oxygenation, and (biochemical) niche. In these screens, simple morphological, expressional, and transcriptional metrics might generally be replaced in scoring regimens by emergent functional metrics specific to the tissue in question, reflecting the final goal of mature tissue. When multiple such metrics are available, decisions must be made concerning how to construct a complexed desirability function, and whether iterative decision making is valuable during an optimization.
Predicting or realizing a specific defined signaling state related to maturation may be exceedingly difficult, due to crosstalk between signaling pathways, second messengers, and the mutability of the related physiological decoding processes or responses (Fig. 4), such as the previously discussed balance between the master signal consolidators Akt and AMPK (Fig. 4a). For example, the response of a cell or tissue to a complex signaling input can be reliant upon the receptor that decodes it and the kinetics of the response process itself (Fig. 4b)376, while composite signals can be highly interdependent (Fig. 4c)248. Depending on the kinetics of the response process, this means that the output level of the response can be constitutively elevated or depressed (e.g., cumulative amplitudes of signals giving rise to the slow downstream process example in Fig. 4c, which might be elicited in cell fate, expressional, secretory, or migratory functions that occur on timescales that are many times longer than a discrete signaling event), or subject to different output frequencies or magnitudes (e.g., the fast, thresholded downstram process). A putative example of the difference between these modalities of response might be found in the myocardial force response to filling. Here, fast [Ca2+]-induced beat-to-beat variation in mature myocardium377,378,379,380, has been shown to be mechanistically distinct from slow (e.g., minute-to-minute scale) adaptations to preload381,382,383; the ability to mount these responses is intrinsically tied to the healthy function of mature myocardium. In the absence of a well-validated signal definitively correlated to maturation, even establishing the correlation of any one measurable signal (e.g., a (de)phosphorylated signaling enzyme, a second messenger concentration, transcript or protein expression, etc.) within relevant signaling cascades may become opaque. The use of prescribed or otherwise actionable signaling states, especially as maturation levels increase and in theory become ever more highly- regulated and sensitive to disruption, might lose utility or even become misleading without the use of neural networks or other AI methods necessary to dissect complexity, in combination with large relevant datasets. For this reason, metrics of cell or tissue physiological function provide a relatively holistic, robust, and correlative insight into maturation state and progress, and are likely to be of more use in optimization workflows of maturation in tissue engineering.

a Opposing and aligned roles of Akt and AMPK in cell processes. b Oscillatory inputs (e.g., [Ca2+] or [cAMP]; top) can be decoded on an amplitude-dependent (left) or frequency-dependent (right) basis based on receiver process kinetics being slow or fast, respectively, lending further mutability to the physiological manifestation of a set of interdependent and multi-step signaling pathways376. Reproduced with the express permission of the publisher and authors. c Simulated interactions between calcium and cAMP levels in a model of mutual enzymatic activation, based on active phospholipase Cβ (PLCβ) concentration and a set adenylate cyclase concentration of 0.17 µM248; demonstrating interdependency of second messenger kinetics on their ultimate manifestation as an output response. Reproduced under a CC-BY license with express permission of the authors. d Sample visualization of an iterative High Dimensional, Differential Evolutionary (HD-DE) algorithm with successive generations outperforming each other on a normalized objective metric, with biological variability represented by temporary differences in performance between independent iterative runs that ultimately converge at highly-performing solution scores; this algorithm was used to optimize hematopoietic stem cell expansion medium efficacy388. Reproduced under a CC-BY license with express permission of the authors.
Traditional optimizations focusing on one factor at a time can fail to model or account for important multi-factor processes that occur in complex systems. However, tissue engineering and biology in general are notorious for uncontrollable or unidentified factors, batch effects, and higher-order interactions that get missed using simple experimental designs or optimizations. Factorial designs (e.g., Design-of-Experiments methodologies) have been implemented to a degree in tissue engineering studies, but are often constrained to very high-level problems and have varying degrees of success at optimization, due to throughput limitations in testing experimental runs which scale exponentially with the factorial complexity of the system in question; these optimization experiments also tend to lose resolution quickly with experimental variability. However, other more-“established” (i.e., greater industrial footprint, formalized practice, and longer history as a recognized field) engineering fields make varying use of many tools to account for complex systems, including fuzzy decision making (i.e., decision making in an environment where the goals or constraints are imprecisely or otherwise vaguely designed or understood; notably a decision problem can be fuzzy even in the presence of highly quantitative or absolute data inputs), the use of objective metrics to encapsulate classes of function, iterative design-benchmarking-evaluation cycles, and holistic multi-criteria optimization (e.g., a weighted composite desirability function as the objective function384, or the use of Pareto fronts to identify acceptable trade-offs between different desirable outcomes385,386,387).
The challenge of efficiently performing high factorial optimizations, as described above, lies in the presence of nonlinear factor main effects, hidden interactions between factors, and the need for many experiments to statistically deconvolute a predictive model. An option for such problems in tissue engineering projects may lie in the adoption of biologically-inspired natural selection or directed evolution algorithms as embraced for chemical, molecular biological, and bioinformatic workflows, or a combination thereof (Fig. 4d)388. These processes hold in common that they do not seek to model the precise relationships between all inputs and outputs, but instead assess output functionality. In complex system, the harnessing of complex factor interactions and the use of limited stochasticity can enable a project’s benefit from iterative successes while navigating away through the solution space from failures. Separate from biological complexity, the realization of highly functional tissue-engineered constructs may involve significant handling of fuzzy problems, which are well-documented in other engineering applications385,389. Although the two concepts are often correlated or conflated, fuzziness in the case of tissue engineering would denote imprecision in the necessary and robust functional behavior toward maturation of a tissue model for downstream application, and in practice represents consensus on an acceptable end product without agreement on how to achieve it, and may conflict with the use of even multiple low-level quantitative physiological metrics with unclear connection(s) to emergent functional phenomena (e.g., the use of multiple cytoskeletal transcript counts as opposed to a contractile endpoint). To date, given the wide range of physiological metrics, both functional and not, fuzziness tends to increase when correlational metrics are more heavily relied-upon (e.g., expressional and transcriptional markers, morphology, etc.); whereas absolute metrics e.g., the peak contractile stress of a myocardial construct as a percentage of adult ex vivo sample stresses) tend to reduce the need for fuzzy decision making, and absolute thresholds can be constructed (e.g., being able to recapitulate >50% of adult contractile stress would be acceptable to stakeholders to leave the developmental cycle and begin using a model for downstream patient-facing applications). In either case, multiple parameters of a system, both functional and logistical, are constantly being weighed against each other during experimental design, either explicitly or implicitly (Table 8).
An alternate way to produce a Pareto-optimal solution toward two or more correlated but not identical outputs is using multiple decision makers386, which will closely approximate the output, if not methodology, of a composite desirability function. However, the use of decision makers still intrinsically includes an element of fuzzy decision making, and may provide a valuable workflow incorporating highly specialized advisors (e.g., a microfluidics engineer and an internal medicine physician) balancing their respective priorities in the realization of a functional tissue model. The use of one or more decision makers can be further formalized in interactive and iterative multi-objective optimization workflows387 that alternate phases of decision making with optimization. This in turn allows for response surface exploration during the developmental process; the non-orthogonality of factors and potential needs for variate re-weighting, assay re-optimization for a changing tissue, or sensitivity or variance testing are potential concerns for tissue engineering process optimizations due to biological complexity. Ultimately, certain objectives may come in conflict with each other, or the relative priority of one objective over another is unclear. In these cases, a Pareto-optimal solution that is acceptable to all stakeholders can be established through defined means385,389. For example, the set of possible solutions can be pruned or iteratively sorted by re-weighting existing parameters, introducing new factors of interest to increase dimensionality in a more focused solution space, or assessing logistical capacity or technical thresholds of interest.
Regardless of the level of complexity and fuzziness in a tissue engineering problem, the objective functions used for optimization will therefore likely be most effective when they represent a reference to or recapitulation of emergent physiology or relevant clinical metrics (especially in comparison to reference ex vivo metrics where available and appropriate). These metrics may include, depending on the tissue in question, protein or hormone excretion, ion reuptake, metabolic processing, barrier function and selectivity, electrophysiology waveforms and metrics, or contractility. Aside from best experimental practices in using functional metrics for designing engineered tissues, the best practices in optimization statistics and decision making under complexity, and especially in the case of multiple competing outputs, therefore recommend stepwise, stakeholder-driven and -corrected trajectories385,386,389. This can include re-weighting of composite desirability metrics, switching objective metrics, and insertion or termination of seeds/paths, depending on the model of decision making used. As the difficulty in advancing tissues in functional maturity increases (a trending theme made clear in the early sections of this review), the incentive to perform highly economical and well-planned experiments will correspondingly increase. As with other fields of engineering, the introduction of formal development methods that allow for multivariate optimization and the harnessing of complexity will likely produce significant advances to tissue engineering and its downstream applications.
Conclusions
The mechanisms underlying functional tissue maturation in vivo require further elucidation, and currently this lack of actionable understanding may limit the development of highly functional engineered tissues for use in drug testing, basic science, or even graft applications. However, the use of systems biology approaches both in vivo and in vitro have yielded notable mechanistic understanding and innovation in tissue engineering studies. As a high level of function is invariably energy-intensive, most highly functional tissues (that are of immediate interest for engineering) are oxidative, and therefore rely on common trends of strict morphological regulation, high mitochondrial biogenesis, maintenance, spatial organization, and intracellular energetic shuttling to active organelles or regions. We hypothesize that by incorporating considerations of preferred substrates, hormones, cofactors, and potentially synthetic signaling-active molecules, a robust and self-perpetuating homeostasis can be established to push engineered tissue to higher levels of functional maturity. This goal may be accelerated by, or may require the use of, 3D organoids, perfusable vasculature, and/or co-culture systems to recapitulate a base level of function.
Finally, the inherent complexity and difficulty in articulating quantitative goals for adoption of a tissue-engineered model for downstream applications may pose a limit to the relatively simple experimental designs, design frameworks, and object-based decision making commonly used in tissue engineering today. The aims of the tissue engineering field instead likely necessitate the adoption of optimization and decision making frameworks common in other engineering fields. We have introduced potential study and decision paradigms and frameworks that may be of value in attempting to produce highly mature tissue models.
The ultimate goal of tissue engineering will be to mirror healthy and pathological function with near-indistinguishability from in vivo physiology. However, these ambitious goals do not need to be fully achieved before engineered tissue function becomes clinically relevant. The widespread introduction of iPSC-derived cells to replace animal models for preclinical drug screening aims to reduce both false positives and false negatives for both efficacy and toxicity endpoints. Moreover, the specifics of many human pathologies will never be fully recapitulated in animal models. By appreciating and accounting for the complexity inherent in living tissues, the field can advance models that interpret environmental stimuli and establish for themselves a homeostasis that recapitulates full mature function.
References
Pound, P. Are animal models needed to discover, develop and test pharmaceutical drugs for humans in the 21st century? Animals 10, 2455 (2020).
Onakpoya, I. J., Heneghan, C. J. & Aronson, J. K. Post-marketing withdrawal of 462 medicinal products because of adverse drug reactions: a systematic review of the world literature. BMC Med. 14, 10 (2016).
Ferri, N. et al. Drug attrition during pre-clinical and clinical development: understanding and managing drug-induced cardiotoxicity. Pharmacol. Ther. 138, 470–484 (2013).
Vunjak-Novakovic, G., Ronaldson-Bouchard, K. & Radisic, M. Organs-on-a-chip models for biological research. Cell 184, 4597–4611 (2021).
Hofer, M. & Lutolf, M. P. Engineering organoids. Nat. Rev. Mater. 6, 402–420 (2021).
Yao, G. Modelling mammalian cellular quiescence. Interface Focus 4, 20130074 (2014).
Gintant, G. et al. Use of human induced pluripotent stem cell–derived cardiomyocytes in preclinical cancer drug cardiotoxicity testing: a scientific statement from the american heart association. Circ. Res. 125, e75–e92 (2019).
Colatsky, T. et al. The comprehensive in vitro proarrhythmia assay (CiPA) initiative — update on progress. J. Pharmacol. Toxicol. Methods 81, 15–20 (2016).
Feric, N. T. & Radisic, M. Maturing human pluripotent stem cell-derived cardiomyocytes in human engineered cardiac tissues. Adv. Drug Deliv. Rev. 96, 110–134 (2016).
Feric, N. T. & Radisic, M. Strategies and challenges to myocardial replacement therapy. Stem Cells Transl. Med. 5, 410–416 (2016).
Romagnuolo, R. et al. Human embryonic stem cell-derived cardiomyocytes regenerate the infarcted pig heart but induce ventricular tachyarrhythmias. Stem Cell Rep. 12, 967–981 (2019).
Montgomery, M. et al. Flexible shape-memory scaffold for minimally invasive delivery of functional tissues. Nat. Mater. 16, 1038–1046 (2017).
Piquereau, J. & Ventura-Clapier, R. Maturation of cardiac energy metabolism during perinatal development. Front. Physiol. 9, 1–10 (2018).
Lopaschuk, G. D., Collins-Nakai, R. L. & Itoi, T. Developmental changes in energy substrate use by the heart. Cardiovasc. Res. 26, 1172–1180 (1992).
Lopaschuk, G. D. & Jaswal, J. S. Energy metabolic phenotype of the cardiomyocyte during development, differentiation, and postnatal maturation. J. Cardiovasc. Pharm. 56, 130–140 (2010).
Vega, R. B. & Kelly, D. P. Cardiac nuclear receptors: architects of mitochondrial structure and function. J. Clin. Invest. 127, 1155–1164 (2017).
Glancy, B. et al. Power grid protection of the muscle mitochondrial reticulum. Cell Rep. 19, 487–496 (2017).
Bedada, F. B., Wheelwright, M. & Metzger, J. M. Maturation status of sarcomere structure and function in human iPSC-derived cardiac myocytes. Biochim. Biophys. Acta 1863, 1829–1838 (2016).
Callaghan, N. I., Hadipour-Lakmehsari, S., Lee, S.-H., Gramolini, A. O. & Simmons, C. A. Modeling cardiac complexity: Advancements in myocardial models and analytical techniques for physiological investigation and therapeutic development in vitro. APL Bioeng. 3, 011501 (2019).
Karbassi, E. et al. Cardiomyocyte maturation: advances in knowledge and implications for regenerative medicine. Nat. Rev. Cardiol. 17, 341–359 (2020).
Ronaldson-Bouchard, K. et al. Advanced maturation of human cardiac tissue grown from pluripotent stem cells. Nature 556, 239–243 (2018).
Zhao, Y. et al. A platform for generation of chamber-specific cardiac tissues and disease modeling. Cell 176, 913–927.e18 (2019).
Mills, R. J. et al. Functional screening in human cardiac organoids reveals a metabolic mechanism for cardiomyocyte cell cycle arrest. Proc. Natl Acad. Sci. USA 114, E8372–E8381 (2017).
Nunes, S. S. et al. Biowire: a platform for maturation of human pluripotent stem cell–derived cardiomyocytes. Nat. Methods 10, 781–787 (2013).
Tandon, N. et al. Electrical stimulation systems for cardiac tissue engineering. Nat. Protoc. 4, 155–173 (2009).
Tandon, N. et al. Optimization of electrical stimulation parameters for cardiac tissue engineering. J. Tissue Eng. Regen. Med. 5, e115–e125 (2011).
Lasher, R. A., Pahnke, A. Q., Johnson, J. M., Sachse, F. B. & Hitchcock, R. W. Electrical stimulation directs engineered cardiac tissue to an age-matched native phenotype. J. Tissue Eng. 3, 204173141245535 (2012).
Marchianò, S., Bertero, A. & Murry, C. E. Learn from your elders: developmental biology lessons to guide maturation of stem cell-derived cardiomyocytes. Pediatr. Cardiol. 40, 1367–1387 (2019).
Breckenridge, R. A. et al. Hypoxic regulation of hand1 controls the fetal-neonatal switch in cardiac metabolism. PLoS Biol. 11, e1001666 (2013).
Neary, M. T. et al. Hypoxia signaling controls postnatal changes in cardiac mitochondrial morphology and function. J. Mol. Cell. Cardiol. 74, 340–352 (2014).
Menendez-Montes, I. et al. Myocardial VHL-HIF signaling controls an embryonic metabolic switch essential for cardiac maturation. Dev. Cell 39, 724–739 (2016).
Kreipke, R., Wang, Y., Miklas, J. W., Mathieu, J. & Ruohola-Baker, H. Metabolic remodeling in early development and cardiomyocyte maturation. Semin. Cell Dev. Biol. 52, 84–92 (2016).
Puente, B. N. et al. The oxygen-rich postnatal environment induces cardiomyocyte cell-cycle arrest through DNA damage response. Cell 157, 565–579 (2014).
Place, T. L., Domann, F. E. & Case, A. J. Limitations of oxygen delivery to cells in culture: an underappreciated problem in basic and translational research. Free Radic. Biol. Med. 113, 311–322 (2017).
Dunn, K. K. et al. Coculture of endothelial cells with human pluripotent stem cell-derived cardiac progenitors reveals a differentiation stage-specific enhancement of cardiomyocyte maturation. Biotechnol. J. 14, e1800725 (2019).
Sun, X. et al. Transplanted microvessels improve pluripotent stem cell–derived cardiomyocyte engraftment and cardiac function after infarction in rats. Sci. Transl. Med. 12, eaax2992 (2020).
Dong, H. W., Zhang, L. F. & Bao, S. L. AMPK regulates energy metabolism through the SIRT1 signaling pathway to improve myocardial hypertrophy. Eur. Rev. Med. Pharmacol. Sci. 22, 2757–2766 (2018).
Funakoshi, S. et al. Generation of mature compact ventricular cardiomyocytes from human pluripotent stem cells. Nat. Commun. 12, 3155 (2021).
Murphy, S. A. et al. PGC1/PPAR drive cardiomyocyte maturation at single cell level via YAP1 and SF3B2. Nat. Commun. 12, 1648 (2021).
Braidy, N. et al. Age related changes in NAD+ metabolism oxidative stress and sirt1 activity in wistar rats. PLoS One 6, e19194 (2011).
Stanley, W. C., Recchia, F. A. & Lopaschuk, G. D. Myocardial substrate metabolism in the normal and failing heart. Physiol. Rev. 85, 1093–1129 (2005).
Kuppusamy, K. T. et al. Let-7 family of microRNA is required for maturation and adult-like metabolism in stem cell-derived cardiomyocytes. Proc. Natl. Acad. Sci. 112, 201424042 (2015).
Yavari, A. et al. Mammalian γ2 AMPK regulates intrinsic heart rate. Nat. Commun. 8, 1258 (2017).
Wang, B. et al. AMPKα2 protects against the development of heart failure by enhancing mitophagy via PINK1 phosphorylation. Circ. Res. 122, 712–729 (2018).
Sakamoto, T. et al. A critical role for estrogen-related receptor signaling in cardiac maturation. Circ. Res. 126, 1685–1702 (2020).
Dufour, C. R. et al. Genome-wide orchestration of cardiac functions by the orphan nuclear receptors ERRα and γ. Cell Metab. 5, 345–356 (2007).
Xu, Y., Li, X., Liu, X. & Zhou, M. Neuregulin-1/ErbB signaling and chronic heart failure. Adv. Pharm. 59, 31–51 (2010).
Lee, D. S. et al. Defined microRNAs induce aspects of maturation in mouse and human embryonic-stem-cell-derived cardiomyocytes. Cell Rep. 12, 1960–1967 (2015).
Rupert, C. E. & Coulombe, K. L. K. IGF1 and NRG1 enhance proliferation, metabolic maturity, and the force-frequency response in hESC-derived engineered cardiac tissues. Stem Cells Int. 2017, 1–13 (2017).
Forhead, A. J. & Fowden, A. L. Thyroid hormones in fetal growth and prepartum maturation. J. Endocrinol. 221, R87–R103 (2014).
Chattergoon, N. N., Louey, S., Stork, P., Giraud, G. D. & Thornburg, K. L. Mid-gestation ovine cardiomyocytes are vulnerable to mitotic suppression by thyroid hormone. Reprod. Sci. 19, 642–649 (2012).
Chattergoon, N. N. et al. Thyroid hormone drives fetal cardiomyocyte maturation. FASEB J. 26, 397–408 (2012).
Correia, C. et al. Distinct carbon sources affect structural and functional maturation of cardiomyocytes derived from human pluripotent stem cells. Sci. Rep. 7, 8590 (2017).
Sebastião, M. J. et al. Bioreactor-based 3D human myocardial ischemia/reperfusion in vitro model: a novel tool to unveil key paracrine factors upon acute myocardial infarction. Transl. Res. 215, 57–74 (2020).
Huang, C. Y. et al. Enhancement of human iPSC-derived cardiomyocyte maturation by chemical conditioning in a 3D environment. J. Mol. Cell. Cardiol. 138, 1–11 (2020).
Feyen, D. A. M. et al. Metabolic maturation media improve physiological function of human iPSC-derived cardiomyocytes. Cell Rep. 32, 107925 (2020).
Parikh, S. S. et al. Thyroid and glucocorticoid hormones promote functional T-tubule development in human-induced pluripotent stem cell-derived cardiomyocytes. Circ. Res. 121, 1323–1330 (2017).
Rog-Zielinska, E. A. et al. Glucocorticoids promote structural and functional maturation of foetal cardiomyocytes: a role for PGC-1α. Cell Death Differ. 22, 1106–1116 (2015).
Hoover, L. L., Burton, E. G., Brooks, B. A. & Kubalak, S. W. The expanding role for retinoid signaling in heart development. Sci. World J. 8, 194–211 (2008).
Miao, S. et al. Retinoic acid promotes metabolic maturation of human embryonic stem cell-derived cardiomyocytes. Theranostics 10, 9686–9701 (2020).
Lee, J. H., Protze, S. I., Laksman, Z., Backx, P. H. & Keller, G. M. Human pluripotent stem cell-derived atrial and ventricular cardiomyocytes develop from distinct mesoderm populations. Cell Stem Cell 21, 179–194.e4 (2017).
Stoppel, W. L., Kaplan, D. L. & Black, L. D. Electrical and mechanical stimulation of cardiac cells and tissue constructs. Adv. Drug Deliv. Rev. 96, 135–155 (2016).
Boateng, S. Y. & Goldspink, P. H. Assembly and maintenance of the sarcomere night and day. Cardiovasc. Res. 77, 667–675 (2008).
van Eldik, W. & Passier, R. Signalling in sarcomeres in development and disease. Neth. Heart J. 21, 367–371 (2013).
Prill, K. & Dawson, J. F. Assembly and maintenance of sarcomere thin filaments and associated diseases. Int. J. Mol. Sci. 21, 1–20 (2020).
Coisy-Quivy, M. et al. TC10 controls human myofibril organization and is activated by the sarcomeric RhoGEF obscurin. J. Cell Sci. 122, 947–956 (2009).
Samarel, A. M., Koshman, Y., Swanson, E. R. & Russell, B. Biophysical Forces Modulate the Costamere and Z-Disc for Sarcomere Remodeling in Heart Failure. in Biophysics ofthe Failing Heart: Physics and Biology of Heart Muscle (eds. Solaro, R. J. & Tardiff, J. C.) 141–174 (2013). https://doi.org/10.1007/978-1-4614-7678-8_7.
Pyle, W. G. & Solaro, R. J. At the crossroads of myocardial signaling: the role of Z-discs in intracellular signaling and cardiac function. Circ. Res. 94, 296–305 (2004).
Nguyen, N. U. N. et al. A calcineurin-Hoxb13 axis regulates growth mode of mammalian cardiomyocytes. Nature 582, 271–276 (2020).
Rath, N., Wang, Z., Lu, M. M. & Morrisey, E. E. LMCD1/Dyxin is a novel transcriptional cofactor that restricts GATA6 function by inhibiting DNA binding. Mol. Cell. Biol. 25, 8864–8873 (2005).
Bian, Z.-Y. et al. LIM and cysteine-rich domains 1 regulates cardiac hypertrophy by targeting calcineurin/nuclear factor of activated T cells signaling. Hypertension 55, 257–263 (2010).
Pasqualini, F. S., Nesmith, A. P., Horton, R. E., Sheehy, S. P. & Parker, K. K. Mechanotransduction and metabolism in cardiomyocyte microdomains. Biomed. Res. Int. 2016, 4081638 (2016).
Kodde, I. F., van der Stok, J., Smolenski, R. T. & de Jong, J. W. Metabolic and genetic regulation of cardiac energy substrate preference. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 146, 26–39 (2007).
Taegtmeyer, H. et al. Assessing cardiac metabolism: a scientific statement from the American Heart Association. Circ. Res. 118, 1659–1701 (2016).
Martínez, M. S. et al. Energetic metabolism in cardiomyocytes: molecular basis of heart ischemia and arrhythmogenesis. Vessel 1, 130–141 (2017).
Pascual, F. & Coleman, R. A. Fuel availability and fate in cardiac metabolism: a tale of two substrates. Biochim. Biophys. Acta 1861, 1425–1433 (2016).
Bround, M. J. et al. Cardiomyocyte ATP production, metabolic flexibility, and survival require calcium flux through cardiac ryanodine receptors in vivo. J. Biol. Chem. 288, 18975–18986 (2013).
Yang, X. et al. Fatty acids enhance the maturation of cardiomyocytes derived from human pluripotent stem cells. Stem Cell Rep. 13, 657–668 (2019).
Hidalgo, A. et al. Modelling ischemia-reperfusion injury (IRI) in vitro using metabolically matured induced pluripotent stem cell-derived cardiomyocytes. APL Bioeng. 2, 026102 (2018).
Cardoso, A. C. et al. Mitochondrial substrate utilization regulates cardiomyocyte cell-cycle progression. Nat. Metab. 2, 167–178 (2020).
Nakano, H. et al. Glucose inhibits cardiac muscle maturation through nucleotide biosynthesis. Elife 6, e29330 (2017).
Skaper, S. D. & Facci, L. Central nervous system neuron-glia co-culture models and application to neuroprotective agents. In (ed. Skaper, S. D.) 63–80 (Springer New York, 2018). https://doi.org/10.1007/978-1-4939-7571-6_5.
Qian, X., Song, H. & Ming, G. Brain organoids: advances, applications and challenges. Development 146, dev166074 (2019).
Cakir, B. et al. Engineering of human brain organoids with a functional vascular-like system. Nat. Methods 16, 1169–1175 (2019).
Wörsdörfer, P. et al. Generation of complex human organoid models including vascular networks by incorporation of mesodermal progenitor cells. Sci. Rep. 9, 15663 (2019).
Mink, J. W., Blumenschine, R. J. & Adams, D. B. Ratio of central nervous system to body metabolism in vertebrates: its constancy and functional basis. Am. J. Physiol. Integr. Comp. Physiol. 241, R203–R212 (1981).
Bélanger, M., Allaman, I. & Magistretti, P. J. Brain energy metabolism: focus on astrocyte-neuron metabolic cooperation. Cell Metab. 14, 724–738 (2011).
Agostini, M. et al. Metabolic reprogramming during neuronal differentiation. Cell Death Differ. 23, 1502–1514 (2016).
Sonnay, S., Gruetter, R. & Duarte, J. M. N. How energy metabolism supports cerebral function: insights from 13c magnetic resonance studies in vivo. Front. Neurosci. 11, 288 (2017).
Jha, M. K. & Morrison, B. M. Glia-neuron energy metabolism in health and diseases: new insights into the role of nervous system metabolic transporters. Exp. Neurol. 309, 23–31 (2018).
Bazargani, N. & Attwell, D. Astrocyte calcium signaling: the third wave. Nat. Neurosci. 19, 182–189 (2016).
Agrawal, A., Pekkurnaz, G. & Koslover, E. F. Spatial control of neuronal metabolism through glucose-mediated mitochondrial transport regulation. Elife 7, e40986 (2018).
Kimura, I. et al. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. 108, 8030–8035 (2011).
Wang, L. et al. Brain development and Akt signaling: the crossroads of signaling pathway and neurodevelopmental diseases. J. Mol. Neurosci. 61, 379–384 (2017).
Sittewelle, M. & Monsoro-Burq, A. H. AKT signaling displays multifaceted functions in neural crest development. Dev. Biol. 444, S144–S155 (2018).
Roy, A. et al. PI3K-Yap activity drives cortical gyrification and hydrocephalus in mice. Elife 8, e45961 (2019).
Sánchez-Alegría, K., Flores-León, M., Avila-Muñoz, E., Rodríguez-Corona, N. & Arias, C. PI3K signaling in neurons: a central node for the control of multiple functions. Int. J. Mol. Sci. 19, 3725 (2018).
Marinangeli, C. et al. AMP-activated protein kinase is essential for the maintenance of energy levels during synaptic activation. iScience 9, 1–13 (2018).
Kleszka, K. et al. Hypoxia-inducible factor-2α is crucial for proper brain development. Sci. Rep. 10, 19146 (2020).
Domise, M. et al. Neuronal AMP-activated protein kinase hyper-activation induces synaptic loss by an autophagy-mediated process. Cell Death Dis. 10, 221 (2019).
Ardalani, H. et al. 3-D culture and endothelial cells improve maturity of human pluripotent stem cell-derived hepatocytes. Acta Biomater. 95, 371–381 (2019).
Mayati, A. et al. Functional polarization of human hepatoma HepaRG cells in response to forskolin. Sci. Rep. 8, 16115 (2018).
Porat-Shliom, N. et al. Liver kinase B1 regulates hepatocellular tight junction distribution and function in vivo. Hepatology 64, 1317–1329 (2016).
Zhang, B. et al. Biodegradable scaffold with built-in vasculature for organ-on-a-chip engineering and direct surgical anastomosis. Nat. Mater. 15, 669–678 (2016).
Peng, W. C., Kraaier, L. J. & Kluiver, T. A. Hepatocyte organoids and cell transplantation: what the future holds. Exp. Mol. Med. 53:1512–1528 (2021).
Chien, D., Dean, D., Saha, A. K., Flatt, J. P. & Ruderman, N. B. Malonyl-CoA content and fatty acid oxidation in rat muscle and liver in vivo. Am. J. Physiol. Metab. 279, E259–E265 (2000).
Boon, R. et al. Amino acid levels determine metabolism and CYP450 function of hepatocytes and hepatoma cell lines. Nat. Commun. 11, 1393 (2020).
Wang, K., Wang, M., Gannon, M. & Holterman, A. Growth hormone mediates its protective effect in hepatic apoptosis through Hnf6. PLoS One 11, e0167085 (2016).
Ogawa, S. et al. Three-dimensional culture and cAMP signaling promote the maturation of human pluripotent stem cell-derived hepatocytes. Development 140, 3285 LP–3283296 (2013).
Homolya, L. et al. LKB1/AMPK and PKA control ABCB11 trafficking and polarization in hepatocytes. PLoS One 9, e91921 (2014).
Fu, D., Wakabayashi, Y., Lippincott-Schwartz, J. & Arias, I. M. Bile acid stimulates hepatocyte polarization through a cAMP-Epac-MEK-LKB1-AMPK pathway. Proc. Natl Acad. Sci. 108, 1403–1408 (2011).
Fu, D., Wakabayashi, Y., Ido, Y., Lippincott-Schwartz, J. & Arias, I. M. Regulation of bile canalicular network formation and maintenance by AMP-activated protein kinase and LKB1. J. Cell Sci. 123, 3294–3302 (2010).
Jackson, L. N. et al. PI3K/Akt activation is critical for early hepatic regeneration after partial hepatectomy. Am. J. Physiol. Liver Physiol. 294, G1401–G1410 (2008).
Manning, B. D. & Toker, A. AKT/PKB signaling: navigating the network. Cell 169, 381–405 (2017).
Kamiya, A., Kinoshita, T. & Miyajima, A. Oncostatin M and hepatocyte growth factor induce hepatic maturation via distinct signaling pathways. FEBS Lett. 492, 90–94 (2001).
Cecchi, F., Rabe, D. C. & Bottaro, D. P. The hepatocyte growth factor receptor: structure, function and pharmacological targeting in cancer. Curr. Signal. Transduct. Ther. 6, 146–151 (2011).
Birchmeier, C., Birchmeier, W., Gherardi, E. & Vande Woude, G. F. Met, metastasis, motility and more. Nat. Rev. Mol. Cell Biol. 4, 915–925 (2003).
Timmermans, S., Souffriau, J. & Libert, C. A general introduction to glucocorticoid biology. Front. Immunol. 10, 1545 (2019).
Karagianni, P., Moulos, P., Schmidt, D., Odom, D. T. & Talianidis, I. Bookmarking by non-pioneer transcription factors during liver development establishes competence for future gene activation. Cell Rep. 30, 1319–1328.e6 (2020).
Qin, J. et al. Connexin 32-mediated cell-cell communication is essential for hepatic differentiation from human embryonic stem cells. Sci. Rep. 6, 37388 (2016).
Avior, Y. et al. Microbial-derived lithocholic acid and vitamin K 2 drive the metabolic maturation of pluripotent stem cells-derived and fetal hepatocytes. Hepatology 62, 265–278 (2015).
Sgodda, M. et al. A scalable approach for the generation of human pluripotent stem cell-derived hepatic organoids with sensitive hepatotoxicity features. Stem Cells Dev. 26, 1490–1504 (2017).
Camp, J. G. et al. Multilineage communication regulates human liver bud development from pluripotency. Nature 546, 533–538 (2017).
Kietzmann, T. Metabolic zonation of the liver: the oxygen gradient revisited. Redox Biol. 11, 622–630 (2017).
Blau, B. J. & Miki, T. The role of cellular interactions in the induction of hepatocyte polarity and functional maturation in stem cell-derived hepatic cells. Differentiation 106, 42–48 (2019).
Chen, C., Soto-Gutierrez, A., Baptista, P. M. & Spee, B. Biotechnology challenges to in vitro maturation of hepatic stem cells. Gastroenterology 154, 1258–1272 (2018).
Nie, Y.-Z. et al. Recapitulation of hepatitis B virus–host interactions in liver organoids from human induced pluripotent stem cells. EBioMedicine 35, 114–123 (2018).
Molinaro, A., Becattini, B. & Solinas, G. Insulin signaling and glucose metabolism in different hepatoma cell lines deviate from hepatocyte physiology toward a convergent aberrant phenotype. Sci. Rep. 10, 12031 (2020).
Wei, J. et al. Glucose and lipid metabolism screening models of hepatocyte spheroids after culture with injectable fiber fragments. J. Tissue Eng. Regen. Med. 14, 774–788 (2020).
Georgakopoulos, N. et al. Long-term expansion, genomic stability and in vivo safety of adult human pancreas organoids. BMC Dev. Biol. 20, 4 (2020).
Balak, J. R. A., Juksar, J., Carlotti, F., Lo Nigro, A. & de Koning, E. J. P. Organoids from the human fetal and adult pancreas. Curr. Diab. Rep. 19, 160 (2019).
Pagliuca, F. W. et al. Generation of functional human pancreatic β cells in vitro. Cell 159, 428–439 (2014).
Raum, J. C. et al. Tshz1 regulates pancreatic β-cell maturation. Diabetes 64, 2905–2914 (2015).
Sinagoga, K. L. et al. Distinct roles for the mTOR pathway in postnatal morphogenesis, maturation and function of pancreatic islets. Development 144, 2402 LP–2402414 (2017).
Kebede, M. A., Alquier, T., Latour, M. G. & Poitout, V. Lipid receptors and islet function: therapeutic implications? Diabetes, Obes. Metab. 11, 10–20 (2009).
Borden, P., Houtz, J., Leach, S. D. & Kuruvilla, R. Sympathetic innervation during development is necessary for pancreatic islet architecture and functional maturation. Cell Rep. 4, 287–301 (2013).
Ceasrine, A. M., Lin, E. E., Lumelsky, D. N., Iyer, R. & Kuruvilla, R. Adrb2 controls glucose homeostasis by developmental regulation of pancreatic islet vasculature. Elife 7, e39689 (2018).
Jacovetti, C., Matkovich, S. J., Rodriguez-Trejo, A., Guay, C. & Regazzi, R. Postnatal β-cell maturation is associated with islet-specific microRNA changes induced by nutrient shifts at weaning. Nat. Commun. 6, 8084 (2015).
Moullé, V. S. & Parnet, P. Effects of nutrient intake during pregnancy and lactation on the endocrine pancreas of the offspring. Nutrients 11, 2708 (2019).
Ni, Q. et al. Raptor regulates functional maturation of murine beta cells. Nat. Commun. 8, 15755 (2017).
Puri, S. et al. Replication confers β cell immaturity. Nat. Commun. 9, 485 (2018).
Yoshihara, E. et al. ERRγ is required for the metabolic maturation of therapeutically functional glucose-responsive β cells. Cell Metab. 23, 622–634 (2016).
Lien, Y.-C., Won, K.-J. & Simmons, R. A. Transcriptomic and quantitative proteomic profiling reveals signaling pathways critical for pancreatic islet maturation. Endocrinology 161, bqaa187 (2020).
Jaafar, R. et al. mTORC1-to-AMPK switching underlies β cell metabolic plasticity during maturation and diabetes. J. Clin. Invest. 129, 4124–4137 (2019).
Fu, A., Eberhard, C. E. & Screaton, R. A. Role of AMPK in pancreatic beta cell function. Mol. Cell. Endocrinol. 366, 127–134 (2013).
Vanderkruk, B. & Hoffman, B. G. Metabolism as a central regulator of β-cell chromatin state. FEBS J. 288, 3683–3693 (2020).
Ghazalli, N. et al. Glucocorticoid signaling enhances expression of glucose-sensing molecules in immature pancreatic beta-like cells derived from murine embryonic stem cells in vitro. Stem Cells Dev. 27, 898–909 (2018).
Chen, C., Xie, Z., Shen, Y. & Xia, S. F. The roles of thyroid and thyroid hormone in pancreas: physiology and pathology. Int. J. Endocrinol. 2018, 1–14 (2018).
Manhas, N. & Pardasani, K. R. Modelling mechanism of calcium oscillations in pancreatic acinar cells. J. Bioenerg. Biomembr. 46, 403–420 (2014).
González-Vélez, V., Piron, A. & Dupont, G. Calcium oscillations in pancreatic α-cells rely on noise and ATP-driven changes in membrane electrical activity. Front. Physiol. 11, 1515 (2020).
Fridlyand, L. E., Tamarina, N. & Philipson, L. H. Bursting and calcium oscillations in pancreatic β-cells: specific pacemakers for specific mechanisms. Am. J. Physiol. Metab. 299, E517–E532 (2010).
Watts, M. et al. Calcium and metabolic oscillations in pancreatic islets: who’s driving the bus? SIAM J. Appl. Dyn. Syst. 13, 683–703 (2014).
Chramiec, A. et al. Integrated human organ-on-a-chip model for predictive studies of anti-tumor drug efficacy and cardiac safety. Lab Chip 20, 4357–4372 (2020).
Zhang, Y. S. et al. Multisensor-integrated organs-on-chips platform for automated and continual in situ monitoring of organoid behaviors. Proc. Natl Acad. Sci. 114, E2293–E2302 (2017).
Maffioletti, S. M. et al. Three-dimensional human iPSC-derived artificial skeletal muscles model muscular dystrophies and enable multilineage tissue engineering. Cell Rep. 23, 899–908 (2018).
Moyle, L. A., Jacques, E. & Gilbert, P. M. Engineering the next generation of human skeletal muscle models: from cellular complexity to disease modeling. Curr. Opin. Biomed. Eng. 16, 9–18 (2020).
Gilbert‐Honick, J. & Grayson, W. Vascularized and innervated skeletal muscle tissue engineering. Adv. Healthc. Mater. 9, 1900626 (2020).
Reid, G., Magarotto, F., Marsano, A. & Pozzobon, M. Next stage approach to tissue engineering skeletal muscle. Bioengineering 7, 118 (2020).
Iotti, S., Borsari, M. & Bendahan, D. Oscillations in energy metabolism. Biochim. Biophys. Acta - Bioenerg. 1797, 1353–1361 (2010).
Friedrichsen, M., Mortensen, B., Pehmøller, C., Birk, J. B. & Wojtaszewski, J. F. P. Exercise-induced AMPK activity in skeletal muscle: Role in glucose uptake and insulin sensitivity. Mol. Cell. Endocrinol. 366, 204–214 (2013).
Khodabukus, A. & Baar, K. Glucose concentration and streptomycin alter in vitro muscle function and metabolism. J. Cell. Physiol. 230, 1226–1234 (2015).
Cheng, C. S., Ran, L., Bursac, N., Kraus, W. E. & Truskey, G. A. Cell density and joint microRNA-133a and microRNA-696 inhibition enhance differentiation and contractile function of engineered human skeletal muscle tissues. Tissue Eng. Part A 22, 573–583 (2016).
Schiaffino, S., Dyar, K. A., Ciciliot, S., Blaauw, B. & Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 280, 4294–4314 (2013).
You, J.-S. et al. The role of diacylglycerol kinase ζ and phosphatidic acid in the mechanical activation of mammalian target of rapamycin (mTOR) signaling and skeletal muscle hypertrophy. J. Biol. Chem. 289, 1551–1563 (2014).
Marcotte, G. R., West, D. W. D. & Baar, K. The molecular basis for load-induced skeletal muscle hypertrophy. Calcif. Tissue Int. 96, 196–210 (2015).
Blattler, S. M. et al. Defective mitochondrial morphology and bioenergetic function in mice lacking the transcription factor yin yang 1 in skeletal muscle. Mol. Cell. Biol. 32, 3333–3346 (2012).
Marabita, M. et al. S6K1 is required for increasing skeletal muscle force during hypertrophy. Cell Rep. 17, 501–513 (2016).
Khodabukus, A. et al. Electrical stimulation increases hypertrophy and metabolic flux in tissue-engineered human skeletal muscle. Biomaterials 198, 259–269 (2019).
Goodman, C. A., McNally, R. M., Hoffmann, F. M. & Hornberger, T. A. Smad3 Induces Atrogin-1, Inhibits mTOR and Protein Synthesis, and Promotes Muscle Atrophy In Vivo. Mol. Endocrinol. 27, 1946–1957 (2013).
Sparrow, J. C. & Schöck, F. The initial steps of myofibril assembly: integrins pave the way. Nat. Rev. Mol. Cell Biol. 10, 293–298 (2009).
Lemke, S. B. & Schnorrer, F. Mechanical forces during muscle development. Mech. Dev. 144, 92–101 (2017).
Ferreira, D. M. S. et al. LIM and cysteine-rich domains 1 (LMCD1) regulates skeletal muscle hypertrophy, calcium handling, and force. Skelet. Muscle 9, 26 (2019).
Shelton, C. D. & Byndloss, M. X. Gut epithelial metabolism as a key driver of intestinal dysbiosis associated with noncommunicable diseases. Infect. Immun. 88, e00939-19 (2020).
Snyder, J. et al. Materials and microenvironments for engineering the intestinal epithelium. Ann. Biomed. Eng. 48, 1916–1940 (2020).
Altay, G. et al. Self-organized intestinal epithelial monolayers in crypt and villus-like domains show effective barrier function. Sci. Rep. 9, 10140 (2019).
Beumer, J. & Clevers, H. Cell fate specification and differentiation in the adult mammalian intestine. Nat. Rev. Mol. Cell Biol. 22, 39–53 (2021).
Gattazzo, F., Urciuolo, A. & Bonaldo, P. Extracellular matrix: a dynamic microenvironment for stem cell niche. BBA - Gen. Subj. 1840, 2506–2519 (2014).
Blachier, F., Boutry, C., Bos, C. & Tomé, D. Metabolism and functions of l-glutamate in the epithelial cells of the small and large intestines. Am. J. Clin. Nutr. 90, 814S–821S (2009).
Beaumont, M. et al. Gut microbiota derived metabolites contribute to intestinal barrier maturation at the suckling-to-weaning transition. Gut Microbes 11, 1268–1286 (2020).
Wang, Q. et al. Energy metabolism in the intestinal crypt epithelial cells of piglets during the suckling period. Sci. Rep. 8, 12948 (2018).
Yang, H., Wang, X., Xiong, X. & Yin, Y. Energy metabolism in intestinal epithelial cells during maturation along the crypt-villus axis. Sci. Rep. 6, 31917 (2016).
Kumar, N. et al. A YY1-dependent increase in aerobic metabolism is indispensable for intestinal organogenesis. Development 143, 3711–3722 (2016).
Srivillibhuthur, M. et al. TFAM is required for maturation of the fetal and adult intestinal epithelium. Dev. Biol. 439, 92–101 (2018).
Jung, K. B. et al. Blockade of STAT3 causes severe in vitro and in vivo maturation defects in intestinal organoids derived from human embryonic stem cells. J. Clin. Med. 8, 976 (2019).
Jung, K. B. et al. Interleukin-2 induces the in vitro maturation of human pluripotent stem cell-derived intestinal organoids. Nat. Commun. 9, 3039 (2018).
Jackson, D. N. & Theiss, A. L. Gut bacteria signaling to mitochondria in intestinal inflammation and cancer. Gut Microbes 11, 285–304 (2020).
Allaire, J. M. et al. The intestinal epithelium: central coordinator of mucosal immunity. Trends Immunol. 39, 677–696 (2018).
Parada Venegas, D. et al. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 10, 277 (2019).
Macia, L. et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 6, 6734 (2015).
Newman, J. C. & Verdin, E. β-Hydroxybutyrate: a signaling metabolite. Annu. Rev. Nutr. 37, 51–76 (2017).
Zhao, Y. et al. GPR43 mediates microbiota metabolite SCFA regulation of antimicrobial peptide expression in intestinal epithelial cells via activation of mTOR and STAT3. Mucosal Immunol. 11, 752–762 (2018).
Kong, S., Zhang, Y. H. & Zhang, W. Regulation of intestinal epithelial cells properties and functions by amino acids. Biomed. Res. Int. 2018, 2819154 (2018).
Min, S., Kim, S. & Cho, S.-W. Gastrointestinal tract modeling using organoids engineered with cellular and microbiota niches. Exp. Mol. Med. 52, 227–237 (2020).
Blutt, S. E., Crawford, S. E., Ramani, S., Zou, W. Y. & Estes, M. K. Engineered human gastrointestinal cultures to study the microbiome and infectious diseases. Cell. Mol. Gastroenterol. Hepatol. 5, 241–251 (2018).
Verma, S., Senger, S., Cherayil, B. J. & Faherty, C. S. Spheres of influence: insights into salmonella pathogenesis from intestinal organoids. Microorganisms 8, 504 (2020).
Zhang, J. et al. Primary human colonic mucosal barrier crosstalk with super oxygen-sensitive faecalibacterium prausnitzii in continuous. Cult. Med 2, 74–98.e9 (2021).
Noel, G. et al. A primary human macrophage-enteroid co-culture model to investigate mucosal gut physiology and host-pathogen interactions. Sci. Rep. 7, 45270 (2017).
Jose, S. S. et al. Comparison of two human organoid models of lung and intestinal inflammation reveals Toll-like receptor signalling activation and monocyte recruitment. Clin. Transl. Immunol. 9, e1131 (2020).
Little, M. H. & Combes, A. N. Kidney organoids: accurate models or fortunate accidents. Genes Dev. 33, 1319–1345 (2019).
Bhargava, P. & Schnellmann, R. G. Mitochondrial energetics in the kidney. Nat. Rev. Nephrol. 13, 629–646 (2017).
Lyu, Z. et al. PPARγ maintains the metabolic heterogeneity and homeostasis of renal tubules. EBioMedicine 38, 178–190 (2018).
Ross, B. D., Espinal, J. & Silva, P. Glucose metabolism in renal tubular function. Kidney Int 29, 54–67 (1986).
Low, J. H. et al. Generation of human PSC-derived kidney organoids with patterned nephron segments and a de novo vascular network. Cell Stem Cell 25, 373–387.e9 (2019).
Zhao, J. et al. Genomic integration of ERRγ-HNF1β regulates renal bioenergetics and prevents chronic kidney disease. Proc. Natl Acad. Sci. 115, E4910 LP–E4914919 (2018).
Fretz, J. A. et al. Early B-cell factor 1 is an essential transcription factor for postnatal glomerular maturation. Kidney Int. 85, 1091–1102 (2014).
Pianezzi, E. et al. Role of somatic cell sources in the maturation degree of human induced pluripotent stem cell-derived cardiomyocytes. Biochim. Biophys. Acta 1867, 118538 (2020).
Khoo, T. S. et al. Retention of somatic memory associated with cell identity, age and metabolism in induced pluripotent stem (iPS) cells reprogramming. Stem Cell Rev. Rep. 16, 251–261 (2020).
Mebratu, Y. & Tesfaigzi, Y. How ERK1/2 activation controls cell proliferation and cell death: is subcellular localization the answer? Cell Cycle 8, 1168–1175 (2009).
Yu, J. S. L. & Cui, W. Proliferation, survival and metabolism: the role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development 143, 3050–3060 (2016).
Bonini, M. G. & Gantner, B. N. The multifaceted activities of AMPK in tumor progression-why the “one size fits all” definition does not fit at all? IUBMB Life 65, 889–896 (2013).
Dagon, Y., Mantzoros, C. S. & Kim, Y.-B. AMPK ↔ Sirt1: from a signaling network to a combination drug. Metabolism 65, 1692–1694 (2016).
Hawley, S. A. et al. Calmodulin-dependent protein kinase kinase-β is an alternative upstream kinase for AMP-activated protein kinase. Cell Metab. 2, 9–19 (2005).
Ewart, M.-A. & Kennedy, S. AMPK and vasculoprotection. Pharmacol. Ther. 131, 242–253 (2011).
Hawley, S. A. et al. Phosphorylation by Akt within the ST loop of AMPK-α1 down-regulates its activation in tumour cells. Biochem. J. 459, 275–287 (2014).
Fogarty, S. & Hardie, D. G. Development of protein kinase activators: AMPK as a target in metabolic disorders and cancer. Biochim. Biophys. Acta 1804, 581–591 (2010).
Jessen, N., Sundelin, E. I. O. & Møller, A. B. AMP kinase in exercise adaptation of skeletal muscle. Drug Discov. Today 19, 999–1002 (2014).
Hardie, D., Ross, F. & Hawley, S. AMP-activated protein kinase: a target for drugs both ancient and modern. Chem. Biol. 19, 1222–1236 (2012).
Gwinn, D. M. et al. AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol. Cell 30, 214–226 (2009).
Cantó, C. & Auwerx, J. PGC-1α, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipido. 20, 98–105 (2009).
Wu, S., Wu, Y., Wu, T. & Wei, Y. Role of AMPK-mediated adaptive responses in human cells with mitochondrial dysfunction to oxidative stress. Biochim. Biophys. Acta 1840, 1331–1344 (2014).
Price, N. L. et al. SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 15, 675–690 (2012).
Sun, C. et al. SIRT1 improves insulin sensitivity under insulin-resistant conditions by repressing PTP1B. Cell Metab. 6, 307–319 (2007).
Cerutti, R. et al. NAD+-dependent activation of Sirt1 corrects the phenotype in a mouse model of mitochondrial disease. Cell Metab. 19, 1042–1049 (2014).
Cantó, C. & Auwerx, J. Targeting Sirtuin 1 to improve metabolism: all you need is NAD +? Pharmacol. Rev. 64, 166–187 (2012).
Halestrap, A. P. & Wilson, M. C. The monocarboxylate transporter family-Role and regulation. IUBMB Life 64, 109–119 (2012).
Lan, F., Cacicedo, J. M., Ruderman, N. & Ido, Y. SIRT1 modulation of the acetylation status, cytosolic localization, and activity of LKB1. J. Biol. Chem. 283, 27628–27635 (2008).
Yamauchi, T. & Kadowaki, T. Adiponectin receptor as a key player in healthy longevity and obesity-related diseases. Cell Metab. 17, 185–196 (2013).
Green, M. F., Anderson, K. A. & Means, A. R. Characterization of the CaMKKβ–AMPK signaling complex. Cell. Signal. 23, 2005–2012 (2011).
Kovacic, S. et al. Akt activity negatively regulates phosphorylation of AMP-activated protein kinase in the heart. J. Biol. Chem. 278, 39422–39427 (2003).
Kazyken, D. et al. AMPK directly activates mTORC2 to promote cell survival during acute energetic stress. Sci. Signal. 12, eaav3249 (2019).
Han, F. et al. The critical role of AMPK in driving Akt activation under stress, tumorigenesis and drug resistance. Nat. Commun. 9, 4728 (2018).
Cherry, A. D. & Piantadosi, C. A. Regulation of mitochondrial biogenesis and its intersection with inflammatory responses. Antioxid. Redox Signal. 22, 965–976 (2015).
de la Cruz López, K. G., Toledo Guzmán, M. E., Sánchez, E. O. & García Carrancá, A. mTORC1 as a regulator of mitochondrial functions and a therapeutic target in cancer. Front. Oncol. 9, 1373 (2019).
Shaerzadeh, F., Motamedi, F. & Khodagholi, F. Inhibition of Akt phosphorylation diminishes mitochondrial biogenesis regulators, tricarboxylic acid cycle activity and exacerbates recognition memory deficit in rat model of Alzheimer’s disease. Cell. Mol. Neurobiol. 34, 1223–1233 (2014).
Goo, C. K. et al. PTEN/Akt signaling controls mitochondrial respiratory capacity through 4E-BP1. PLoS One 7, e45806 (2012).
Imamura, H. et al. Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encoded indicators. Proc. Natl Acad. Sci. 106, 15651–15656 (2009).
Baker, J. S., McCormick, M. C. & Robergs, R. A. Interaction among skeletal muscle metabolic energy systems during intense exercise. J. Nutr. Metab. 2010, 905612 (2010).
Zala, D. et al. The advantage of channeling nucleotides for very processive functions. F1000Research 6, 724 (2017).
Arkun, Y. & Yasemi, M. Dynamics and control of the ERK signaling pathway: sensitivity, bistability, and oscillations. PLoS One 13, e0195513 (2018).
Shankaran, H. et al. Rapid and sustained nuclear–cytoplasmic ERK oscillations induced by epidermal growth factor. Mol. Syst. Biol. 5, 332 (2009).
McNeil, L. K., Starr, T. K. & Hogquist, K. A. A requirement for sustained ERK signaling during thymocyte positive selection in vivo. Proc. Natl Acad. Sci. USA102, 13574 LP–13513579 (2005).
Liu, P., Kevrekidis, I. G. & Shvartsman, S. Y. Substrate-dependent control of ERK phosphorylation can lead to oscillations. Biophys. J. 101, 2572–2581 (2011).
Cooling, M. T., Hunter, P. & Crampin, E. J. Sensitivity of NFAT cycling to cytosolic calcium concentration: implications for hypertrophic signals in cardiac myocytes. Biophys. J. 96, 2095–2104 (2009).
Zhu, L. et al. Ca2+ oscillation frequency regulates agonist-stimulated gene expression in vascular endothelial cells. J. Cell Sci. 121, 2511–2518 (2008).
Dupont, G., Combettes, L., Bird, G. S. & Putney, J. W. Calcium oscillations. Cold Spring Harb. Perspect. Biol. 3, a004226–a004226 (2011).
Huang, K.-P. & Huang, F. L. How is protein kinase C activated in CNS. Neurochem. Int. 22, 417–433 (1993).
Mogami, H. et al. Decoding of short-lived Ca2+ influx signals into long term substrate phosphorylation through activation of two distinct classes of protein kinase C. J. Biol. Chem. 278, 9896–9904 (2003).
Siso-Nadal, F., Fox, J. J., Laporte, S. A., Hébert, T. E. & Swain, P. S. Cross-talk between signaling pathways can generate robust oscillations in calcium and cAMP. PLoS One 4, e7189 (2009).
Roux, E., Bougaran, P., Dufourcq, P. & Couffinhal, T. Fluid shear stress sensing by the endothelial layer. Front. Physiol. 11, 861 (2020).
Reneman, R. S., Arts, T. & Hoeks, A. P. G. Wall shear stress – an important determinant of endothelial cell function and structure – in the arterial system in vivo. J. Vasc. Res. 43, 251–269 (2006).
Wong, A. Y. K. & Klassen, G. A. A model of electrical activity and cytosolic calcium dynamics in vascular endothelial cells in response to fluid shear stress. Ann. Biomed. Eng. 23, 822–832 (1995).
Tsoukias, N. M. Calcium dynamics and signaling in vascular regulation: computational models. Wiley Interdiscip. Rev. Syst. Biol. Med. 3, 93–106 (2011).
Yokota, Y. et al. Endothelial Ca2+ oscillations reflect VEGFR signaling-regulated angiogenic capacity in vivo. Elife 4, e08817 (2015).
Sato, D., Dixon, R. E., Santana, L. F. & Navedo, M. F. A model for cooperative gating of L-type Ca2+ channels and its effects on cardiac alternans dynamics. PLOS Comput. Biol. 14, e1005906 (2018).
Matveev, V., Bertram, R. & Sherman, A. Ca2+ current versus Ca2+ channel cooperativity of exocytosis. J. Neurosci. 29, 12196–12209 (2009).
Schneider, R. et al. Mobility of calcium channels in the presynaptic membrane. Neuron 86, 672–679 (2015).
Hancock, C. R., Janssen, E. & Terjung, R. L. Contraction-mediated phosphorylation of AMPK is lower in skeletal muscle of adenylate kinase-deficient mice. J. Appl. Physiol. 100, 406–413 (2006).
Richter, E. A. & Ruderman, N. B. AMPK and the biochemistry of exercise: implications for human health and disease. Biochem. J. 418, 261–275 (2009).
Merrins, M. J. et al. Metabolic oscillations in pancreatic islets depend on the intracellular Ca2+ level but not Ca2+ oscillations. Biophys. J. 99, 76–84 (2010).
Gilon, P., Ravier, M. A., Jonas, J.-C. & Henquin, J.-C. Control mechanisms of the oscillations of insulin secretion in vitro and in vivo. Diabetes 51, S144–S151 (2002).
Du Pré, B. C. et al. Circadian rhythms in cell maturation. Physiology 29, 72–83 (2014).
Dierickx, P. et al. Circadian Rhythms in Stem Cell Biology and Function. in Stem Cells and Cardiac Regeneration (ed. Madonna, R.) 57–78, https://doi.org/10.1007/978-3-319-25427-2_5 (Springer International Publishing, 2016).
Rakshit, K. et al. Postnatal ontogenesis of the islet circadian clock plays a contributory role in β-cell maturation process. Diabetes 67, 911 LP–911922 (2018).
Vining, K. H. & Mooney, D. J. Mechanical forces direct stem cell behaviour in development and regeneration. Nat. Rev. Mol. Cell Biol. 18, 728–742 (2017).
Schwartz, M. A. & Assoian, R. K. Integrins and cell proliferation: regulation of cyclin-dependent kinases via cytoplasmic signaling pathways. J. Cell Sci. 114, 2553–2560 (2001).
Carlier, A. & Alsberg, E. Harnessing topographical cues for tissue engineering. Tissue Eng. Part A 22, 995–996 (2016).
Li, L., Eyckmans, J. & Chen, C. S. Designer biomaterials for mechanobiology. Nat. Mater. 16, 1164–1168 (2017).
Romani, P., Valcarcel-Jimenez, L., Frezza, C. & Dupont, S. Crosstalk between mechanotransduction and metabolism. Nat. Rev. Mol. Cell Biol. 22, 22–38 (2020).
Farge, E. Chapter eight - Mechanotransduction in Development. in Forces and Tension in Development (ed. Labouesse, M. B. T.-C. T. in D. B.) vol. 95 243–265 (Academic Press, 2011).
Wozniak, M. A. & Chen, C. S. Mechanotransduction in development: a growing role for contractility. Nat. Rev. Mol. Cell Biol. 10, 34–43 (2009).
Martino, F., Perestrelo, A. R., Vinarský, V., Pagliari, S. & Forte, G. Cellular mechanotransduction: from tension to function. Front. Physiol. 9, 824 (2018).
Kaarj, K. & Yoon, J.-Y. Methods of delivering mechanical stimuli to organ-on-a-chip. Micromachines 10, 700 (2019).
Sun, Z., Guo, S. S. & Fässler, R. Integrin-mediated mechanotransduction. J. Cell Biol. 215, 445–456 (2016).
Vethe, H. et al. The effect of Wnt pathway modulators on human iPSC-derived pancreatic beta cell maturation. Front. Endocrinol. 10, 293 (2019).
Perugorria, M. J. et al. Wnt–β-catenin signalling in liver development, health and disease. Nat. Rev. Gastroenterol. Hepatol. 16, 121–136 (2019).
Yamaguchi, T., Nishijima, M., Tashiro, K. & Kawabata, K. Wnt- β -catenin signaling promotes the maturation of mast cells. Biomed. Res. Int. 2016, 1–8 (2016).
Clevers, H. Wnt/β-catenin signaling in development and disease. Cell 127, 469–480 (2006).
Klezovitch, O. & Vasioukhin, V. Cadherin signaling: keeping cells in touch. F1000Research 4, 550 (2015).
Halbleib, J. M. & Nelson, W. J. Cadherins in development: cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 20, 3199–3214 (2006).
Armeanu, S., Bühring, H. J., Reuss-Borst, M., Müller, C. A. & Klein, G. E-cadherin is functionally involved in the maturation of the erythroid lineage. J. Cell Biol. 131, 243–249 (1995).
Palmer, A. & Klein, R. Multiple roles of ephrins in morphogenesis, neuronal networking, and brain function. Genes Dev. 17, 1429–1450 (2003).
De Santis, G., Miotti, S., Mazzi, M., Canevari, S. & Tomassetti, A. E-cadherin directly contributes to PI3K/AKT activation by engaging the PI3K-p85 regulatory subunit to adherens junctions of ovarian carcinoma cells. Oncogene 28, 1206–1217 (2009).
Barber, A. G. et al. PI3K/AKT pathway regulates E-cadherin and Desmoglein 2 in aggressive prostate cancer. Cancer Med. 4, 1258–1271 (2015).
Li, P. et al. N-cadherin-mediated activation of PI3K/Akt-GSK-3β signaling attenuates nucleus pulposus cell apoptosis under high-magnitude compression. Cell. Physiol. Biochem. 44, 229–239 (2017).
Kumar, A., Placone, J. K. & Engler, A. J. Understanding the extracellular forces that determine cell fate and maintenance. Development 144, 4261 LP–4264270 (2017).
Mertz, A. F. et al. Cadherin-based intercellular adhesions organize epithelial cell-matrix traction forces. Proc. Natl. Acad. Sci. 110, 842–847 (2013).
Yim, E. K. F., Darling, E. M., Kulangara, K., Guilak, F. & Leong, K. W. Nanotopography-induced changes in focal adhesions, cytoskeletal organization, and mechanical properties of human mesenchymal stem cells. Biomaterials 31, 1299–1306 (2010).
Dalby, M. J., Gadegaard, N. & Oreffo, R. O. C. Harnessing nanotopography and integrin-matrix interactions to influence stem cell fate. Nat. Mater. 13, 558–569 (2014).
Abagnale, G. et al. Surface topography enhances differentiation of mesenchymal stem cells towards osteogenic and adipogenic lineages. Biomaterials 61, 316–326 (2015).
Higuchi, A. et al. Polymeric design of cell culture materials that guide the differentiation of human pluripotent stem cells. Prog. Polym. Sci. 65, 83–126 (2017).
Bonakdar, S. et al. Cell-imprinted substrates modulate differentiation, redifferentiation, and transdifferentiation. ACS Appl. Mater. Interfaces 8, 13777–13784 (2016).
Yang, C., Tibbitt, M. W., Basta, L. & Anseth, K. S. Mechanical memory and dosing influence stem cell fate. Nat. Mater. 13, 645–652 (2014).
Ireland, R. G. & Simmons, C. A. Human pluripotent stem cell mechanobiology: manipulating the biophysical microenvironment for regenerative medicine and tissue engineering applications. Stem Cells 33, 3187–3196 (2015).
Joo, S. et al. Effects of ECM protein micropatterns on the migration and differentiation of adult neural stem cells. Sci. Rep. 5, 13043 (2015).
Ventre, M., Natale, C. F., Rianna, C. & Netti, P. A. Topographic cell instructive patterns to control cell adhesion, polarization and migration. J. R. Soc. Interface 11, 20140687 (2014).
Luo, J. et al. The influence of nanotopography on cell behaviour through interactions with the extracellular matrix – a review. Bioact. Mater. 15, 145–159 (2022).
Meran, L., Baulies, A. & Li, V. S. W. Intestinal stem cell niche: the extracellular matrix and cellular components. Stem Cells Int. 2017, 7970385 (2017).
Wang, Y. et al. Bioengineered systems and designer matrices that recapitulate the intestinal stem cell niche. Cell. Mol. Gastroenterol. Hepatol. 5, 440–453.e1 (2018).
Gagné, D. et al. Integrin-linked kinase regulates migration and proliferation of human intestinal cells under a fibronectin-dependent mechanism. J. Cell. Physiol. 222, 387–400 (2010).
Kim, S., Uroz, M., Bays, J. L. & Chen, C. S. Harnessing mechanobiology for tissue engineering. Dev. Cell 56, 180–191 (2021).
Yuan, S. & Rigor, R. Chapter 5: Signaling Mechanisms in the Regulation of Endothelial Permeability. in Regulation of Endothelial Barrier Function (Morgan & Claypool Life Sciences; 2010).
Vion, A.-C. et al. Endothelial cell orientation and polarity are controlled by shear stress and VEGF through distinct signaling pathways. Front. Physiol. 11, 623769 (2021).
Bakhchova, L. et al. On-a-chip-based sensitive detection of drug-induced apoptosis in polarized gastric epithelial cells. ACS Biomater. Sci. Eng. 7, 5474–5483 (2021).
Bein, A. et al. Microfluidic organ-on-a-chip models of human intestine. Cell. Mol. Gastroenterol. Hepatol. 5, 659–668 (2018).
Deng, J. et al. Engineered liver-on-a-chip platform to mimic liver functions and its biomedical applications: a review. Micromachines 10, 676 (2019).
Smith, L. R., Cho, S. & Discher, D. E. Stem cell differentiation is regulated by extracellular matrix mechanics. Physiology 33, 16–25 (2018).
Wang, E. Y. et al. Biowire model of interstitial and focal cardiac fibrosis. ACS Cent. Sci. 5, 1146–1158 (2019).
Rodansky, E. S., Johnson, L. A., Huang, S., Spence, J. R. & Higgins, P. D. R. Intestinal organoids: a model of intestinal fibrosis for evaluating anti-fibrotic drugs. Exp. Mol. Pathol. 98, 346–351 (2015).
Tan, Q. et al. Nascent lung organoids reveal epithelium- and bone morphogenetic protein–mediated suppression of fibroblast activation. Am. J. Respir. Cell Mol. Biol. 61, 607–619 (2019).
Lee, J., Kim, J.-H., Hong, S.-H. & Yang, S.-R. Organoid model in idiopathic pulmonary fibrosis. Int. J. Stem Cells 14, 1–8 (2021).
Beech, D. J. & Kalli, A. C. Force sensing by piezo channels in cardiovascular health and disease. Arterioscler. Thromb. Vasc. Biol. 39, 2228–2239 (2019).
Douguet, D. & Honoré, E. Mammalian mechanoelectrical transduction: structure and function of force-gated ion channels. Cell 179, 340–354 (2019).
Murthy, S. E., Dubin, A. E. & Patapoutian, A. Piezos thrive under pressure: mechanically activated ion channels in health and disease. Nat. Rev. Mol. Cell Biol. 18, 771–783 (2017).
Lee, W. et al. Synergy between Piezo1 and Piezo2 channels confers high-strain mechanosensitivity to articular cartilage. Proc. Natl Acad. Sci. 111, E5114–E5122 (2014).
Sarre, A., Gabrielli, J., Vial, G., Leverve, X. M. & Assimacopoulos-Jeannet, F. Reactive oxygen species are produced at low glucose and contribute to the activation of AMPK in insulin-secreting cells. Free Radic. Biol. Med. 52, 142–150 (2012).
Low, L. A., Mummery, C., Berridge, B. R., Austin, C. P. & Tagle, D. A. Organs-on-chips: into the next decade. Nat. Rev. Drug Discov. 20, 345–361 (2020).
Laird, D. W. Life cycle of connexins in health and disease. Biochem. J. 394, 527–543 (2006).
Figueroa, X. F., Lillo, M. A., Gaete, P. S., Riquelme, M. A. & Sáez, J. C. Diffusion of nitric oxide across cell membranes of the vascular wall requires specific connexin-based channels. Neuropharmacology 75, 471–478 (2013).
Sun, J., Hoying, J. B., Deymier, P. A., Zhang, D. D. & Wong, P. K. Cellular architecture regulates collective calcium signaling and cell contractility. PLOS Comput. Biol. 12, e1004955 (2016).
Aratyn-Schaus, Y. et al. Coupling primary and stem cell–derived cardiomyocytes in an in vitro model of cardiac cell therapy. J. Cell Biol. 212, 389–397 (2016).
Spyrou, J., Gardner, D. K. & Harvey, A. J. Metabolomic and transcriptional analyses reveal atmospheric oxygen during human induced pluripotent stem cell generation impairs metabolic reprogramming. Stem Cells 37, 1042–1056 (2019).
Mullur, R., Liu, Y.-Y. & Brent, G. A. Thyroid hormone regulation of metabolism. Physiol. Rev. 94, 355–382 (2014).
Müller, J. & Heuer, H. Expression pattern of thyroid hormone transporters in the postnatal mouse brain. Front. Endocrinol. (Lausanne). 5, 92 (2014).
Groeneweg, S., van Geest, F. S., Peeters, R. P., Heuer, H. & Visser, W. E. Thyroid hormone transporters. Endocr. Rev. 41, 146–201 (2020).
Hsu, J. H., Zavacki, A. M., Harney, J. W. & Brent, G. A. Retinoid-X receptor (RXR) differentially augments thyroid hormone response in cell lines as a function of the response element and endogenous RXR content. Endocrinology 136, 421–430 (1995).
Tripathi, M., Yen, P. M. & Singh, B. K. Estrogen-related receptor alpha: an under-appreciated potential target for the treatment of metabolic diseases. Int. J. Mol. Sci. 21, 1645 (2020).
Bougarne, N. et al. PPAR blocks glucocorticoid receptor -mediated transactivation but cooperates with the activated glucocorticoid receptor for transrepression on NF- B. Proc. Natl Acad. Sci. 106, 7397–7402 (2009).
Ferris, H. A. & Kahn, C. R. New mechanisms of glucocorticoid-induced insulin resistance: make no bones about it. J. Clin. Invest. 122, 3854–3857 (2012).
Harmon, G. S., Lam, M. T. & Glass, C. K. PPARs and lipid ligands in inflammation and metabolism. Chem. Rev. 111, 6321–6340 (2011).
Wang, Y.-X. PPARs: diverse regulators in energy metabolism and metabolic diseases. Cell Res. 20, 124–137 (2010).
Wei, W. et al. Ligand activation of ERRα by Cholesterol Mediates Statin And Bisphosphonate Effects. Cell Metab. 23, 479–491 (2016).
Audet-Walsh, É. & Giguére, V. The multiple universes of estrogen-related receptor α and γ in metabolic control and related diseases. Acta Pharmacol. Sin. 36, 51–61 (2015).
Pei, L. et al. Dependence of hippocampal function on ERRγ-regulated mitochondrial metabolism. Cell Metab. 21, 628–636 (2015).
Farese, R. V. & Sajan, M. P. Metabolic functions of atypical protein kinase C: “good” and “bad” as defined by nutritional status. Am. J. Physiol. Metab. 298, E385–E394 (2010).
Turban, S. & Hajduch, E. Protein kinase C isoforms: mediators of reactive lipid metabolites in the development of insulin resistance. FEBS Lett. 585, 269–274 (2011).
Schmitz-Peiffer, C. The tail wagging the dog - regulation of lipid metabolism by protein kinase C. FEBS J. 280, 5371–5383 (2013).
Harrison, D. A. The JAK/STAT pathway. Cold Spring Harb. Perspect. Biol. 4, a011205–a011205 (2012).
Speirs, C., Williams, J. J. L., Riches, K., Salt, I. P. & Palmer, T. M. Linking energy sensing to suppression of JAK-STAT signalling: A potential route for repurposing AMPK activators? Pharmacol. Res. 128, 88–100 (2018).
Rutherford, C. et al. Phosphorylation of Janus kinase 1 (JAK1) by AMP-activated protein kinase (AMPK) links energy sensing to anti-inflammatory signaling. Sci. Signal. 9, ra109–ra109 (2016).
Ferretti, A. C. et al. AMPK and PKA interaction in the regulation of survival of liver cancer cells subjected to glucose starvation. Oncotarget 7, 17815–17828 (2016).
Tan, H. W. S., Sim, A. Y. L. & Long, Y. C. Glutamine metabolism regulates autophagy-dependent mTORC1 reactivation during amino acid starvation. Nat. Commun. 8, 338 (2017).
Jing, H. & Lin, H. Sirtuins in epigenetic regulation. Chem. Rev. 115, 2350–2375 (2015).
Miyamoto, J. et al. Ketone body receptor GPR43 regulates lipid metabolism under ketogenic conditions. Proc. Natl. Acad. Sci. 116, 23813–23821 (2019).
Cheng, C.-W. et al. Ketone body signaling mediates intestinal stem cell homeostasis and adaptation to diet. Cell 178, 1115–1131.e15 (2019).
Shimazu, T. et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science 339, 211 LP–211214 (2013).
Cotter, D. G., Schugar, R. C. & Crawford, P. A. Ketone body metabolism and cardiovascular disease. Am. J. Physiol. Circ. Physiol. 304, H1060–H1076 (2013).
Chai, J. T., Digby, J. E. & Choudhury, R. P. GPR109A and vascular inflammation. Curr. Atheroscler. Rep. 15, 325 (2013).
Danielsen, E. T., Moeller, M. E. & Rewitz, K. F. Nutrient signaling and developmental timing of maturation. Curr. Top. Developmental Biol. 105, 37–67 (2013).
Khodabukus, A. & Baar, K. The effect of serum origin on tissue engineered skeletal muscle function. J. Cell. Biochem. 115, 2198–2207 (2014).
Arigony, A. L. V. et al. The influence of micronutrients in cell culture: a reflection on viability and genomic stability. Biomed. Res. Int. 2013, 597282 (2013).
Koning, M., van den Berg, C. W. & Rabelink, T. J. Stem cell-derived kidney organoids: engineering the vasculature. Cell. Mol. Life Sci. 77, 2257–2273 (2020).
Devall, M. et al. Modeling the effect of prolonged ethanol exposure on global gene expression and chromatin accessibility in normal 3D colon organoids. PLoS One 15, e0227116 (2020).
Kim, H. et al. Modeling G2019S-LRRK2 sporadic parkinson’s disease in 3d midbrain organoids. Stem Cell Rep. 12, 518–531 (2019).
Karnieli, O. et al. A consensus introduction to serum replacements and serum-free media for cellular therapies. Cytotherapy 19, 155–169 (2017).
McKee, T. J. & Komarova, S. V. Is it time to reinvent basic cell culture medium? Am. J. Physiol. Physiol. 312, C624–C626 (2017).
Serlin, Y., Shelef, I., Knyazer, B. & Friedman, A. Anatomy and physiology of the blood–brain barrier. Semin. Cell Dev. Biol. 38, 2–6 (2015).
Bansode, S. et al. Glycation changes molecular organization and charge distribution in type I collagen fibrils. Sci. Rep. 10, 3397 (2020).
Chung, S. et al. Modulating cell culture oxidative stress reduces protein glycation and acidic charge variant formation. MAbs 11, 205–216 (2019).
Patel, S. H. et al. Advanced glycation end-products suppress mitochondrial function and proliferative capacity of achilles tendon-derived fibroblasts. Sci. Rep. 9, 12614 (2019).
Alsabeeh, N., Chausse, B., Kakimoto, P. A., Kowaltowski, A. J. & Shirihai, O. Cell culture models of fatty acid overload: Problems and solutions. Biochim. Biophys. Acta 1863, 143–151 (2018).
Cho, H. et al. Signaling dynamics of palmitate-induced ER stress responses mediated by ATF4 in HepG2 cells. BMC Syst. Biol. 7, 9 (2013).
Drenjančević, I., Jukić, I., Mihaljević, Z., Ćosić, A. & Kibel, A. The metabolites of arachidonic acid in microvascular function. In Microcirculation Revisited - From Molecules to Clinical Practice (ed. Jukić, I.) Ch. 5, https://doi.org/10.5772/64676 (InTech, 2016).
Smith, A. N., Muffley, L. A., Bell, A. N., Numhom, S. & Hocking, A. M. Unsaturated fatty acids induce mesenchymal stem cells to increase secretion of angiogenic mediators. J. Cell. Physiol. 227, 3225–3233 (2012).
Islam, K. et al. Co-culture methods used to model atherosclerosis in vitro using endothelial, smooth muscle and monocyte cells. SM J. Biomed. Eng. 2, 1008 (2016).
McBane, J. E., Matheson, L. A., Sharifpoor, S., Santerre, J. P. & Labow, R. S. Effect of polyurethane chemistry and protein coating on monocyte differentiation towards a wound healing phenotype macrophage. Biomaterials 30, 5497–5504 (2009).
Lesher-Pérez, S. C. et al. Dispersible oxygen microsensors map oxygen gradients in three-dimensional cell cultures. Biomater. Sci. 5, 2106–2113 (2017).
Moraes, C. et al. Integrating polyurethane culture substrates into poly(dimethylsiloxane) microdevices. Biomaterials 30, 5241–5250 (2009).
Talman, V. & Kivelä, R. Cardiomyocyte—endothelial cell interactions in cardiac remodeling and regeneration. Front. Cardiovasc. Med. 5, 1–8 (2018).
Koh, M. Y. & Powis, G. Passing the baton: the HIF switch. Trends Biochem. Sci. 37, 364–372 (2012).
Tafani, M. et al. The interplay of reactive oxygen species, hypoxia, inflammation, and sirtuins in cancer initiation and progression. Oxid. Med. Cell. Longev. 2016, 3907147 (2016).
Clanton, T. L. Hypoxia-induced reactive oxygen species formation in skeletal muscle. J. Appl. Physiol. 102, 2379–2388 (2007).
Misra, H. P. & Fridovich, I. The univalent reduction of oxygen by reduced flavins and quinones. J. Biol. Chem. 247, 188–192 (1972).
Schieber, M. & Chandel, N. S. ROS function in redox signaling and oxidative stress. Curr. Biol. 24, R453–R462 (2014).
Thannickal, V. J. & Fanburg, B. L. Reactive oxygen species in cell signaling. Am. J. Physiol. Cell. Mol. Physiol. 279, L1005–L1028 (2000).
Patel, R., Rinker, L., Peng, J. & Chilian, W. M. Reactive oxygen species: the good and the bad. in Reactive Oxygen Species (ROS) in Living Cells, https://doi.org/10.5772/intechopen.71547 (2018).
Cheong, R. & Levchenko, A. Oscillatory signaling processes: the how, the why and the where. Curr. Opin. Genet. Dev. 20, 665–669 (2010).
Ghosh, D. et al. Dynamic L-type CaV1.2 channel trafficking facilitates CaV1.2 clustering and cooperative gating. Biochim. Biophys. Acta 1865, 1341–1355 (2018).
Peyronnet, R., Nerbonne, J. M. & Kohl, P. Cardiac mechano-gated ion channels and arrhythmias. Circ. Res. 118, 311–329 (2016).
Toepfer, C. N., West, T. G. & Ferenczi, M. A. Revisiting Frank-Starling: regulatory light chain phosphorylation alters the rate of force redevelopment (k tr) in a length-dependent fashion. J. Physiol. 594, 5237–5254 (2016).
Sequeira, V. & van der Velden, J. Historical perspective on heart function: the Frank–Starling law. Biophys. Rev. 7, 421–447 (2015).
Kaur, S., Shen, X., Power, A. & Ward, M.-L. Stretch modulation of cardiac contractility: importance of myocyte calcium during the slow force response. Biophys. Rev. 12, 135–142 (2020).
Johnson, D. M. & Antoons, G. Arrhythmogenic mechanisms in heart failure: linking β-adrenergic stimulation, stretch, and calcium. Front. Physiol. 9, 1–23 (2018).
Dowrick, J. M. et al. The slow force response to stretch: controversy and contradictions. Acta Physiol. 226, e13250 (2019).
Park, S. H. & Park, J. O. Simultaneous Optimization of Multiple Responses Using a Weighted Desirability Function. in Quality Improvement Through Statistical Methods (ed. Abraham, B.) 299–311, https://doi.org/10.1007/978-1-4612-1776-3_24 (Birkhäuser Boston, 1998).
Wismans, L. J. J., Brands, T., Van Berkum, E. C. & Bliemer, M. C. J. Pruning and ranking the Pareto optimal set, application for the dynamic multi-objective network design problem. J. Adv. Transp. 48, 588–607 (2014).
Ehtamo, H. et al. Generating pareto solutions in a two-party setting: constraint proposal methods. Manag. Sci. 45, 1697–1709 (1999).
Tappeta, R. V. & Renaud, J. E. Interactive multiobjective optimization design strategy for decision based design. J. Mech. Des. 123, 205–215 (2001).
Kim, M. M. & Audet, J. On-demand serum-free media formulations for human hematopoietic cell expansion using a high dimensional search algorithm. Commun. Biol. 2, 1–11 (2019).
Garai, A., Mandal, P. & Roy, T. K. Iterative solution process for multiple objective stochastic linear programming problems under fuzzy environment. Fuzzy Inf. Eng. 0, 1–17 (2020).
Cho, G.-S. et al. Neonatal transplantation confers maturation of PSC-derived cardiomyocytes conducive to modeling cardiomyopathy. Cell Rep. 18, 571–582 (2017).
Mueller, C. et al. Effects of external stimulators on engineered skeletal muscle tissue maturation. Adv. Mater. Interfaces 8, 2001167 (2021).
Wang, J. et al. Engineered skeletal muscles for disease modeling and drug discovery. Biomaterials 221, 119416 (2019).
Acknowledgements
This research is supported by the Dalhousie Medical Research Foundation-Faculty of Dentistry Early Career Researcher Award to L.D.H. N.I.C. and R.G.I. were funded by Vanier Canada Graduate Scholarships, from the Natural Sciences and Engineering Research Council of Canada (NSERC) and the Canadian Institutes of Health Research (CIHR), respectively. L.J.D. was funded by the Translational Biology and Engineering Program Ted Rogers Center for Heart Research.
Author information
Authors and Affiliations
Contributions
N.I.C. conceived the manuscript. N.I.C., L.J.D., and R.G.I. wrote the manuscript. N.I.C., L.J.D., J.P.S., C.A.S., and L.D.H. edited the manuscript and approved the final version.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Callaghan, N.I., Durland, L.J., Ireland, R.G. et al. Harnessing conserved signaling and metabolic pathways to enhance the maturation of functional engineered tissues. npj Regen Med 7, 44 (2022). https://doi.org/10.1038/s41536-022-00246-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41536-022-00246-3
