
Abstract
The underlying mechanisms by which renal denervation (RD) decreases blood pressure (BP) remain incompletely understood. In this study, we investigated the effects of ischemic kidney denervation on different sympathetic outflows, brain and renal expression of angiotensin-II receptors, oxidative stress and renal function markers in the 2-kidney, 1-clip (2K-1C) rat model. Surgical RD was performed in Wistar male rats 4–5 weeks after clip implantation. After 10 days of RD, BP, and the activity of sympathetic nerves projecting to the contralateral kidney (rSNA) and splanchnic region were partially reduced in 2K-1C rats, with no change in systemic renin-angiotensin system (RAS). To distinguish the effects of RD from the reduction in BP, 2K-1C rats were treated with hydralazine by oral gavage (25 mg/kg/day for 1 week). RD, but not hydralazine, normalized oxidative stress in the sympathetic premotor brain regions and improved intrarenal RAS, renal injury, and proteinuria. Furthermore, different mechanisms led to renal injury and oxidative stress in the ischemic and contralateral kidneys of 2K-1C rats. Injury and oxidative stress in the ischemic kidney were driven by the renal nerves. Although RD attenuated rSNA, injury and oxidative stress persisted in the contralateral kidney, probably due to increased BP. Therefore, nerves from the ischemic kidney at least partially contribute to the increase in BP, sympathetic outflows, brain oxidative stress, and renal alterations in rats with renovascular hypertension. Based on these findings, the reduction in oxidative stress in the brain is a central mechanism that contributes to the effects of RD on Goldblatt hypertension.
Similar content being viewed by others
Introduction
There is extensive evidence that sympathetic activation plays a critical role in the onset and maintenance of arterial hypertension. The importance of sympathoexcitation in hypertension has been reinforced by studies in humans showing that bilateral renal denervation (RD) leads to a sustained reduction in blood pressure (BP) [1, 2]. Additionally, a recent study provided biological proof of principle for the BP-lowering efficacy of RD in patients with uncontrolled hypertension in the absence of antihypertensive medications [3]. However, the underlying mechanisms by which RD decreases BP remain incompletely understood.
In addition to sympathetic innervation, the kidneys are also innervated by afferent fibers [4]. It is well-known that renal sympathetic fibers regulate renal tubular water and sodium reabsorption throughout the nephron, the constriction of the renal vasculature, and renin release from juxtaglomerular cells that activates the renin–angiotensin system (RAS) [5]. However, accumulating evidence suggests that renal afferent fibers project to the autonomic central nuclei and thereby influence the control of BP [6, 7]. Hence, the renal nerves mediate crosstalk between the brain and renal mechanisms that regulate BP under normal and pathological conditions [8, 9]. Thus, afferent nerves from the ischemic kidney may act by modulating neurotransmission in central cardiovascular nuclei in the brain [10], while efferent nerves are responsible for modulating kidney function, including sodium handling, renin secretion, and vasoconstriction [4]; altogether, these mechanisms are the major factors involved in the mechanism controlling cardiovascular function and hypertension. In fact, a positive relationship between renal afferent fibers and sympathetic overactivity has been observed in humans [11] and animals with hypertension, such as 2-kidney, 1-clip (2K-1C) rats [12, 13].
Sympathetic premotor neurons within the rostral ventrolateral medulla (RVLM) [14] and the paraventricular nucleus of the hypothalamus (PVN) [15] have been shown to be involved in the pathway regulating renal function. These cardiovascular brain nuclei contain a high density of angiotensin II (Ang II) type 1 receptors (AT1R) [16] through which brain Ang II produces several effects that influence cardiovascular function. In a model of renovascular hypertension, the activation of AT1R within the RVLM contributes to increased BP, renal sympathoexcitation and baroreflex dysfunction [17, 18] by increasing oxidative stress [19, 20]. However, researchers have not determined whether these mechanisms are mediated by renal nerves. In fact, renal ischemia and injury have been shown to increase afferent activity and Fos expression in the RVLM [21], and the electrical stimulation of renal afferents has been shown to trigger the activation of tyrosine hydroxylase (TH)-containing neurons in the RVLM and non-TH neurons in the nucleus of the solitary tract in normotensive rats [10]. Thus, we hypothesized that nerves from the ischemic kidney play an important role in the mechanisms that increase the activity of different sympathetic outflows and BP, such as Ang II receptor expression and brain oxidative stress.
In addition to brain oxidative stress, our previous study reported the induction of renal oxidative stress by the AT1R-NADPH oxidase pathway in the ischemic kidneys of 2K-1C rats [22]. Increased NADPH oxidase activity is also detected in patients with cardiovascular diseases [23, 24]. Oxidative stress accelerates the progression of renal injury and induces inflammatory responses in rats with chronic kidney disease (CKD) [25]. Moreover, RD modulates the renal expression of AT1R and AT2R [26], and Ang II receptors contribute to renal injury in rats with CKD [27]. Thus, we also hypothesized that renal nerves contribute to renal oxidative stress, injury and function, as well as Ang II receptor expression during renovascular hypertension. Myofibroblasts, which are alpha-smooth muscle actin (α-SMA)-expressing activated fibroblasts, play a major role in the development of glomerulosclerosis and interstitial fibrosis in both humans and experimental models of renal disease, representing a great prognostic indicator of disease progression [28,29,30]. Ang II promotes the shift from the fibroblast phenotype to the myofibroblast phenotype, the latter of which proliferate and invade the periglomerular and peritubular spaces, contributing to matrix deposition in the tubulointerstitial area [31, 32].
Several studies in humans and experimental models of cardiovascular diseases, such as hypertension and heart failure, have shown that RD improves the baroreflex control of renal sympathetic drive and other target organs [33,34,35]. In Dahl hypertensive rats, RD attenuates the development of left ventricle hypertrophy and renal damage and activation of the RAS independent of its effects on BP [36]. In CKD rats, RD prevents renal fibrosis and reduces sympathetic hyperactivity to splanchnic and lumbar nerves [37]. Thus, central mechanisms underlie the effects of RD on the cardiovascular system.
In the present study, we examined the effects of total denervation (afferent and efferent fibers) of the ischemic kidney on the levels of AT1R and AT2R and oxidative stress in brain cardiovascular nuclei, contralateral renal and splanchnic sympathetic nerve discharge, as well as renal changes in rats with renovascular hypertension.
Materials and methods
All experimental procedures described in this study were conducted according to the guidelines recommended by the National Institutes of Health and were approved by the Ethics in Research Committee of the Escola Paulista de Medicina—Universidade Federal de São Paulo—(process No. 1018/10). Male Wistar rats (150–180 g) were housed in groups of 4–5 in standard polypropylene cages in a room maintained at 22 ± 1 °C with a 12:12 h light-dark cycle (lights on at 7 am) and were allowed free access to food and water.
Experimental protocol
A silver clip (gap width of 0.2 mm) was implanted around the left renal artery of rats anesthetized with an i.p. injection of ketamine and xylazine (40 and 20 mg/kg, respectively) (Vetbrands, Jacareí, SP, Brazil). Age-matched rats were used as controls and did not undergo clip implantation. Hypertensive and control rats were divided into four independent groups: a control group that underwent sham surgery for RD, a 2K-1C group that underwent sham surgery for RD, a 2K-1C group that underwent RD surgery, and a 2K-1C group that underwent hydralazine treatment (oral dose of 25 mg/kg/day). Two sets of experiments were performed: one set for cardiovascular and nerve recordings and the other set for sample collection.
RD surgery
Unilateral denervation of the ischemic kidney of 2K-1C rats was performed 4–5 weeks after the implantation of the renal artery clip. Under ketamine and xylazine anesthesia, the left kidney was retroperitoneally exposed, and the renal artery and vein were carefully isolated from the connective tissue. After stripping all the visible nerves, the renal artery was painted with a solution of 10% phenol diluted in absolute ethanol. Rats were assessed 10 days after RD before complete reinnervation occurred [38].
Confirmation of RD
In a different series of experiments, four control rats were submitted to the RD surgery described above. After 10 days, the effectiveness of the procedure was determined by measuring the noradrenaline content in the denervated and innervated kidneys by using high-performance liquid chromatography (HPLC—Shimadzu, Kyoto, Japan) as previously described [39].
Hydralazine treatment
To distinguish the effects of RD from the effects of BP reduction, a direct-acting vasodilator was used to reduce BP. Five weeks after clip implantation, hydralazine (25 mg/kg/day) (Sigma-Aldrich Co., St. Louis, MO, USA) was orally administered by gavage. The treatment was performed once daily for seven consecutive days, and the administered dose was described in a previous report [40].
Recordings of mean arterial pressure and heart rate in conscious rats
Ten days after RD or after seven days of hydralazine treatment, rats were anesthetized with ketamine and xylazine; then, the femoral artery was catheterized to enable direct recordings of the arterial pressure, and the femoral vein was catheterized for drug infusions. After recovery from surgery (≥24 h), pulsatile arterial pressure was recorded in conscious rats using a BP signal amplifier (PowerLab System, ADInstruments, Sydney, Australia). The average values of baseline mean arterial pressure (MAP) and heart rate (HR) were obtained from the arterial pressure wave through a 15-min continuous recording.
Recordings of the activity of sympathetic nerves projecting to the contralateral kidney (rSNA) and splanchnic (sSNA) region in urethane-anesthetized rats
After recording the cardiovascular parameters in conscious rats, the animals were slowly anesthetized with urethane (1.2 g/kg, i.v.) (Sigma-Aldrich Co., St. Louis, MO, USA) to avoid any changes in baseline cardiovascular conditions. The splanchnic nerve and/or the right renal nerve in control rats or the contralateral kidney in 2K-1C rats were retroperitoneally exposed and placed on bipolar silver electrodes. When the conditions for nerve recordings were established, the nerves and electrodes were covered with paraffin oil. Nerve signals were displayed on an oscilloscope TDS 220 (Tektronix, Beaverton, OR, USA). The nerve activity was amplified (gain 20 K, Neurolog Digitimer, Welwyn Garden City, Herts, UK), filtered with a bandpass filter (100–1000 Hz), and collected for display and subsequent analysis using a PowerLab data acquisition system (ADInstruments, Sydney, Australia). At the end of the experiments, the background noise level was determined by administering hexamethonium bromide (30 mg/kg, i.v.) (Sigma-Aldrich). Then, the raw nerve signal was passed through a spike discriminator using Spike Histogram software (PowerLab, ADInstruments) to remove background noise, and the total nerve activity was expressed in spikes per second (spikes/s), as commonly used by previous studies that compared multifiber nerve recordings among different groups [41,42,43,44,45,46]. The number of spikes/burst reflects cardiovascular barosensitive fibers, as previously reported [47, 48]. The analysis of SNA by counting discriminated spikes/second above an established threshold may be influenced by the electrode contact with the nerve fibers; however, only experiments in which the level of background noise was confirmed at the end of the experiments following hexamethonium administration were included in this study.
Urine, blood, renal tissue, and brain tissue collection
In another set of experiments, rats were housed in metabolic cages (Nalgene, Ugo Basile, Italy) for the last 48 h of the experiments. The first 24 h were considered as the adaptation period to the cages, and the urine collected over the last 24 h was stored at −80 °C until subsequent analyses. Then, rats were decapitated to collect blood samples and renal and brain tissues. Left and right kidneys were removed and bisected transversely; half of the kidney was fresh frozen for the analysis of gene expression and dihydroethidium (DHE) staining, and the other half of the kidney was immersed in Bouin's solution for 24 h at room temperature. Brains were quickly removed, placed in a rat brain matrix (Insight, Ribeirão Preto, SP, Brazil), and frozen on dry ice. Coronal slices (500 µm) were prepared according to a rat brain atlas [49]: 12.5–12.7 mm from the bregma for the RVLM and 1.6–2.12 mm from the bregma for the PVN. Bilateral tissue punches were collected from the brain slices containing the RVLM and the PVN with a blunt needle (internal diameter: 0.5 mm) using a surgical microscope (×40 magnification) to identify each region. The RVLM region was bounded laterally by the spinal trigeminal tract, medially by the inferior olive and pyramids, and dorsally by the compact formation of the nucleus ambiguous. The 3rd ventricle was used as a reference to identify the PVN region.
Plasma renin activity assay
Blood samples were collected in tubes containing EDTA (5.0 mmol/L) and protease inhibitors (1.5 mmol/L ortho-phenanthroline and 10 µmol/L PMSF). Plasma was separated from blood cells by centrifugation at 2000g for 15 min at 4 °C. Plasma renin activity was determined by assessing the generation of angiotensin I using reverse-phase HPLC after incubating the plasma with a synthetic tetradecapeptide substrate at 37 °C.
Determination of plasma urea and creatinine concentrations
Plasma urea concentrations were determined with an enzymatic-colorimetric method using a commercially available kit (Labtest Diagnóstica, Lagoa Santa, Brazil), and plasma creatinine concentrations were measured by Jaffe's reaction.
Determination of proteinuria and plasma and urinary angiotensinogen concentrations
Total urinary protein excretion was measured with the pyrogallol red-molybdate method using a commercially available kit (Labtest Diagnóstica, Brazil). Plasma and urinary angiotensinogen concentrations were measured using a commercial solid-phase sandwich enzyme-linked immunosorbent assay (ELISA) (Immuno-Biological Laboratories, Japan). The urinary levels of angiotensinogen were normalized to the urinary creatinine content and reported as the angiotensinogen:creatinine ratio. All assays were performed according to the manufacturer's instructions.
Quantification of the levels of the AT1R and AT2R mRNAs in the RVLM and PVN, as well as the mRNA levels of oxidative stress markers in the ischemic and contralateral kidneys
Total RNA was extracted from bilateral punches of the RVLM and PVN or from the kidneys using the ReliaPrep™ RNA Tissue Miniprep System (Promega, USA) or Trizol reagent (Ambion-Life Technologies, USA), respectively, according to the manufacturer’s instructions. The reverse transcription reaction was performed using the High Capacity cDNA Reverse Transcription Kit (AB-Life Technologies, USA) according to the manufacturer’s directions. Quantitative real-time PCR (qRT-PCR) was conducted using the RotorGene Thermocycler (Qiagen, Dusseldorf, Germany) and the Power SybrGreen PCR Master Mix (AB-Life Technologies, USA) according to the manufacturer’s recommendations. The expression of target genes was normalized to HPRT1 mRNA expression levels that were measured concurrently. The primer list is available in the Supplementary material, and PCR parameters will be provided upon request.
Analysis of ROS production in the kidneys, RVLM and PVN
Reactive oxygen species (ROS) production was estimated using DHE staining. Fresh frozen renal and brain tissues were sectioned at 20 μm thickness using a cryostat. Sections were incubated with 20 µm DHE (Invitrogen, Carlsbad, CA) for 15 min at 37 °C and protected from light. The fluorescence intensity was quantified using a previously described method [17].
Analysis of renal injury
After fixation with Bouin's solution, the kidneys were dehydrated and embedded in paraffin using standard techniques, and then, sections were prepared at a thickness of 5 μm. Renal interstitial injury was determined by immunostaining with a monoclonal antibody for α-smooth muscle actin (α-SMA; 1:1000; Dako Corporation, Carpinteria, USA) and detected with a Dako LSAB System HRP and DAB kit (Dako Corporation). Sirius red staining was performed by incubating slides in a 0.1% Sirius red solution diluted in saturated picric acid (PSR) for 20 min. Slides were visualized with an Eclipse 80i microscope (Nikon Instruments, Melville, NY, USA) and all images were uniformly adjusted for brightness and contrast and captured with a Nikon digital sight DSRi1 camera. Images of 20 fields of the renal cortex (3–4 sections/slide; each section approximately 150 μm apart) at ×200 magnification (area/field: 276816.61 µm [2]) were blindly analyzed using the NIS-Elements software (Nikon Instruments). α-SMA expression and PSR staining were quantified as a percentage of the labeled area/field (vessels were excluded in the α-SMA analysis).
Statistical analysis
Values are presented as means ± standard deviations. Homogeneity of variance was assessed using the Bartlett test and the normal distribution of the data was analyzed using the Kolmogorov–Smirnov test. Data with an unequal distribution of the samples or a lack of homogeneity of variance were analyzed by Kruskal–Wallis ANOVA followed by the Mann–Whitney U test (α-SMA in the ischemic and contralateral kidneys and ROS production and AT1R gene expression in the ischemic kidney). The noradrenaline content was evaluated using the paired t test. The remaining data were evaluated with one-way ANOVA followed by the Newman–Keuls test. P < 0.05 was considered statistically significant.
Results
Effects of ischemic kidney denervation or hydralazine treatment on renal and cardiovascular parameters
As shown in Table 1, no significant differences in baseline HR or plasma angiotensinogen and renin activities were observed among groups. However, plasma creatinine levels were increased in the 2K-1C and hydralazine-treated 2K-1C groups. Denervation significantly reduced the plasma creatinine levels.
In the control group (n = 4), the intrarenal noradrenaline content showed that the RD procedure was successful (Fig. 1a). A significant, 95% reduction in the noradrenaline content was observed in the denervated kidney (5 ± 2 ng/g tissue) compared with the intact kidney (102 ± 57 ng/g tissue) 10 days after unilateral RD.

Measurements of the renal noradrenaline content (n = 4) (a), baseline mean arterial pressure (n = 8/group) (b), baseline contralateral kidney (n = 5/group) (c) and splanchnic (n = 5/group) (d) sympathetic nerve activity (SNA), urinary protein concentrations (n = 5/group) (e), angiotensinogen concentrations (n = 5 animals/group) (f), and representative tracings of baseline renal and splanchnic SNA following the normalization of the background noise level after hexamethonium administration (30 mg/kg, iv). *p < 0.05 compared with the control group. #p < 0.05 compared with the 2K-1C group
The MAP (n = 8 animals/group) was significantly higher in the 2K-1C rats (192 ± 12 mmHg) than in the control rats (112 ± 5 mmHg). Unilateral RD of the ischemic kidney (167 ± 15 mmHg) and hydralazine treatment (150 ± 9 mmHg) decreased the MAP compared to the hypertensive group, as shown in Fig. 1b. Increased baseline contralateral rSNA and sSNA (n = 5 animals/group) were observed in the 2K-1C group (rSNA: 180 ± 36 spikes/s; and sSNA: 98 ± 8 spikes/s) compared with the control group (rSNA: 103 ± 11 spikes/s; and sSNA: 69 ± 8 spikes/s). RD (rSNA: 134 ± 12 spikes/s and sSNA: 84 ± 5 spikes/s), but not the hydralazine treatment (rSNA: 174 ± 22 spikes/s and sSNA: 99 ± 13 spikes/s), significantly reduced rSNA and sSNA in the hypertensive group (Fig. 1c, d). Typical traces are shown in Fig. 1g.
Proteinuria (n = 5/group) was increased in the 2K-1C group (45 ± 13 mg/24 h) compared with control group (22 ± 3 mg/24 h). RD (23 ± 4 mg/24 h, n = 4), but not the hydralazine treatment (43 ± 17 mg/24 h), significantly reduced proteinuria (Fig. 1e). Urinary angiotensinogen concentrations (n = 5 animals/group) were increased in the nondenervated (38 ± 19 angiotensinogen:creatinine ratio) and hydralazine-treated (45 ± 15 angiotensinogen:creatinine ratio) 2K-1C groups compared with the control group (2 ± 1 angiotensinogen:creatinine ratio). After RD, but not the hydralazine treatment, urinary angiotensinogen concentrations were significantly reduced in the 2K-1C group (5 ± 3 angiotensinogen:creatinine ratio), as shown in Fig. 1f.
Effects of ischemic kidney denervation or hydralazine treatment on ROS production and the expression of Ang II receptors within the PVN and RVLM
ROS production (n = 4/group) was significantly increased in the PVN (173 ± 16%) and RVLM (148 ± 12%) of the 2K-1C group. RD (117 ± 15% and 101 ± 12%, respectively), but not the hydralazine treatment (148 ± 21% and 143 ± 18%, respectively), reduced ROS production in the PVN and RVLM. Representative images of ROS production are shown in Fig. 2a, b.

Representative images (a, b) and quantification of reactive oxygen species (ROS) levels (c, d) and the expression of the angiotensin II receptor type 1 (AT1R) (e, f) and 2 (AT2R) (g, h) mRNAs within the paraventricular nucleus of the hypothalamus (PVN) (n = 4/group) and rostral ventrolateral medulla (RVLM) (n = 4/group). *p < 0.05 compared with the control group. #p < 0.05 compared with the 2K-1C group
The expression of both the AT1R (twofold) and AT2R (twofold) mRNAs was significantly upregulated in the PVN (n = 4/group) of the hypertensive group (Fig. 2e, g, respectively). RD and the hydralazine treatment normalized the levels of both receptors in the PVN of 2K-1C rats. Within the RVLM (n = 4/group) of 2K-1C rats, the expression of the AT2R mRNA was significantly increased (1.7-fold) (Fig. 2h). Both RD and the hydralazine treatment normalized the expression of AT2R (Fig. 2h) and reduced the expression of AT1R (Fig. 2f) within the RVLM of the hypertensive groups.
Effects of ischemic kidney denervation or hydralazine treatment on injury, oxidative stress, and the expression of the NADPH oxidase subunit and Ang II receptors in the ischemic and contralateral kidneys
Differential responses to RD and hydralazine treatment were observed in the ischemic and contralateral kidneys. In both the ischemic and contralateral kidneys, renal injury, as indicated by α-SMA expression (Fig. 3a, b, respectively) and PSR staining (Fig. 3c, d, respectively), and oxidative stress, as indicated by DHE fluorescence (Fig. 3e, f, respectively), were increased in the 2K-1C group (n = 4). RD (n = 4), but not the hydralazine treatment (n = 4), decreased the levels of these markers in the ischemic kidney. However, in the contralateral kidney, both RD (n = 4) and the hydralazine treatment (n = 4) attenuated the levels of these markers. Representative images of renal immunohistochemistry and histology are shown in Fig. 4.

Quantification of α-smooth muscle actin (α-SMA) expression (a, b), picrosirius red (PSR) staining (c, d) and reactive oxygen species (ROS) production (e, f) in the ischemic (n = 4/group) and contralateral kidneys (n = 4/group). *p < 0.05 compared with the control group. #p < 0.05 compared with the 2K-1C group

Representative images of renal damage, as indicated by α-smooth muscle actin (α-SMA) expression and picrosirius red (PSR) staining, and reactive oxygen species (ROS) production, as indicated by dihydroethidium staining (DHE), in the ischemic and contralateral kidneys. The scale bar represents 50 µm
The expression (n = 4/group) of the p47phox (Fig. 5a), gp91phox (Fig. 5c), AT1R (Fig. 5e), and AT2R (Fig. 5g) mRNAs was significantly upregulated (2–3-fold) in the ischemic kidneys of the 2K-1C group. Both RD and the hydralazine treatment normalized the expression of these markers in the ischemic kidney. However, no significant changes were observed in the contralateral kidney (Fig. 5b, d, f, h).

Quantification of the expression of the mRNAs encoding NADPH oxidase subunits p47phox (a, b) and gp91phox (c, d) and angiotensin II receptor type 1 (AT1R) (e, f) and 2 (AT2R) (g, h) in the ischemic (n = 4 animals/group) and contralateral kidneys (n = 4/group). *p < 0.05 compared with the control group. #p < 0.05 compared with the 2K-1C group
Discussion
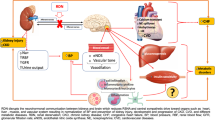
Based on the data from the present study, denervation of the ischemic kidneys in 2K-1C hypertensive rats reduces the baseline BP to some extent, without modifying plasma renin activity; however, a partial reduction in sympathoexcitation to the contralateral kidney and splanchnic region, as well as normalization of oxidative stress in the premotor sympathetic brain regions RVLM and PVN are observed. In addition to these effects, the reduction in renal angiotensinogen excretion and the levels of renal dysfunction markers induced by RD seems to be independent of its effects on reducing BP. Furthermore, the underlying mechanisms of renal injury and oxidative stress differ in the ischemic and contralateral kidneys of rats with renovascular hypertension. Injury and oxidative stress in the ischemic kidney are driven by the renal nerve and the increased expression of AT1R and NADPH oxidase subunits. Although rSNA to the contralateral kidney is reduced by the denervation of the ischemic kidney, injury and oxidative stress in the contralateral kidney are partially driven by the increase in BP and not by AT1R and NADPH oxidase subunit expression.
Our findings that total denervation (afferent and efferent fibers) of the ischemic kidney reduced BP to some extent without reducing plasma renin activity are consistent with the data reported by Katholi et al. [50]. Although the authors observed increased plasma renin activity in the 2K1C group, this parameter was not changed after RD [50]. Moreover, denervation of the ischemic kidney, but not the contralateral kidney, reduced systolic BP to some extent without altering creatinine clearance or sodium excretion [50]. Furthermore, these findings support the hypothesis that the ischemic kidney contributes to the pathogenesis of renovascular hypertension in the 2K-1C model by stimulating renal afferent fibers, leading to enhanced activity of the sympathetic nervous system. Indeed, selective afferent RD by thoracolumbar dorsal rhizotomy decreases BP in the 2K-1C model [13]. Although we have not performed the selective denervation of afferent fibers, the current study shows that the removal of both the afferent and efferent fibers of the ischemic kidney decreases the contralateral rSNA and sSNA, and the relative contributions of afferent and efferent fibers remain to be elucidated. In humans, diseased kidneys display considerable contributions to the elevated sympathetic nerve activity in renal transplant recipients, which was probably mediated by renal afferent nerves [11]. Renal transplantation patients with excellent graft function only achieved normalization of the muscle sympathetic nerve activity after bilateral kidney nephrectomy [11]. In the renovascular model, afferent activity from the contralateral kidney leads to a normal inhibitory renorenal reflex in the ischemic kidney that is similar to the healthy kidney, while increased afferent activity from the ischemic kidney leads to an abnormal excitatory reflex that increases sympathetic drive to the contralateral kidney [51]. Thus, changes in the neural activity from the ischemic kidney trigger a disproportionate increase in sympathetic outflows in 2K-1C rats that may be integrated in the brain, particularly in the sympathetic premotor regions.
In 2K-1C rats, AT1R expressed in the PVN and RVLM is a major mediator of renal sympathoexcitation and hypertension [17, 18, 52]. The stimulation of AT2R counteracts most of the effects of AT1R stimulation within the PVN and RVLM of normal and chronic heart failure animals [53]. Surprisingly, both AT1R and AT2R were upregulated within the PVN of 2K-1C rats, and RD and the hydralazine-induced BP reduction decreased Ang II receptor expression in both the RVLM and PVN of the hypertensive group. The roles of AT2R in these brain regions in rats with renovascular hypertension remains unclear. However, based on our data, changes in AT1R and AT2R expression are mostly mediated by changes in BP rather than RD.
Interestingly, the reduction in oxidative stress observed in the sympathetic premotor regions after RD may contribute to the reductions in BP and the activity of sympathetic nerves projecting to the contralateral kidney and the splanchnic bed. Oxidative stress and activation of AT1R in the RVLM and PVN of rats with renovascular hypertension are major contributors to sympathoexcitation [19, 54]. The downregulation of AT1R in the PVN and RVLM may contribute to the observed reductions in oxidative stress in the brain and sympathoexcitation in the denervated hypertensive group but not in the hydralazine-treated group. However, further studies are needed to determine whether the denervation of ischemic kidney directly or indirectly reduces oxidative stress in the brains of 2K-1C rats. Moreover, other receptors, such as 5-HT [55] or opioid [56] receptors, are known to influence the hypertensive state in rats with renovascular hypertension; thus, we cannot exclude the possibility that other mechanisms are involved in reducing blood pressure after RD.
In addition to the reduced contralateral rSNA and sSNA after RD, we observed the amelioration of renal function and the reduction of proteinuria and urinary angiotensinogen concentrations in 2K-1C rats. These effects probably occurred independently of the effects of RD on reducing BP, because the hydralazine treatment significantly reduced BP but did not change these parameters. The urinary angiotensinogen concentration is an indicator of intrarenal RAS activation in hypertensive rodents [57, 58] and patients [59]. Although urinary proteins and angiotensinogen are derived from both the ischemic and contralateral kidneys in the 2K-1C group, the upregulation of AT1R observed only in the ischemic kidney, along with our previous studies showing increased mRNAexpression of the AT1R [22] and ACE and angiotensinogen protein expressions [60] only in the ischemic kidney, potentially indicates that RAS is activated in this kidney. However, further experiments in which the left and right ureters are cannulated are required to determine the different effects of each kidney on proteinuria,
We observed normal levels of plasma renin activity and increased intrarenal RAS activation in the 2K-1C group, as previously reported in other studies [61, 62]. Moreover, although RD did not change plasma renin activity, the activity of the intrarenal RAS was attenuated after RD, which may have contributed to the decrease in BP and improvement in renal function and proteinuria in the 2K-1C group. Interestingly, 2K-1C rats with a moderate increase in BP present increased levels of circulating Ang II and normal renal function; moreover, ischemic kidney denervation by cryoablation normalizes circulating Ang II levels and contralateral rSNA after 3 weeks [63]. Despite the differences in the literature regarding circulating RAS levels in rats with renovascular hypertension, we show that the effects of RD on the activity of the intrarenal RAS and renal function markers were accompanied by reductions in renal injury and oxidative stress in the ischemic kidney.
Our data show that renal damage and oxidative stress are induced by distinct mechanisms in the ischemic and contralateral kidneys of 2K-1C rats. We observed that α-SMA, PSR, and ROS production were partially reduced in the ischemic kidney after RD, and the levels of these markers were also decreased to some extent by the BP reduction induced by the hydralazine treatment in the contralateral kidneys of hypertensive rats. Although oxidative stress is mediated by the AT1R-NADPH oxidase pathway in the ischemic kidney [22], our data indicate that this axis/pathway is also sensitive to changes in BP. Both RD and the hydralazine treatment decreased the expression of the p47phox, gp91phox, AT1R, and AT2R mRNAs in the ischemic kidney, whereas the levels of these markers were unchanged in the contralateral kidneys of the 2K-1C group. Previous studies have shown that elevated renal perfusion pressure was the dominant cause of renal damage in both noradrenaline- and Ang II-induced hypertensive rats [64], and both Ang II and increased renal perfusion pressure induce the renal activation of NADPH oxidase and oxidative stress [65]. Nevertheless, further experiments are necessary to verify whether the amelioration of renal injury and oxidative stress are direct effects of RD, a consequence of attenuated intrarenal RAS, or both in rats with renovascular hypertension. According to the results of an in vitro study, exposure to noradrenaline for 24 h significantly and dose-dependently increased angiotensinogen mRNA levels in immortalized human proximal tubular cells [66].
Therefore, nerves from the ischemic kidney at least partially contribute to the development of hypertension, high contralateral rSNA and sSNA and renal alterations, such as proteinuria and intrarenal RAS activation, in the 2K-1C model of renovascular hypertension. Reductions in oxidative stress and levels of Ang II receptors in the RVLM and PVN induced by RD may contribute to the observed decreases in hypertension and sympathetic hyperactivity in 2K1C rats.
Limitations
In the present study, the SNA was compared between groups from baseline and expressed as spikes/s. Although we recognize that this method may be a limitation, the SNA baseline variation expressed as spikes/s among individuals using this approach was relatively small and highly responsive to baroreceptor reflex loading or unloading and was abolished by the hexamethonium administration confirming that the rSNA is a very homogenous population of fibers, consistent with a previous report [67]. Increased SNA was previously reported using an assessment of noradrenaline spillover in humans [68], an µV/s analysis [63], and by calculating the percentage of maximum rSNA response [69] in rats with renovascular hypertension. Moreover, RD, ganglionic blockade or RVLM inhibition lead to a larger decrease in BP in 2K1C rats compared to controls [43, 70, 71] and support our finding that rSNA is significantly increased in the renovascular hypertension model. Another limitation of the present study is the number of animals used in some experiments. For the data obtained from n = 4 animals, we calculated the power analysis using G*Power software (Düsseldorf, Germany) and the value ranged from 60 to 80%. The ideal power analysis is ≥80% to avoid increased type II error. However, the gene expression analysis in the RVLM and PVN bilateral punches requires a pool of 3 animals per sample and therefore the ethics committee of our university recommended the use of 12 animals per group in our study to adhere to the 3Rs Principle.
References
Krum H, Schlaich MP, Sobotka PA, Böhm M, Mahfoud F, Rocha-Singh K, et al. Percutaneous renal denervation in patients with treatment-resistant hypertension: final 3-year report of the Symplicity HTN-1 study. Lancet. 2014;383:622–9.
Esler M, Guo L. The future of renal denervation. Auton Neurosci Basic Clin. 2016; https://doi.org/10.1016/j.autneu.2016.08.004.
Townsend RR, Mahfoud F, Kandzari DE, Kario K, Pocock S, Weber MA. et al. Catheter-based renal denervation in patients with uncontrolled hypertension in the absence of antihypertensive medications (SPYRAL HTN-OFF MED): a randomised, sham-controlled, proof-of-concept trial. Lancet. 2017; https://doi.org/10.1016/S0140-6736(17)32281-X). (e-pub ahead of print August 2017).
Kopp UC Neural Control of Renal Function. 1st ed. San Rafael (CA): Morgan & Claypool Publishers. https://www.ncbi.nlm.nih.gov/books/NBK57113/.
DiBona GF, Kopp UC. Neural control of renal function. Physiol Rev. 1997;77:75–197. https://doi.org/10.1152/physrev.1997.77.1.75.
Ye S, Ozgur B, Campese VM. Renal afferent impulses, the posterior hypothalamus, and hypertension in rats with chronic renal failure. Kidney Int. 1997;51:722–7. http://www.ncbi.nlm.nih.gov/pubmed/9067904.
Ciriello J, de Oliveira CVR. Renal afferents and hypertension. Curr Hypertens Rep. 2002;4:136–42. http://www.ncbi.nlm.nih.gov/pubmed/11884269.
Johns EJ. The neural regulation of the kidney in hypertension and renal failure. Exp Physiol. 2014;99:289–94.
Nishi EE, Bergamaschi CT, Campos RR. The crosstalk between the kidney and the central nervous system: the role of renal nerves in blood pressure regulation. Exp Physiol. 2015;100:479–84.
Nishi EE, Martins BS, Milanez MIO, Lopes NR, de Melo JF, Pontes RB, et al. Stimulation of renal afferent fibers leads to activation of catecholaminergic and non-catecholaminergic neurons in the medulla oblongata. Auton Neurosci Basic Clin. 2017;204:48–56.
Hausberg M. Sympathetic nerve activity in end-stage renal disease. Circulation. 2002;106:1974–9.
Katholi RE, Winternitz SR, Oparil S. Decrease in peripheral sympathetic nervous system activity following renal denervation or unclipping in the one-kidney one-clip Goldblatt hypertensive rat. J Clin Invest. 1982;69:55–62.
Oparil S, Sripairojthikoon W, Wyss JM. The renal afferent nerves in the pathogenesis of hypertension. Can J Physiol Pharmacol. 1987;65:1548–58.
Ding ZQ, Li YW, Wesselingh SL, Blessing WW. Transneuronal labelling of neurons in rabbit brain after injection of herpes simplex virus type 1 into the renal nerve. J Auton Nerv Syst. 1993;42:23–31.
Haselton JR, Vari RC. Neuronal cell bodies in paraventricular nucleus affect renal hemodynamics and excretion via the renal nerves. Am J Physiol. 1998;275:R1334–42.
Allen AM, Moeller I, Jenkins TA, Zhuo J, Aldred GP, Chai SY, et al. Angiotensin receptors in the nervous system. Brain Res Bull. 1998;47:17–28.
Nishi EE, Bergamaschi CT, Oliveira-Sales EB, Simon KA, Campos RR. Losartan reduces oxidative stress within the rostral ventrolateral medulla of rats with renovascular hypertension. Am J Hypertens. 2013;26:858–65.
de Oliveira-Sales EB, Nishi EE, Boim MA, Dolnikoff MS, Bergamaschi CT, Campos RR. Upregulation of AT1R and iNOS in the rostral ventrolateral medulla (RVLM) is essential for the sympathetic hyperactivity and hypertension in the 2K-1C Wistar rat model. Am J Hypertens. 2010;23:708–15.
Campos RR, Oliveira-Sales EB, Nishi EE, Paton JFR, Bergamaschi CT. Mechanisms of renal sympathetic activation in renovascular hypertension. Exp Physiol. 2015;100:496–501.
Campos RR, Oliveira-Sales EB, Nishi EE, Boim MA, Dolnikoff MS, Bergamaschi CT. The role of oxidative stress in renovascular hypertension. Clin Exp Pharmacol Physiol. 2011;38:144–52.
Ding YF, Zhang XX, Shi GM, He RR. Renal ischemia enhances electrical activity and Fos protein expression of the rostral ventrolateral medullary neurons in rats. Sheng Li Xue Bao. 2001;53:369. http://www.ncbi.nlm.nih.gov/pubmed/11833421.
Nishi EE, Oliveira-Sales EB, Bergamaschi CT, Oliveira TGC, Boim MA, Campos RR. Chronic antioxidant treatment improves arterial renovascular hypertension and oxidative stress markers in the kidney in wistar rats. Am J Hypertens. 2010; 23. https://doi.org/10.1038/ajh.2010.11.
Ravarotto V, Pagnin E, Maiolino G, Fragasso A., Carraro G, Rossi B, Calo L a. The blocking of angiotensin II type 1 receptor and RhoA/Rho kinase activity in hypertensive patients: effect of olmesartan medoxomil and implication with cardiovascular-renal remodeling. J Renin Angiotensin Aldosterone Syst. 2015; 1–6. https://doi.org/10.1177/1470320315594324.
Avogaro A, Pagnin E, Calò L. Monocyte NADPH oxidase subunit p22phox and inducible hemeoxygenase-1 gene expressions are increased in type II diabetic patients: Relationship with oxidative stress. J Clin Endocrinol Metab. 2003;88:1753–9.
Modaresi A, Nafar M, Sahraei Z. Oxidative stress in chronic kidney disease. Iran J Kidney Dis. 2015;9:165–79.
Clayton SC, Haack KKV, Zucker IH. Renal denervation modulates angiotensin receptor expression in the renal cortex of rabbits with chronic heart failure. Am J Physiol Ren Physiol. 2011;300:F31–9.
Naito T, Ma L-J, Yang H, Zuo Y, Tang Y, Han JY, et al. Angiotensin type 2 receptor actions contribute to angiotensin type 1 receptor blocker effects on kidney fibrosis. Am J Physiol Ren Physiol. 2010;298:F683–F691.
Zhang G, Moorhead PJ, el Nahas AM. Myofibroblasts and the progression of experimental glomerulonephritis. Exp Nephrol. 1995;3:308–18.
Roberts IS, Burrows C, Shanks JH, Venning M, McWilliam LJ. Interstitial myofibroblasts: predictors of progression in membranous nephropathy. J Clin Pathol. 1997;50:123. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC499736/.
Alpers CE, Hudkins KL, Floege J, Johnson RJ. Human renal cortical interstitial cells with some features of smooth muscle cells participate in tubulointerstitial and crescentic glomerular injury. J Am Soc Nephrol. 1994;5:201–9. http://www.ncbi.nlm.nih.gov/pubmed/7993999.
Mezzano Sa, Ruiz-Ortega M, Egido J. Angiotensin II and renal fibrosis. Hypertension. 2001;38:635–8.
Johnson RJ, Alpers CE, Yoshimura A, Lombardi D, Pritzl P, Floege J, et al. Renal injury from angiotensin II-mediated hypertension. Hypertension. 1992;19:464–74.
Booth LC, Schlaich MP, Nishi EE, Yao ST, Xu J, Ramchandra R, et al. Short-term effects of catheter-based renal denervation on cardiac sympathetic drive and cardiac baroreflex function in heart failure. Int J Cardiol 2015; 190. https://doi.org/10.1016/j.ijcard.2015.03.440.
Hart EC, McBryde FD, Burchell AE, Ratcliffe LE, Stewart LQ, Baumbach A, et al. Translational examination of changes in baroreflex function after renal denervation in hypertensive rats and humans. Hypertension. 2013;62:533–41.
Schiller AM, Haack KK, Pellegrino PR, Curry PL, Zucker IH. Unilateral renal denervation improves autonomic balance in conscious rabbits with chronic heart failure. Am J Physiol Regul Integr Comp Physiol. 2013;305:R886–92.
Watanabe H, Iwanaga Y, Miyaji Y, Yamamoto H, Miyazaki S. Renal denervation mitigates cardiac remodeling and renal damage in Dahl rats: a comparison with β-receptor blockade. Hypertens Res. 2016;39:217–26.
Veiga GL, Nishi EE, Estrela HF, Lincevicius GS, Gomes GN, Simões Sato AY, et al. Total renal denervation reduces sympathoexcitation to different target organs in a model of chronic kidney disease. Auton Neurosci Basic Clin. 2017; 204. https://doi.org/10.1016/j.autneu.2016.11.006.
Mulder J, Hökfelt T, Knuepfer MM, Kopp UC. Renal sensory and sympathetic nerves reinnervate the kidney in a similar time-dependent fashion after renal denervation in rats. Am J Physiol Regul Integr Comp Physiol. 2013;304:R675–82.
Cavalheiro EA, Fernandes MJ, Turski L, Naffah-Mazzacoratti MG. Spontaneous recurrent seizures in rats: amino acid and monoamine determination in the hippocampus. Epilepsia. 1994;35:1–11.
Intengan HD, Schiffrin EL. Disparate effects of carvedilol versus metoprolol treatment of stroke-prone spontaneously hypertensive rats on endothelial function of resistance arteries. J Cardiovasc Pharmacol. 2000;35:763–8.
Morgan DA, Anderson EA, Mark AL. Renal sympathetic nerve activity is increased in obese Zucker rats. Hypertension. 1995;25:834–8.
Morgan DA, Despas F, Rahmouni K. Effects of leptin on sympathetic nerve activity in conscious mice. Physiological Reports. 2015;3:e12554. https://doi.org/10.14814/phy2.12554.
Oliveira-Sales EB, Nishi EE, Carillo BA, Boim MA, Dolnikoff MS, Bergamaschi CT, Campos RR. Oxidative stress in the sympathetic premotor neurons contributes to sympathetic activation in renovascular hypertension. Am J Hypertens. 2009;22:484–92. https://doi.org/10.1038/ajh.2009.17.
Ramchandra R, Hood SG, Denton DA, Woods RL, McKinley MJ, McAllen RM, et al. Basis for the preferential activation of cardiac sympathetic nerve activity in heart failure. Proc Natl Acad Sci USA. 2009;106:924–8.
Wei S-G, Felder RB. Forebrain renin-angiotensin system has a tonic excitatory influence on renal sympathetic nerve activity. Am J Physiol Heart Circ Physiol. 2002;282:H890–5.
Iwashita S, Tanida M, Terui N, Ootsuka Y, Shu M, Kang D, et al. Direct measurement of renal sympathetic nervous activity in high-fat diet-related hypertensive rats. Life Sci. 2002;71:537–46.
Malpas SC, Ninomiya I. A new approach to analysis of synchronized sympathetic nerve activity. Am J Physiol. 1992;263:H1311–7.
Malpas SC, Ninomiya I. The amplitude and periodicity of synchronized renal sympathetic nerve discharges in anesthetized cats: differential effect of baroreceptor activity. J Auton Nerv Syst. 1992;40:189–98.
Paxinos G, Watson CR. The rat brain in stereotaxic coordinates. 6th ed. Elsevier Academic Press: London.
Katholi RE, Whitlow PL, Winternitz SR, Oparil S. Importance of the renal nerves in established two-kidney, one clip Goldblatt hypertension. Hypertension. 1982;4:166. http://www.ncbi.nlm.nih.gov/pubmed/6175572.
Kopp UC, Buckley-Bleiler RL. Impaired renorenal reflexes in two-kidney, one clip hypertensive rats. Hypertension. 1989;14:445. http://www.ncbi.nlm.nih.gov/pubmed/2793217.
Chen A-D, Zhang S-J, Yuan N, Xu Y, De W, Gao X-Y, et al. Angiotensin AT1 receptors in paraventricular nucleus contribute to sympathetic activation and enhanced cardiac sympathetic afferent reflex in renovascular hypertensive rats. Exp Physiol. 2011;96:94–103.
Gao L, Zucker IH. AT2 receptor signaling and sympathetic regulation. Curr Opin Pharmacol. 2011;11:124–30.
Oliveira-Sales EB, Colombari DS, Davisson RL, Kasparov S, Hirata AE, Campos RR, et al. Kidney-induced hypertension depends on superoxide signaling in the rostral ventrolateral medulla. Hypertension. 2010;56:290–6.
Bergamaschi CT, Silva NF, Pires JG, Campos RR, Neto HAF. Participation of 5-HT and AT1 receptors within the rostral ventrolateral medulla in the maintenance of hypertension in the Goldblatt 1 kidney-1 clip model. Int J Hypertens. 2014;2014:723939.
Yeganeh-Hajahmadi M, Najafipour H, Rostamzadeh F The differential effects of low and high doses of apelin through opioid receptors on the blood pressure of rats with renovascular hypertension. Hypertens Res. 2017; 40. https://doi.org/10.1038/hr.2017.28.
Kobori H, Katsurada A, Miyata K, Ohashi N, Satou R, Saito T, et al. Determination of plasma and urinary angiotensinogen levels in rodents by newly developed ELISA. Am J Physiol Ren Physiol. 2008;294:F1257–63.
Kobori H, Harrison-Bernard LM, Navar LG. Enhancement of angiotensinogen expression in angiotensin II-dependent hypertension. Hypertension. 2001;37:1329–35.
Katsurada A, Hagiwara Y, Miyashita K, Satou R, Miyata K, Ohashi N, et al. Novel sandwich ELISA for human angiotensinogen. Am J Physiol Ren Physiol. 2007;293:F956–60.
Shimoura CG, Lincevicius GS, Nishi EE, Girardi ACC, Simon KA, Bergamaschi CT, et al. Increased dietary salt changes baroreceptor sensitivity and intrarenal renin-angiotensin system in goldblatt hypertension. Am J Hypertens. 2017;30:28–36.
Burke SL, Head GA, Lambert GW, Evans RG. Renal sympathetic neuroeffector function in renovascular and angiotensin II-dependent hypertension in rabbits. Hypertension. 2007;49:932–8.
Freeman RH, Davis JO, Watkins BE, Lohmeier TE. Mechanisms involved in two-kidney renal hypertension induced by constriction of one renal artery. Circ Res. 1977;40:I29–35.
Rossi NF, Pajewski R, Chen H, Littrup PJ, Maliszewska-Scislo M. Hemodynamic and neural responses to renal denervation of the nerve to the clipped kidney by cryoablation in two-kidney, one-clip hypertensive rats. Am J Physiol Regul Integr Comp Physiol. 2016;310:R197–208.
Polichnowski AJ, Cowley AW. Pressure-induced renal injury in angiotensin II versus norepinephrine-induced hypertensive rats. Hypertension. 2009;54:1269–77.
Polichnowski AJ, Jin C, Yang C, Cowley AW. Role of renal perfusion pressure versus angiotensin ii on renal oxidative stress in angiotensin II-induced hypertensive rats. Hypertension. 2010;55:1425–30.
Rafiq K, Noma T, Fujisawa Y, Ishihara Y, Arai Y, Nabi AH, et al. Renal sympathetic denervation suppresses de novo podocyte injury and albuminuria in rats with aortic regurgitation. Circulation. 2012;125:1402–13.
Montano N, Furlan R, Guzzetti S, Mcallen RM, Julien C. Analysis of sympathetic Neural discharge in rats and humans. Philos Trans R Soc A Math Phys Eng Sci. 2009;367:1265–82.
Johansson M, Elam M, Rundqvist B, Eisenhofer G, Herlitz H, Lambert G, et al. Increased sympathetic nerve activity in renovascular hypertension. Circulation. 1999;99:2537–42.
Zhu GQ, Xu Y, Zhou LM, Li YH, Fan LM, Wang W, et al. Enhanced cardiac sympathetic afferent reflex involved in sympathetic overactivity in renovascular hypertensive rats. Exp Physiol. 2009;94:785–94.
Bergamaschi C, Campos RR, Schor N, Lopes OU. Role of the rostral ventrolateral medulla in maintenance of blood pressure in rats with Goldblatt hypertension. Hypertension. 1995;26:1117–20.
Lincevicius GS, Shimoura CG, Nishi EE, Oliveira T, Cespedes JG, Bergamaschi CT, et al. Differential effects of renal denervation on arterial baroreceptor function in Goldblatt hypertension model. Auton Neurosci Basic Clin. 2017;208:43–50.
Acknowledgments
We acknowledge American Journal Experts for providing English language editing services. This work was supported by grants from the São Paulo Research Foundation (FAPESP 13/22522-9), CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), and Brazilian National Research Council (CNPq). EEN is a recipient of a FAPESP (10/09003-4) PhD scholarship. NRL is a recipient of a FAPESP undergraduate scholarship (15/23858-6). RRC, JCP, and CTB are recipients of the CNPq productive research fellowship.
Author contributions
Conception or design of the study: EEN, CTB, and RRC. Acquisition and analysis of qRT-PCR data: NRL and AYS. Measurement and analysis of noradrenaline concentrations: MGN. Acquisition, analysis, and interpretation of the data reported in the study: EEN, NRL, GNG, and JCP. Drafting the manuscript: EEN and RRC. All authors revised the manuscript critically for important intellectual content, approved the final version of the manuscript, agreed to be accountable for all aspects of the work and ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved and all persons designated as authors qualify for authorship, and all individuals who qualify for authorship are listed.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Nishi, E.E., Lopes, N.R., Gomes, G.N. et al. Renal denervation reduces sympathetic overactivation, brain oxidative stress, and renal injury in rats with renovascular hypertension independent of its effects on reducing blood pressure. Hypertens Res 42, 628–640 (2019). https://doi.org/10.1038/s41440-018-0171-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41440-018-0171-9
Keywords:
This article is cited by
-
Effects of renal denervation on blood pressures in patients with hypertension: a systematic review and meta-analysis of randomized sham-controlled trials
Hypertension Research (2022)
-
Neural suppression of miRNA-181a in the kidney elevates renin expression and exacerbates hypertension in Schlager mice
Hypertension Research (2020)
-
Anti-hypertensive effect of hydrogen peroxide acting centrally
Hypertension Research (2020)
-
Afferent innervation of the ischemic kidney contributes to renal dysfunction in renovascular hypertensive rats
Pflügers Archiv - European Journal of Physiology (2020)
-
Melatonin as a rational alternative in the conservative treatment of resistant hypertension
Hypertension Research (2019)
