
Abstract
De novo DDX3X variants account for 1–3% of syndromic intellectual disability (ID) in females and have been occasionally reported in males. Furthermore, somatic DDX3X variants occur in several aggressive cancers, including medulloblastoma. We report three unrelated females with severe ID, dysmorphic features, and a common brain malformative pattern characterized by malformations of cortical development, callosal dysgenesis, basal ganglia anomalies, and midbrain–hindbrain malformations. A pilocytic astrocytoma was incidentally diagnosed in Patient 1 and trigonocephaly was found in Patient 2. With the use of family based whole exome sequencing (WES), we identified three distinct de novo variants in DDX3X. These findings expand the phenotypic spectrum of DDX3X-related disorders, demonstrating unique neuroradiological features resembling those of the tubulinopathies, and support a role for DDX3X in neuronal development. Our observations further suggest a possible link between germline DDX3X variants and cancer development.
Similar content being viewed by others

Introduction
De novo DDX3X variants have been recently reported in 1–3% of females with unexplained intellectual disability (ID) [1, 2]. DDX3X maps to Xp11.3–11.23 and is ubiquitously transcribed in human tissues, escaping X-inactivation [1, 3]. It encodes an ATP-dependent ‘DEAD-box’ RNA helicase involved in RNA processing through secondary structure remodeling, with a pivotal role in the regulation of gene expression, cell cycle control, innate immunity, and viral replication [4,5,6]. We present three unrelated females with ID, dysmorphic features, and a peculiar brain malformative pattern, thus expanding the phenotypic spectrum of DDX3X-related disorders.
Patients and methods
The study was approved by the Institutional Ethics Committee and written informed consent was obtained according to Telethon Undiagnosed Program guidelines. Whole exome sequencing (WES) was performed by using the Agilent SureSelect QXT Clinical Research Exome (Agilent Technologies, Santa Clara, CA, USA). Sequencing data were processed using an in-house software for the execution of the GATK Best Practices pipeline for WES variant analysis (see Supplementary Material). After filtering for population genetics, family segregation, and predicted impact on protein sequence, the best candidates for all probands were de novo variants in DDX3X. The identified variants were validated by Sanger sequencing in all members of the trios and submitted into the gene variant database LOVD at https://www.LOVD.nl/DDX3X (Patient IDs 00219101, 00219102, 00219099).
Results
Clinical and neuroimaging findings
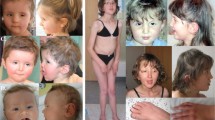
Patient 1 was the second-born to unrelated healthy parents with a healthy son. Antenatal history revealed intrauterine growth retardation (IUGR) due to placental insufficiency. She was delivered at 30+5 weeks’ gestation. Her birth weight (BW) was 890 g (−1.79 SD), length 38 cm (−0.66 SD), and occipito-frontal circumference (OFC) 27 cm (−0.65 SD). Neonatal course was uneventful. During childhood, global DD was noticed and she developed spastic tetraparesis and severe scoliosis. At the age of 8 years, she was incidentally diagnosed with a pilocytic astrocytoma (PA) (WHO grade I). Physical examination at 11 years of age revealed occipital plagiocephalywith normal OFC (51.5 cm, −0.96 SD), several dysmorphisms (Fig. 1a, panel a), absent speech, spastic tetraparesis, strabismus, and horizontal nystagmus.

a Dysmorphic facial features of the reported patients with de novo DDX3X variants: a Patient 1 presents with occipital plagiocephaly, frizzy hair, flat face with midface hypoplasia, and short neck. Dysmorphic facial features include: arched eyebrows, mild esotropia of the left eye, low-set wide and simplified ears, long smooth philtrum, thin lips and upper vermillion border, absent cupid bow, and retrognathia; b Dysmorphic features of Patient 2 include prominent metopic ridge with glabellar nevus simplex, arched eyebrows with synophrys, hypotelorism with epicanthal folds, strabismus, low-set posteriorly rotated ears, malar hypoplasia, and microretrognathia; c Clinical photographs of Patient 3 showing arched eyebrows, midface hypoplasia, large ears, long philtrum, and retrognathia. b Brain MRI findings: a, b Patient 1 at the age of 8 years; c, d Patient 2 at the age of 2.5 years; e, f Patient 3 at the age of 12 years. In all patients, axial images reveal malformations of cortical development, characterized by bilateral frontal polymicrogyria in Patient 1 (a) and Patient 3 (b). In Patient 2, axial T1-weighted sequences show an abnormal fronto-insular gyral pattern (c). The anterior limb of the internal capsule is very small with dysmorphic appearance of the basal ganglia (a, c, d, arrowheads). The periventricular white matter is reduced, especially in Patients 1 and 2, with consequent enlargement of the lateral ventricles, especially in frontal and anterior temporal regions. Sagittal T1-weighted images demonstrate severe callosal hypodysgenesis with prevalent involvement of the isthmus and splenium in Patient 1 (b, thick arrow) and 3 (d, thick arrow), and milder callosal hypoplasia in Patient 2 (f, thick arrow). Note the marked hypoplasia of the anterior commissure in patient 1 and 2, and anterior commissure agenesis in Patient 3. There is pontine hypoplasia in all patients (arrows) associated with hypoplasia and mild rotation of the inferior portion of the vermis (open arrows, Patients 2 and 3). In patient 1 sagittal T1-weighted image demonstrates a hypointense cerebellar mass lesion (detailed depiction available in online Supplementary Fig. 1). c DDX3X gene transcripts (top) and DDX3X protein (bottom): a UCSC genome browser map (GRCh37/hg19) shows multiple gene products of DDX3X, with alternative splicing resulting in multiple transcript variants. The gene has a 5′−3′ orientation. The main transcript (black rectangle) of 16,874 bp is encoded by 17 exons; b Schematic, not to scale, representation of the domains of the full-length DDX3X protein and localization of the variants identified in the reported patients. N- and C-terminus are variable regions. The consensus eIF4E-binding sequence is important for the interaction with the eukaryotic initiation factor 4E (eIF4E), a translation initiation factor modulated by DDX3X. The RecA-like domains 1 and 2 are essential for the helicase activity. Each of these catalytic domains is composed of different conserved functional motifs, involved in ATP and RNA binding. aa amino acid
Patient 2 was the only child of unrelated healthy parents. Prenatal history revealed IUGR, increased nuchal translucency, oligohydramnios, and ventriculomegaly. She was born at 34+2 weeks’ gestation. Her BW was 1,550 g (−1.46 SD), length 38 cm (−2.5 SD), and OFC 29 cm (−1.39 SD). Neonatal course was complicated by diffuse hypotonia and feeding difficulties. Brain ultrasonography confirmed isolated ventriculomegaly. Electroencephalogram (EEG) showed slow waves in the right cerebral hemisphere. Bilateral sensorineural hearing loss was found and fundoscopic examination showed optic subatrophy, but visual evoked potentials were normal. At 2 years of age, she had severe ID/DD, dysmorphic features, and microcephaly (OFC 43.2 cm, −3.13 SD) with prominent metopic bridge (Fig. 1a, panel b). Physical examination showed dysphagia, alternating esotropia, axial hypotonia, and generalized dystonia.
Patient 3 was the only daughter of non-consanguineous healthy parents. Pregnancy was complicated by IUGR due to placental insufficiency. She was delivered at 38 weeks’ gestation. Her BW was 2,000 g (−2.50 SD) length 45 cm (−1.83 SD), and OFC 30.5 cm (−2.39 SD). Neonatal course was uneventful. At 12 months of age, she experienced prolonged akinetic seizures with motor arrest and hypotonia, requiring phenobarbital therapy. At 10 years of age, she was nonverbal and microcephalic (OFC 46 cm, −4.90 SD). Physical examination showed dysmorphisms (Fig. 1a, panel c), truncal hypotonia, hand stereotypies, and scoliosis. Sleep EEG revealed low-voltage fast activity over polymicrogyric cortex.
Brain MRI studies revealed a similar malformative pattern in all patients (Fig. 1b), characterized by bilateral frontal and perysilvian polymicrogyria (Patients 1 and 3) and fronto-insular dysgyria (Patient 2), variable degrees of callosal hypo-dysgenesis, dysmorphic basal ganglia with indistinct anterior limbs of internal capsules, small olfactory bulbs, pontine and inferior vermis hypoplasia. White matter was globally reduced, especially at the level of ventral cingulum, with enlargement of lateral ventricles and peculiar temporal horn dilatation. Additional findings included an incidental solid mass in left cerebellar hemisphere in Patient 1 and trigonocephaly in Patient 2. DTI studies showed marked hypoplasia of the corpus callosum with prevalent posterior involvement, and reduced volume of the anterior limbs of the internal capsule and ventral cingulum. No aberrant course of the cortico-spinal tracts or other major white matter bundles was noted (Fig. 2 and Fig. 3 Supplementary Material).
Molecular findings
WES identified three distinct de novo heterozygous variants in DDX3X: c.1511G>A, p.(Gly504Glu) (Patient 1); c.1436_1439delinsTCTC, p.(Asp479Arg480delinsValSer) (Patient 2); c.641_643delTCA, p.(Ile214del)(Patient 3). Genomic coordinates refer to genome assembly hg19 and DDX3X nucleotide and amino acid variants to RefSeq entries NM_001356.3and NP_001347.3, respectively. The variants identified in patients 1 and 2 have not been previously reported in the literature or in any public database (including COSMIC, Decipher, Clinvar, and gnomAD). As to p.(Ile214del), the frameshift mutation affecting the same residue c.641_643delTCAinsCC, p.(Ile214Thrfs*7) has been reported as ‘pathogenic’ in ClinVar. All variants were interpreted according to the American College of Medical Genetics and Genomics (ACMG) guidelines and were classified as class 5 (Mutation Analysis Supplementary Material).
Discussion
As of September 2018, 53 patients harboring DDX3X variants have been collected in the Decipher database (Table 1). Most of the described variants are predicted to lead to loss of protein function (Table 2 Supplementary Material). Two large case series of 38 and 31 affected females harboring de novo or presumably de novo DDX3X variants have been reported by Snijders Blok L et al. and Wang et al., respectively [2, 7]. Conversely, only 10 male patients have been described so far, suggesting a lethal effect in most fetal cases [1, 7,8,9]. Six of them harbored maternally inherited missense variants, two carried de novo missense variants, and one carried a de novo splice site variant. The mode of inheritance in one patient harboring a missense variant was not confirmed [1, 7,8,9]. Regardless of gender, most DDX3X patients carry missense variants affecting the ATP-binding or C-terminal helicase domains, similarly to those identified in our patients, without a clear genotype-phenotype correlation [2, 7, 10].
DDX3X-related disorders are characterized by a relevant phenotypic variability (Table 1), especially with regard to the severity of neurodevelopmental involvement. While DD and/or ID are present in the vast majority of patients, a variable proportion of affected individuals show movement disorders, significant behavioral abnormalities, hypotonia, and seizures [2, 7]. Facial dysmorphisms are common, including clinical features of Toriello-Carey syndrome [2, 7, 8]. Interestingly, the existence of sex-specific phenotypes has been hypothesized [9]. Although all reported male patients have ID and global DD, and most of them suffer from movement disorders, they exhibit peculiar phenotypic features. In particular, male patients are rarely hypotonic as compared to females and often show craniofacial deformities (especially brachycephaly and plagiocephaly), ophthalmological abnormalities, and congenital heart disease [9].
In line with previous reports, our patients showed severe ID/DD, neurological abnormalities, microcephaly, and dysmorphic features (Table 3 Supplementary Material) [1]. Interestingly, occipital plagiocephaly was observed in Patient 1 and Patient 2 was diagnosed with sensorineural hearing loss and trigonocephaly. However, a true craniosynostosis was radiologically confirmed only in the latter (Fig. 3 Supplementary Material). While hearing impairment has been occasionally associated with DDX3X spectrum, trigonocephaly has not been reported in patients with DDX3X-related ID so far [2, 8].
All patients displayed a common brain malformative pattern characterized by bilateral frontal and perysilvian polymicrogyria and/or dysgyria, callosal hypo-dysgenesis, dysmorphic basal ganglia with indistinct anterior limbs of internal capsules, hypoplasia of the ventral cingulum, incomplete hippocampal rotation, and pontine and inferior vermis hypoplasia. Although abnormal neuroradiological findings have been described in several females harboring DDX3X variants, malformations of the cortical development have been only occasionally reported [1]. In particular, polymicrogyria was found in 4 out of 37 individuals of the largest case series published by Blok et al., and simplified gyral pattern was present in one patient reported by Dikow et al. [2, 8]. Remarkably, no mention of associated midbrain-hindbrain anomalies, basal ganglia dysmorphisms, and/or marked hypoplasia of ventral cingulum was made in these case series.
The neuroradiological features observed in our patients are similar, though less severe, to those reported in individuals with tubulinopathies or GRIN2B encephalopathy [11, 12]. These findings suggest that DDX3X might play a pivotal role not only in neuronal proliferation and migration, but also in white matter development. Despite the presence of callosal abnormalities and pontine hypoplasia, we did not find aberrant white matter bundles on DTI tractography, thus ruling out major axonal pathfinding defects. DDX3X is required for neurite outgrowth and dendritic spine formation through the translational activation of mRNAs involved in Rac1 activation [13,14,15]. This would lead to speculate about a common underlying pathogenic mechanism for brain malformations in tubulinopathies and DDX3X-related disorders, possibly involving altered microtubule stability and defective migration of cortical GABAergic interneurons due to impaired Rac1 signaling [14]. However, the presence of similar brain malformations in a minority of patients with DDX3X missense variants suggests a possible genotype-phenotype correlation requiring further validation in larger studies.
The incidental diagnosis of a cerebellar PA in Patient 1 represents another relevant aspect of this study. DDX3X plays a crucial role in cell cycle progression and is involved in Wnt/β-catenin signaling pathway and cancer dissemination [3, 5, 16]. Somatic DDX3X variants have been reported in several aggressive tumors, including Wnt-driven medulloblastoma [3, 5, 17]. Furthermore, a significant correlation between DDX3X expression and Snail levels has been reported in glioblastoma multiforme [16]. The missense variant p.(Gly504Glu) identified in Patient 1 affects the central helicase core of DDX3X (Fig. 1c, panel b), similarly to the somatic variants occurring in medulloblastoma. These variants impair DDX3X-RNA binding resulting in altered protein function, which might play a relevant role in ‘DDX3X-driven cancerogenesis’ [3]. Accordingly, we speculate that the p.(Gly504Glu) variant might have contributed to PA development as part of a more complex pathogenic mechanism. However, obvious limitations are worth noting. First, functional studies on PA tissues were not performed. Furthermore, we did not seek a second hit in DDX3X or further genes involved in tumorigenesis. Indeed, since DDX3X exerts tumor suppressing functions, patients harboring DDX3X germline variants might have a greater risk of developing somatic second hit mutations, leading to a possible increase in cancer risk [18].
In conclusion, this report expands the phenotypic spectrum of DDX3X-related disorders. Our findings suggest novel consequences of DDX3X variants, resulting in altered neuronal migration and proliferation, abnormal white matter development, and possible promotion of tumorigenesis. Accordingly, DDX3X should be considered in the differential diagnosis of tubulinopathies and included in the “cortical malformations” Next Generation Sequencing (NGS) panel. However, in the absence of pathognomonic clinical and radiological features due to the significant phenotypic variability, additional DDX3X cases are expected to be detected by WES alone in the next future. Further studies will help clarify the actual clinical relevance and implications of these observations.
References
Kellaris G, Khan K, Baig SM, Tsai IC, Zamora FM, Ruggieri P, et al. A hypomorphic inherited pathogenic variant in DDX3X causes male intellectual disability with additional neurodevelopmental and neurodegenerative features. Hum Genomics. 2018;12:11.
Snijders Blok L, Madsen E, Juusola J, Gilissen C, Baralle D, Reijnders MR, et al. Mutations in DDX3X are a common cause of unexplained intellectual disability with gender-specific effects on Wnt signaling. Am J Hum Genet. 2015;97:343–52.
Bol GM, Xie M, Raman V. DDX3, a potential target for cancer treatment. Mol Cancer. 2015;14:188.
Linder P, Lasko PF, Ashburner M, Leroy P, Nielsen PJ, Nishi K, et al. Birth of the D-E-A-D box. Nature. 1989;337:121–2.
Epling LB, Grace CR, Lowe BR, Partridge JF, Enemark EJ. Cancer-associated mutants of RNA helicase DDX3X are defective in RNA-stimulated ATP hydrolysis. J Mol Biol. 2015;427:1779–96.
Schröder M. Human DEAD-box protein 3 has multiple functions in gene regulation and cell cycle control and is a prime target for viral manipulation. Biochem Pharmacol. 2010;79:297–306.
Wang X, Posey JE, Rosenfeld JA, Bacino CA, Scaglia F, Immken L, et al. Phenotypic expansion in DDX3X—a common cause of intellectual disability in females. Ann Clin Transl Neurol. 2018;5:1277–85.
Dikow N, Granzow M, Graul-Neumann LM, Karch S, Hinderhofer K, Paramasivam N, et al. DDX3X mutations in two girls with a phenotype overlapping Toriello-Carey syndrome. Am J Med Genet A. 2017;173:1369–73.
Nicola P, Blackburn PR, Rasmussen KJ, Bertsch NL, Klee EW, Hasadsri L, et al. De novo DDX3X missense variants in males appear viable and contribute to syndromic intellectual disability. Am J Med Genet A. 2019 Feb 7. [Epub ahead of print]
Fieremans N, Van Esch H, Holvoet M, Van Goethem G, Devriendt K, Rosello M, et al. Identification of intellectual disability genes in female patients with a skewed X-inactivation pattern. Hum Mutat. 2016;37:804–11.
Mutch CA, Poduri A, Sahin M, Barry B, Walsh CA, Barkovich AJ. Disorders of microtubule function in neurons: imaging correlates. Am J Neuroradiol. 2016;37:528–35.
Platzer K, Yuan H, Schütz H, Winschel A, Chen W, Hu C, et al. GRIN2B encephalopathy: novel findings on phenotype, variant clustering, functional consequences and treatment aspects. J Med Genet. 2017;54:460–70.
Chen HH, Yu HI, Tarn WY. DDX3 modulates neurite development via translationally activating an RNA regulon involved in Rac1 activation. J Neurosci. 2016;36:9792–804.
Tivodar S, Kalemaki K, Kounoupa Z, Vidaki M, Theodorakis K, Denaxa M, et al. Rac-GTPases regulate microtubule stability and axon growth of cortical GABAergic interneurons. Cereb Cortex. 2015;25:2370–82.
Zamboni V, Armentano M, Berto G, Ciraolo E, Ghigo A, Garzotto D, et al. Hyperactivity of Rac1-GTPase pathway impairs neuritogenesis of cortical neurons by altering actin dynamics. Sci Rep. 2018;8:7254.
Sun M, Song L, Zhou T, Gillespie GY, Jope RS. The role of DDX3 in regulating Snail. Biochim Biophys Acta. 2011;1813:438–47.
Pugh TJ, Weeraratne SD, Archer TC, Pomeranz Krummel DA, Auclair D, Bochicchio J, et al. Medulloblastoma exome sequencing uncovers subtype-specific somatic mutations. Nature. 2012;488:106–10.
Dunford A, Weinstock DM, Savova V, Schumacher SE, Cleary JP, Yoda A, et al. Tumor-suppressor genes that escape from X-inactivation contribute to cancer sex bias. Nat Genet. 2017;49:10–6.
Acknowledgements
We thank the patients and their families for the collaboration and the consent to the publication of this paper.
Author information
Authors and Affiliations
Consortia
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This study was approved by the Ethical Committees of IRCCS Istituto Giannina Gaslini and Federico II University.
Informed consent
Written informed consent was obtained from the parents of the subjects of the study.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Telethon Undiagnosed Diseases Program (TUDP) consortium
Supplementary information
Rights and permissions
About this article

Cite this article
Scala, M., Torella, A., Severino, M. et al. Three de novo DDX3X variants associated with distinctive brain developmental abnormalities and brain tumor in intellectually disabled females. Eur J Hum Genet 27, 1254–1259 (2019). https://doi.org/10.1038/s41431-019-0392-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41431-019-0392-7
This article is cited by
-
Social and emotional characteristics of girls and young women with DDX3X-associated intellectual disability: a descriptive and comparative study
Journal of Autism and Developmental Disorders (2023)
-
An approach for brain tumour detection based on dual-tree complex Gabor wavelet transform and neural network using Hadoop big data analysis
Multimedia Tools and Applications (2022)
-
Diagnostic Approach to Cerebellar Hypoplasia
The Cerebellum (2021)
