
Abstract
KR-31543, (2S, 3R, 4S)-6-amino-4-[N-(4-chlorophenyl)-N-(2-methyl-2H-tetrazol-5-ylmethyl) amino]-3,4-dihydro-2-dimethyoxymethyl-3-hydroxy-2-methyl-2H-1-benz opyran is a new neuroprotective agent for ischemia-reperfusion damage. It has also been reported that KR-31543 has protective effects on lipid peroxidation and H2O2-induced reactive oxygen species production. In this study, we investigated the anti-inflammatory and anti-atherogenic properties of KR-31543. We observed that KR-31543 treatment reduced the production of MCP-1, IL-8, and VCAM-1 in HUVECs, and of MCP-1 and IL-6 in THP-1 human monocytes. We also examined the effect of KR-31543 on monocytes migration in vitro. KR-31543 treatment effectively reduced the migration of THP-1 human monocytes to the HUVEC monolayer in a dose-dependent manner. We next examined the effects of this compound on atherogenesis in LDL receptor deficient (Ldlr-/-) mice. After 10 weeks of western diet, the formation of atherosclerotic lesion in aorta was reduced in the KR-31543-treated group compared to the control group. The accumulation of macrophages in lesion was also reduced in KR-31543 treated group. However, the plasma levels of total cholesterol, HDL, LDL, and triglyceride were not affected by KR-31543 treatment. Taken together, these results show that KR-31543 has anti-inflammatory properties on human monocytes and endothelial cells, and inhibits fatty streak lesion formation in mouse model of atherosclerosis, suggesting the potential of KR-31543 for the treatment for atherosclerosis.
Similar content being viewed by others

Introduction
Atherosclerosis is a representative chronic vascular inflammatory disease. The early stages of the disease involve the recruitment of monocytes and lymphocytes into the arterial intima, and localized accumulation of the cells (Ross, 1999). This early inflammatory step of atherosclerosis is mostly initiated by reactive oxygen species (ROS)-induced oxidative modification of lipoproteins in subintimal space (Hansson and Hermansson, 2011). Oxidized low-density lipoprotein (LDL) particularly induces the expression of inflammatory molecules, including vascular cell adhesion moledule-1, intercellular adhesion molecule-1, interleukin-8 (IL-8), macrophage chemoattractant protein-1 (MCP-1), etc. (Terkeltaub et al., 1994; Khan et al., 1995; Claise et al., 1996; Wang et al., 1997). These multiple processes influence the recruitment of monocytes to the intima of arterial wall. Therefore, the blockage of oxidative modification of LDL is an important strategy for reducing early stage of atherosclerosis.
(2S, 3R, 4S)-6-amino-4-[4-chloro-(2-methyltetrazol-5-ylmethylphenyl) amino]-3,4-dihydro-3-hydroxy-2-dimethoxy-methyl-2H-1-benzopyran, or KR-31543, synthesized by the Korea Research Institute of Chemical Technology, was originally developed as a neuroprotective agent for ischemia and reperfusion damage. Possessing both antioxidant and potassium channel-modulating properties, KR-31543 protected cultured rat cortex neurons against iron-induced oxidative injury in vitro, and significantly reduced infarct volume following occlusion in the rat model of transient cerebral ischemia (Kim et al., 2002; Lee et al., 2003). Since the inhibition of ROS generation can attenuate the generation of atherosclerotic lesions (Park and Oh, 2011), the anti-oxidative activity of KR-31543 led us to investigate its anti-atherogenic activity. Here we first demonstrated the anti-inflammatory and anti-atherosclerotic potential of KR-31543 using THP-1 human monocytes, HUVECs, and LDL receptor deficient (Ldlr-/-) mice.
Results
KR-31543 down-regulated the production of proinflammatory molecules in HUVECs and THP-1 monocytes
In a previous paper, KR-31378, an antioxidant and potassium channel modulator similar to KR-31543, has been reported to reduce the production of inflammatory molecules in cytokine-stimulated leukocytes and endothelial cells directly (Kim et al., 2004). In this study, the expression levels of two chemotactic molecules, MCP-1 and IL-8, were determined in HUVECs treated and untreated with several indicated concentrations of KR-31543, having no cytotoxicity in mitochondrial tetrazolium assay. Incubation of HUVECs with KR-31543 for 2 h prior to TNF-α stimulation resulted in a significant decrease in the secretion of MCP-1 in a dose-dependent manner (Figure 1B). Moreover, KR-31543 markedly decreased the production of IL-8 in HUVECs (Figure 1C). VCAM-1 expression was also decreased at high concentration of KR-31543, but there was no change in lower concentration (Figure 1D). It was also tested whether KR-31543 decreased cytokine production in macrophages, and found that KR-31543 effectively reduced MCP-1 and IL-6 productions in THP-1 human monocyte (Figures 1E and 1F).

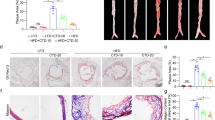
Effect of KR-31543 on the expressions of proinflammatory molecules in human endothelial cells and THP-1 monocytes. (A) The chemical structure of KR-31543. (B-D) The effect of KR-31543 treatment on several proinflammatory molecules including (B) MCP-1 and (C) IL-8 and (D) VCAM-1 in HUVECs. (E, F) The effect of KR-31543 treatment on the expression of (E) MCP-1 and (F) IL-6 in THP-1 human monocytes. *indicates P < 0.01 compared to control group.
KR-31543 reduced migration of macrophage induced by TNF-α
In this study, in vitro leukocyte migration and adhesion assays were performed to see the effects of KR-31543 on early inflammatory process. First, it was investigated whether KR-31543 reduces the adhesion of monocytes to endothelial cells. We found that although 10-6 M of KR-31543 KR-31543 did not affect the expression of VCAM-1 (Figure 1D), KR-31543 effectively reduced the attachment of THP-1 human monocytes to HUVEC monolayers at this concentration (Figure 2A). It seems that attenuated expressions of IL-8 and MCP-1 by 10-6 M of KR-31543 are responsible for reduced monocyte attachment. The observation that KR-31543 reduced THP-1 human monocyte adhesion to HUVEC monolayers prompted an examination of whether KR-31543 could also affect cell migration. HUVECs were pretreated with KR-31543 for 2 h, then stimulated with TNF-α for 12 h; stimulated with TNF-α alone; or left untreated. The conditioned culture media were then placed in the bottom chambers of a ChemoTx plate. THP-1 human monocytes were loaded onto the 5-µm pore filter and incubated for 6 h. Incubation with TNF-α-conditioned medium resulted in approximately 6-fold increase of migrated THP-1 human monocytes compared to the untreated HUVEC conditioned medium. In contrast, a conditioned medium taken from cells pre-incubated with KR-31543 markedly reduced the migration of THP-1 human monocytes in a dose-dependent manner (Figure 2B).

The effect of KR-31543 on monocyte adhesion and transmigration. (A) Monocyte adhesion assay. HUVECs were pre-incubated with KR-31543 (10-6 and 10-7 M) for 2 h, and then stimulated with TNF-α for 12 h. Next THP-1 human monocytes were added to the culture. After 20 min, unbound cells were washed and the remaining attached cells were counted in five randomly selected optical fields per well. (B) Monocyte migration assay. THP-1 human monocytes were loaded into the upper chamber of Transwell plates and the lower chambers were filled with conditioned medium from HUVECs treated with KR-31543 (10-6 or 10-7 M) or left untreated for 2 h and subsequently activated with TNF-α for 12 h. *indicates P < 0.05 compared to TNF-α stimulated cells without KR-31543 treatment.
KR-31543 significantly reduced the atherosclerotic lesion and lesional macrophage accumulation in atherogenic diet-fed Ldlr-/- mice
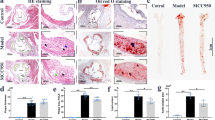
To analyze the pharmacokinetics of KR-31543, the mice were administered 3 mg of KR-31543 once orally. This dose was equivalent to the daily amount ingested from the diet used in mouse study (KR-31543 1% w/w). The plasma KR-31543 level in the treated animals was 1.0 × 10-7 M at 24 h after giving the chemical. Therefore, considering that KR-31543 was effective to reduce inflammatory cytokines from endothelial cells and macrophages at the concentration of 1 × 10-7 M, it seems that the administrated KR-31543 effectively attenuate the vascular inflammatory processes. To determine the effect of KR-31543 on the development of atherosclerosis, atherosclerotic lesions in aortic vessels from the control and KR-31543 groups which had been fed a Western diet for 10 weeks were analyzed. Necropsy and histopathological examinations did not reveal notable lesions in the parenchymal organs of both groups of mice (data not shown). In addition, daily food intake and weight gain was similar between the two groups (data not shown). Measurement of the lesion size by computer-associated morphometry revealed that the mean lesion area of the aortic sinus in the KR-31543 group was smaller than that of the control group by around 40% (Figure 3A). The results from cell study prompted a measurement of the macrophage accumulation in atherosclerotic lesion. The lesions were immunestained with anti-macrophage antibodies (MOMA-2) and measured the macrophage positive area in the aortic sinus to determine whether KR-31543 treatment reduced monocyte recruitment to the lesion. The macrophage positive area was reduced significantly in the KR-31543 group compared to the control group (Figure 3B).

The anti-atherogenic potential of KR-31543 in Ldlr-/- mice. (A) Representative figures of Oil red O-positive atherosclerotic lesion from Control (n = 8) and KR-31543 groups (n = 8). The mean lesion size was significantly reduced in KR-31543 group compared to control group. (B) Representative immunohistochemical staining of macrophages in control (n = 8) and KR-31543-treated (n = 8) mice are shown. KR-31543 effectively reduced the macrophage accumulation in the lesion. The graph shows the percent mean area of macrophage accumulation in total atherosclerotic lesion of each mouse. *indicates P < 0.05 compared to control group.
KR-31543 did not affect plasma lipid levels
Since lipid lowering agent also can reduce the formation of atherosclerotic lesion, the plasma lipid profiles were analyzed in KR-31543 treated mice. KR-31543 supplementation did not affect plasma concentrations of total cholesterol, HDL-cholesterol, LDL-cholesterol and triglyceride (Figure 4). These results suggest that KR-31543 reduces atherosclerosis not by modulating plasma lipid profiles, but by its anti-inflammatory properties.

The effect of KR-31543 on plasma lipid profiles. (A) Total cholesterol, (B) Triglyceride, (C) HDL-cholesterol, and (D) LDL-cholesterol were measured by blood chemical analyzer. There was no significant difference between groups.
Taken together, our results suggest that KR-31543 reduces the atherosclerotic lesion formation possibly by disturbing the inflammatory processes leading to attenuation of atherosclerosis (Figure 5).

Suggested possible mechanism for the anti-atherogenic activity of KR-31543. KR-31543 attenuates the atherosclerotic lesion formation by reducing the production of cytokines and disturbing the adhesion and transmigration of monocytes (red line). The anti-oxidative activity of KR-31543 may also reduce the oxidation of LDL, which can be an additional possible mechanism of its antiatherogenic characteristic (dashed red line).
Discussion
Many previous studies have suggested that anti-oxidants inhibit inflammatory responses by blocking the expression of pro-inflammatory molecules, leading to attenuation of the atherosclerotic lesion formation (Sugamura and Keaney, 2011). In this study, we demonstrated that the antioxidant KR-31543 decreases diet-induced lesion formation and macrophage infiltration in Ldlr-/- mice. Treatment with KR-31543 decreases monocyte migration and attachment to endothelial cells in vitro, which may be attributed to a reduction in the production of IL-8, IL-6 and MCP-1.
Hyperlipidemia is one of the major risk factors for atherosclerosis, and many studies have suggested that lipid-lowering drugs are effective to reduce atherosclerosis (Ross and Harker, 1976; Gotto and LaRosa, 2005; Hackam, 2006; Ford et al., 2007). However, our analysis of the plasma level of total cholesterol, HDL, LDL, and triglycerides revealed that treatment with KR-31543 did not influence the lipid profiles of the mice fed high fat diet. Thus, it was concluded that unlike the lipid profile-modulating drugs like statins, KR-31543 exerts its anti-atherogenic effect through mechanisms other than improvement of lipid profiles.
In this study, our cell adhesion assay demonstrated that KR-31543 efficiently reduced THP-1 monocyte attachment to endothelial cells stimulated by TNF-α without attendant cytotoxicity. These results led us to investigate the effect of KR-31543 on the transmigration of monocytic cells. In our migration assays, we found that the conditioned media from HUVEC cells challenged with TNF-α significantly increased THP-1 migration, whereas pretreatment with KR-31543 decreased the number of migrating cells in a dose-dependent manner. Taken together, these results suggest that KR-31543 may block the monocyte adhesion and transmigration induced by the factors derived from endothelial cells and monocytes. Consistent with this data, KR-31543 effectively reduced TNF-α-induced production of chemo-attracting molecules in HUVECs. The secretion of IL-8, a well-known regulator of neutrophil migration and a mediator of acute inflammatory reactions, was significantly reduced. Furthermore, the production of MCP-1, a mediator in chronic inflammation was decreased by pre-incubation with KR-31543 not only in HUVECs, but also in THP-1 human monocytes (Scapini et al., 2000; Ito and Ikeda, 2003). Therefore, we speculate that KR-31543 acts as a regulator of both endothelial cell and monocyte activity by down-regulation of monocyte chemo-attractant production, resulting in a synergistic suppression of recruitment in vivo. This hypothesis is supported by the observation that in the lesions formed in Ldlr-/- mice, macrophage accumulation was reduced in the KR31543-treated group as compared to the control group.
In this study, it is not clarified how KR-31543 exerts an inhibitory effect on monocyte adhesion on endothelial cell monolayer and transmigration. Previously, it has been well-known that hydrogen peroxide and oxygen radicals are produced during inflammatory processes and play second messengers directly or indirectly mediating the activation of pro-inflammatory nuclear transcription factors, such as NF-κB (Schreck et al., 1991). Therefore, the reduction of reactive oxygen species (ROS) by anti-oxidants, such as flavonoid, α-tocopherol, ascorbate, and gallate, has potential anti-atherogenic effects via decreased pro-inflammatory molecules needed for monocyte migration and adhesion (Gerritsen et al., 1995; Murase et al., 1999; Carr et al., 2000). MCP-1 and IL-8, which play roles in the recruitment of monocytes, are regulated via the TNF-α-mediated NF-κB signaling pathway (Collins and Cybulsky, 2001; Wajant et al., 2003). Thus, we speculate that the mechanism of the anti-atherogenic activity of KR-31543 involves antioxidant action leading to a reduction in the activation of pro-inflammatory transcriptional factors such as NF-κB.
In conclusion, we showed that KR-31543 effectively reduced atherosclerotic lesion formation and macrophage accumulation in the lesions via its anti-inflammatory properties, which modulate multiple cellular activities and interactions. Our results indicate that in addition to its neuro-protective characteristics, KR-31543 may be a valid therapeutic candidate for the treatment of atherosclerosis.
Methods
Cell culture
Human umbilical vein endothelial cells (HUVECs) were purchased from Clonetics (BioWhittaker, Walkersville, MD) and maintained in EGM-2 Bullet kit media (BioWhittaker) at 37℃ with 5% CO2. All HUVECs used in this study were between passages 3 and 7.
THP-1 human monocytes were obtained from the American Type Culture Collection. THP-1 human monocytes were cultured in RPMI1640 containing 10% Fetal Bovine Serum supplemented with 100 units/ml of penicillin and 100 µg/ml of streptomycin (Gibco BRL, CA).
Chemicals
KR-31543 (Figure 1A, generously donated by the Korea Research Institute of Chemical Technology, Daejon, Korea) was dissolved in dimethyl sulfoxide as a 100 mM stock solution and kept at -20℃.
Animal model
Ldlr-/- mice were randomly divided into two groups (contol n = 8, KR-31543 group n = 8). The control group was fed a Western diet (CRF-1 supplemented with 0.15% cholesterol and 20% fat, Oriental Yeast Co. Ltd., Tokyo, Japan), while the KR-31543 group was fed the same diet supplemented with 0.1% (w/w) KR-31543. All mice were given water and food ad libitum. After 10 weeks, the mice were killed, and plasma chemical profiles were generated using an automatic blood chemical analyzer (HITACHI, Japan). Parenchymal organs were also harvested for histopathological examination. This study was performed in accordance with the recommendations from the Declaration of Helsinki. All animal study protocols were approved by the Institutional Animal Care and Usage Committee of Ewha Womans University (Seoul, Korea).
Assessment of atherosclerosis
Fatty streak lesions were quantified by evaluating the lesion size in the aortic sinus, as previously described (Choi et al., 2003). The heart and aorta were perfused with phosphate-buffered saline (PBS) for 10 min, with 4% paraformaldehyde for 5 min, and then promptly removed. After fixation for 1 day in 10% buffered neutral formalin, the hearts and aortas were embedded in OCT medium (Life Science International, UK) and frozen at -70℃. All samples were sectioned using a cryostat at 20℃, and six consecutive 9 µm-thick sections were cut from the aorta where the valve cusp becomes visible. Atherosclerotic plaques were stained with Oil red O and counter-stained with Harris hematoxylin. The lesion area was then quantified by computer-assisted morphometry (Image pro plus, MD) and the average lesion size was calculated for each animal.
Measurement of macrophage accumulation in atherosclerotic lesions
Frozen sections were stained with a rat monoclonal antibody against mouse macrophages (MOMA-2, Serotec Inc., NC) using the ABC method (Novastatin Super ABC kit, Novocastra Lab. Ltd., UK). The macrophage contents of lesions was calculated as the percentage of mean MOMA-2 positive area versus mean total lesion area using five consecutive sections per mouse by computer-aided morphometric analysis.
Macrophage adhesion assay
Adhesion of THP-1 human monocytes to HUVEC monolayer was assayed as described previously (Che et al., 2002; Kim et al., 2004). HUVECs were plated on 6-well plates at a density of 1 × 105 cells per well and cultured to 90% confluence in EGM-2 Bullet kit media. After 2 h of KR-31543 treatment, the cells were incubated with 20 ng/ml of TNF-α for another 12 h. THP-1 human monocytes in the exponential growth phase were washed with serum-free DMEM and then re-suspended in the same medium to 2 × 104 cells/ml. The THP-1 human monocyte suspension (1 ml) was added into the wells containing HUVEC monolayers and incubated for 20 min at 37℃. Unbound cells were washed three times with serum-free DMEM. The total number of adherent cells was counted in five randomly selected optical fields per well.
Monocyte migration assay
Monocyte migration assay was performed as described previously (Kim et al., 2004). Briefly, the migration of THP-1 human monocytes was assayed using the ChemoTx kit (Neuro Probe, Inc. MD) in a 96-well plate housing a collagen-precoated polycarbonate filter with 5.0-µm pores, according to the manufacturer's instructions. The lower chambers were filled with 30 µl of medium obtained from cultured human umbilical vein endothelial cells (HUVECs), treated with KR-31543 for 2 h, and subsequently stimulated with TNF-α (Promega). The filter membranes were loaded with 1.25 × 104 cells in 25 µl. After incubation for 6 h at 37℃, cells that had migrated into the lower chamber were counted by flow cytometry.
Measurement of proinflammatory molecules
THP-1 human monocytes were differentiated into macrophages by PMA treatment for 24 h. Then LPS and indicated doses of KR-31543 were added to culture media. After pre-incubation with the indicated dose of KR-31543, HUVECs were stimulated by 3 ng/ml of TNF-α. The levels of IL-8 and MCP-1 were determined by ELISA method (R&D) according to the manufacturer's protocol. The measurement of VCAM-1 expression was performed as described previously (Kim et al., 2004). Briefly, HUVECs were treated with KR-31543 for 2 h and then stimulated with 1 ng/ml of TNF-α for 12 h. The cells were collected and stained for 1 h on ice with mouse anti-human VCAM-1 monoclonal antibody (PharMingen) or mouse IgG1κ (PharMingen) as an isotype control. After this, cells were washed and then analyzed by FACScaliber (BD).
Statistical analysis
Comparisons between two groups were made using a two-tailed Student's t test. ANOVA followed by the Fisher's Protected Least Significant Difference method was used for multiple comparisons. All data in this study are expressed as the mean ± standard deviation. Values of P < 0.05 were considered significant.
Abbreviations
- HUVEC:
-
human umbilical vein endothelial cell
- MCP-1:
-
monocyte chemotatic protein-1
- ROS:
-
reactive oxygen species
References
Carr AC, Zhu BZ, Frei B . Potential antiatherogenic mechanisms of ascorbate (vitamin C) and alpha-tocopherol (vitamin E) . Circ Res 2000 ; 87 : 349 - 354
Che W, Lerner-Marmarosh N, Huang Q, Osawa M, Ohta S, Yoshizumi M, Glassman M, Lee JD, Yan C, Berk BC, Abe J . Insulin-like growth factor-1 enhances inflammatory responses in endothelial cells: role of Gab1 and MEKK3 in TNF-alpha-induced c-Jun and NF-kappaB activation and adhesion molecule expression . Circ Res 2002 ; 90 : 1222 - 1230
Choi JH, Jeong TS, Kim DY, Kim YM, Na HJ, Nam KH, Lee SB, Kim HC, Oh SR, Choi YK, Bok SH, Oh GT . Hematein inhibits atherosclerosis by inhibition of reactive oxygen generation and NF-kappaB-dependent inflammatory mediators in hyperlipidemic mice . J Cardiovasc Pharmacol 2003 ; 42 : 287 - 295
Claise C, Edeas M, Chalas J, Cockx A, Abella A, Capel L, Lindenbaum A . Oxidized low-density lipoprotein induces the production of interleukin-8 by endothelial cells . FEBS Lett 1996 ; 398 : 223 - 227
Collins T, Cybulsky MI . NF-kappaB: pivotal mediator or innocent bystander in atherogenesis ? J Clin Invest 2001 ; 107 : 255 - 264
Ford ES, Ajani UA, Croft JB, Critchley JA, Labarthe DR, Kottke TE, Giles WH, Capewell S . Explaining the decrease in U.S. deaths from coronary disease, 1980-2000 . N Engl J Med 2007 ; 356 : 2388 - 2398
Gerritsen ME, Carley WW, Ranges GE, Shen CP, Phan SA, Ligon GF, Perry CA . Flavonoids inhibit cytokine-induced endothelial cell adhesion protein gene expression . Am J Pathol 1995 ; 147 : 278 - 292
Gotto AM, LaRosa JC . The benefits of statin therapy: what questions remain ? Clin Cardiol 2005 ; 28 : 499 - 503
Hackam DG . Intensive reduction of low-density lipoprotein-cholesterol: implications of recent trials . Am J Cardiovasc Drugs 2006 ; 6 : 367 - 371
Hansson GK, Hermansson A . The immune system in atherosclerosis . Nat Immunol 2011 ; 12 : 204 - 212
Ito T, Ikeda U . Inflammatory cytokines and cardiovascular disease . Curr Drug Targets Inflamm Allergy 2003 ; 2 : 257 - 265
Khan BV, Parthasarathy SS, Alexander RW, Medford RM . Modified low density lipoprotein and its constituents augment cytokine-activated vascular cell adhesion molecule-1 gene expression in human vascular endothelial cells . J Clin Invest 1995 ; 95 : 1262 - 1270
Kim J, Ji HY, Lee SS, Yoo SE, Kim SO, Lee DH, Lim H, Lee HS . Metabolism of a new neuroprotective agent for ischemia-reperfusion damage, KR-31543 in the rat using liquid chromatography/electrospray mass spectrometry . Arch Pharm Res 2002 ; 25 : 664 - 668
Kim J, Nam KH, Kim SO, Choi JH, Kim HC, Yang SD, Kang JH, Ryu YH, Oh GT, Yoo SE . KR-31378 ameliorates atherosclerosis by blocking monocyte recruitment in hypercholestrolemic mice . FASEB J 2004 ; 18 : 714 - 716
Lee MH, Bae SK, Kim EJ, Kim YG, Kim SO, Lee DH, Lim H, Yoo SE, Lee MG . Dose-independent pharmacokinetics of a new neuroprotective agent for ischemia-reperfusion damage, KR-31543, after intravenous and oral administration to rats: hepatic and intestinal first-pass effects . J Pharm Sci 2003 ; 92 : 190 - 201
Murase T, Kume N, Hase T, Shibuya Y, Nishizawa Y, Tokimitsu I, Kita T . Gallates inhibit cytokine-induced nuclear translocation of NF-kappaB and expression of leukocyte adhesion molecules in vascular endothelial cells . Arterioscler Thromb Vasc Biol 1999 ; 19 : 1412 - 1420
Park JG, Oh GT . The role of peroxidases in the pathogenesis of atherosclerosis . BMB Rep 2011 ; 44 : 497 - 505
Ross R, Harker L . Hyperlipidemia and atherosclerosis . Science 1976 ; 193 : 1094 - 1100
Ross R . Atherosclerosis--an inflammatory disease . N Engl J Med 1999 ; 340 : 115 - 126
Scapini P, Lapinet-Vera JA, Gasperini S, Calzetti F, Bazzoni F, Cassatella MA . The neutrophil as a cellular source of chemokines . Immunol Rev 2000 ; 177 : 195 - 203
Schreck R, Rieber P, Baeuerle PA . Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-kappa B transcription factor and HIV-1 . EMBO J 1991 ; 10 : 2247 - 2258
Sugamura K, Keaney JF . Reactive oxygen species in cardiovascular disease . Free Radic Biol Med 2011 ; 51 : 978 - 992
Terkeltaub R, Banka CL, Solan J, Santoro D, Brand K, Curtiss LK . Oxidized LDL induces monocytic cell expression of interleukin-8, a chemokine with T-lymphocyte chemotactic activity . Arterioscler Thromb 1994 ; 14 : 47 - 53
Wajant H, Pfizenmaier K, Scheurich P . Tumor necrosis factor signaling . Cell Death Differ 2003 ; 10 : 45 - 65
Wang GP, Deng ZD, Ni J, Qu ZL . Oxidized low density lipoprotein and very low density lipoprotein enhance expression of monocyte chemoattractant protein-1 in rabbit peritoneal exudate macrophages . Atherosclerosis 1997 ; 133 : 31 - 36
Acknowledgements
We author appreciate Dr. Jiyun Kim for the supporting on this study. This work was support by a National Core Research Center glant (R15-2006-020), and the National Research Foundation (20100019542) from the Ministry of Education, Science & Technology, South Korea. This work was also supported by a grant from Basic Science Research Program (2011-0013669) through the National Research Foundation funded by the Ministry of Education, Science & Technology, South Korea.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Choi, JH., Yoo, JY., Kim, SO. et al. KR-31543 reduces the production of proinflammatory molecules in human endothelial cells and monocytes and attenuates atherosclerosis in mouse model. Exp Mol Med 44, 733–739 (2012). https://doi.org/10.3858/emm.2012.44.12.081
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3858/emm.2012.44.12.081
