
Abstract
Penguins use twice as much metabolic energy as other terrestrial animals of a similar mass to walk a given distance1,2, which was thought to be because side-to-side waddling requires excessive work. Here we show that waddling actually conserves mechanical energy and suggest instead that walking is expensive for penguins because their short legs require them to generate muscular force rapidly.
Similar content being viewed by others


Main
Many animals reduce the muscular work of walking by exchanging the gravitational potential energy and kinetic energy of the centre of mass, like an inverted pendulum3. But penguins' waddling gait appears to involve large fluctuations in lateral kinetic energy and a poor exchange of mechanical energy1. To test the idea that penguin walking is expensive because it requires more mechanical work to lift and accelerate their body than other animals of a similar mass, we used a force platform to measure the vertical, fore–aft and lateral ground-reaction forces of five walking emperor penguins (Aptenodytes forsteri). Forces were integrated to calculate the centre-of-mass velocities, displacements, and fluctuations in gravitational potential energy and kinetic energy4.
The fluctuations in lateral velocity were nearly three times those in the fore–aft direction. Lateral-velocity fluctuations occurred about zero, whereas the fore–aft velocity fluctuated about the average walking speed. Thus, kinetic energy fluctuates much less laterally than in the fore–aft direction. At moderate walking speeds of about 0.5 m s− 1, fluctuations in lateral kinetic energy accounted for about 30% of the total kinetic-energy fluctuations — a large value compared with other terrestrial vertebrates5,6.
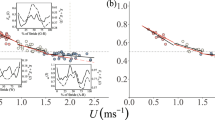
Penguins recovered up to 80% of the mechanical energy during each stride, because of the similar amplitudes and out-of-phase fluctuations in gravitational potential and kinetic energies (Fig. 1a–c). This value is among the highest reported for any animal, indicating a large reduction in the mechanical work done by the penguin's muscular system ( Fig. 1d). Recovery of mechanical energy was greatest and the work performed on the centre of mass per distance walked was least at about 0.5 m s− 1, a speed commonly used in nature2.

a,b, Fluctuations in gravitational potential and kinetic energy of the centre of mass for an emperor penguin walking at 0.54 m s− 1. Bars, times when left or right foot was in contact with the ground. J, joules. c,d, Inverted pendulum recovery of mechanical energy ( c) and work (W) per unit distance at a range of walking speeds (d). Dashed lines in b–d indicate the effect of excluding lateral kinetic energy from the analysis. e, Rate of generating muscular force, as indicated by the inverse of ground-contact time (1/tc), for emperor penguin, guinea fowl (Numida meleagris)10 and rhea (Rhea americana) (P. G. Weyand, unpublished results), and, f, standing hip height11 (leg length). The metabolic costs of transport of emperor penguins (8.64 J kg− 1 m− 1) and guinea fowl (8.38 J kg− 1 m− 1) are twice those of rheas (4.23 J kg− 1 m− 1) (ref. 8).
Surprisingly, excluding lateral kinetic energy — that is, waddling — from our calculations would result in the recovery of less mechanical energy and more work being required from the muscles (Fig. 1b–d). Lateral movements increase the kinetic energy available to convert into gravitational potential energy and also cause these energies to fluctuate more completely out of phase.
Other components of work, such as rotating the body about the centre of mass and swinging the limbs, probably do not account for the high cost: rotational work is approximately 10% of the total centre-of-mass work, and penguins have comparatively light, short limbs which offset their quick strides. At their preferred speed, emperor penguins perform the same amount of mechanical work to lift and accelerate their centre of mass as other walking animals of the same mass. Waddling therefore does not explain the high metabolic cost of penguin walking.
We propose instead that this high cost is due to rapid force generation. Regardless of the work muscles perform, they must actively generate force to support bodyweight, and this cost is proportional to the magnitude and rate of generating force7,8. At the same walking speed, emperor penguins and rheas — a flightless bird of a similar mass — generate the same average force on the ground. But because of their comparatively short legs, emperors generate this force twice as fast (Fig. 1e, f), requiring faster, less economical muscle fibres9. Our results indicate that it is their short legs, and not their waddling gait, that accounts for the high cost of penguin walking.
References
Pinshow, B., Fedak, M. A. & Schmidt-Nielsen, K. Science 195, 592– 594 (1977).
Dewasmes, G., Le Maho, Y., Cornet, A. & Groscolas, R. J. Appl. Physiol. 49, 888–896 ( 1980).
Cavagna, G. A., Heglund, N. C. & Taylor, C. R. Am. J. Physiol. 233, R243– R261 (1977).
Cavagna, G. A. J. Appl. Physiol. 39, 174–179 (1975).
Farley, C. T. & Ko, T. C. J. Exp. Biol. 200, 2177–2188 (1997).
Cavagna, G. A., Saibene, F. P. & Margaria, R. J. Appl. Physiol. 18, 1– 9 (1963).
Kram, R. & Taylor, C. R. Nature 346, 265–267 (1990).
Roberts, T. J., Kram, R., Weyand, P. G. & Taylor, C. R. J. Exp. Biol. 201, 2745–2751 ( 1998).
Rall, J. A. Exercise Sport Sci. Rev. 13, 33–74 (1985).
Gatesy, S. M. J. Morphol. 240, 115–125 (1999).
Gatesy, S. M. & Biewener, A. A. J. Zool. 224, 127–147 (1991).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Griffin, T., Kram, R. Penguin waddling is not wasteful. Nature 408, 929 (2000). https://doi.org/10.1038/35050167
Issue Date:
DOI: https://doi.org/10.1038/35050167
This article is cited by
-
Terrestrial locomotion energy costs vary considerably between species: no evidence that this is explained by rate of leg force production or ecology
Scientific Reports (2019)
-
Effort reduction in articulation in sign languages and dance
Journal of Cultural Cognitive Science (2019)
-
Neither season nor sex affects the cost of terrestrial locomotion in a circumpolar diving duck: the common eider (Somateria mollissima)
Polar Biology (2014)
-
Evolutionary Developmental Biology and Human Language Evolution: Constraints on Adaptation
Evolutionary Biology (2012)
-
Inefficient use of inverted pendulum mechanism during quadrupedal walking in the Japanese macaque
Primates (2012)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
