


















« Prev Next »

The ability to communicate effectively with other individuals plays a critical role in the lives of all animals. Whether we are examining how moths attract a mate, ground squirrels convey information about nearby predators, or chimpanzees maintain positions in a dominance hierarchy, communication systems are involved. Here, I provide a primer about the types of communication signals used by animals and the variety of functions they serve.
Animal communication is classically defined as occurring when “...the action of or cue given by one organism [the sender] is perceived by and thus alters the probability pattern of behavior in another organism [the receiver] in a fashion adaptive to either one both of the participants” (Wilson 1975). While both a sender and receiver must be involved for communication to occur (Figure 1), in some cases only one player benefits from the interaction. For example, female Photuris fireflies manipulate smaller, male Photinus fireflies by mimicking the flash signals produced by Photinus females. When males investigate the signal, they are voraciously consumed by the larger firefly (Lloyd 1975; Figure 2). This is clearly a case where the sender benefits and the receiver does not. Alternatively, in the case of fringe-lipped bats, Trachops cirrhosus, and tungara frogs, Physalaemus pustulosus, the receiver is the only player that benefits from the interaction. Male tungara frogs produce advertisement calls to attract females to their location; while the signal is designed to be received by females, eavesdropping fringe-lipped bats also detect the calls, and use that information to locate and capture frogs (Ryan et al. 1982). Despite these examples, there are many cases in which both the sender and receiver benefit from exchanging information. Greater sage grouse nicely illustrate such “true communication”; during the mating season, males produce strutting displays that are energetically expensive, and females use this honest information about male quality to choose which individuals to mate with (Vehrencamp et al. 1989).
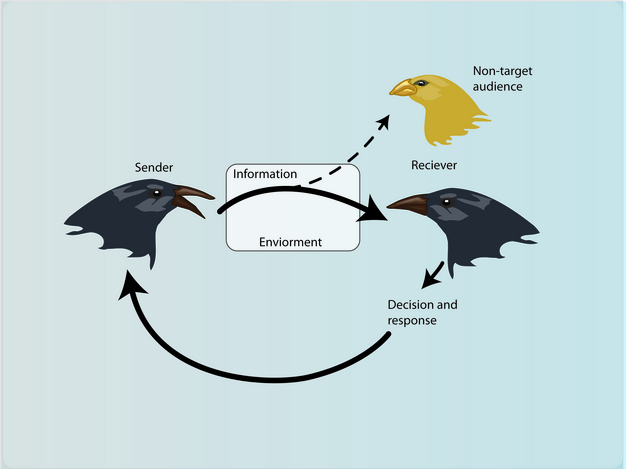
Figure 1
A model of animal communication.

Figure 2:
Photinus fireflies.
Courtesy of Tom Eisner.
Signal Modalities
Animals use a variety of sensory channels, or signal modalities, for communication. Visual signals are very effective for animals that are active during the day. Some visual signals are permanent advertisements; for example, the bright red epaulets of male red-winged blackbirds, Agelaius phoeniceus, which are always displayed, are important for territory defense. When researchers experimentally blackened epaulets, males were subject to much higher rates of intrusion by other males (Smith 1972). Alternatively, some visual signals are actively produced by an individual only under appropriate conditions. Male green anoles, Anolis carolinensis, bob their head and extend a brightly colored throat fan (dewlap) when signaling territory ownership.
Acoustic communication is also exceedingly abundant in nature, likely because sound can be adapted to a wide variety of environmental conditions and behavioral situations. Sounds can vary substantially in amplitude, duration, and frequency structure, all of which impact how far the sound will travel in the environment and how easily the receiver can localize the position of the sender. For example, many passerine birds emit pure-tone alarm calls that make localization difficult, while the same species produce more complex, broadband mate attraction songs that allow conspecifics to easily find the sender (Marler 1955). A particularly specialized form of acoustic communication is seen in microchiropteran bats and cetaceans that use high-frequency sounds to detect and localize prey. After sound emission, the returning echo is detected and processed, ultimately allowing the animal to build a picture of their surrounding environment and make very accurate assessments of prey location.
Compared to visual and acoustic modalities, chemical signals travel much more slowly through the environment since they must diffuse from the point source of production. Yet, these signals can be transmitted over long distances and fade slowly once produced. In many moth species, females produce chemical cues and males follow the trail to the female’s location. Researchers attempted to tease apart the role of visual and chemical signaling in silkmoths, Bombyx mori, by giving males the choice between a female in a transparent airtight box and a piece of filter paper soaked in chemicals produced by a sexually receptive female. Invariably, males were drawn to the source of the chemical signal and did not respond to the sight of the isolated female (Schneider 1974; Figure 3). Chemical communication also plays a critical role in the lives of other animals, some of which have a specialized vomeronasal organ that is used exclusively to detect chemical cues. For example, male Asian elephants, Elaphus maximus, use the vomeronasal organ to process chemical cues in female’s urine and detect if she is sexually receptive (Rasmussen et al. 1982).
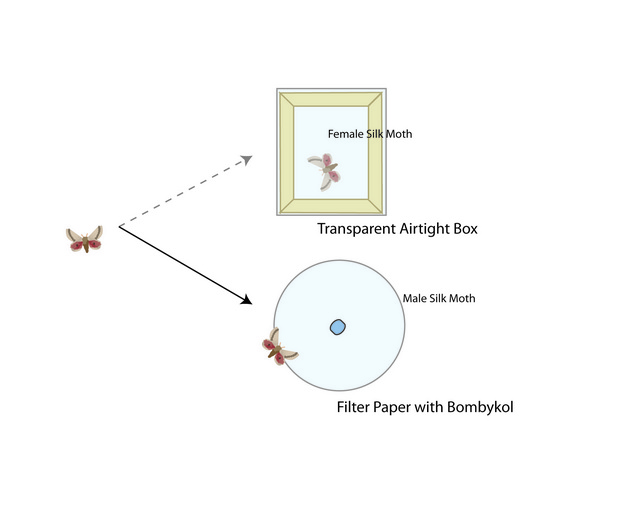
Figure 3
Male silkmoths are more strongly attracted to the pheromones produced by females (chemical signal) than the sight of a female in an airtight box (visual signal).
Tactile signals, in which physical contact occurs between the sender and the receiver, can only be transmitted over very short distances. Tactile communication is often very important in building and maintaining relationship among social animals. For example, chimpanzees that regularly groom other individuals are rewarded with greater levels of cooperation and food sharing (de Waal 1989).
For aquatic animals living in murky waters, electrical signaling is an ideal mode of communication. Several species of mormyrid fish produce species-specific electrical pulses, which are primarily used for locating prey via electrolocation, but also allow individuals searching for mates to distinguish conspecifics from heterospecifics. Foraging sharks have the ability to detect electrical signals using specialized electroreceptor cells in the head region, which are used for eavesdropping on the weak bioelectric fields of prey (von der Emde 1998).
Signal Functions
Some of the most extravagant communication signals play important roles in sexual advertisement and mate attraction. Successful reproduction requires identifying a mate of the appropriate species and sex, as well as assessing indicators of mate quality. Male satin bowerbirds, Ptilonorhynchus violaceus, use visual signals to attract females by building elaborate bowers decorated with brightly colored objects. When a female approaches the bower, the male produces an elaborate dance, which may or may not end with the female allowing the male to copulate with her (Borgia 1985). Males that do not produce such visual signals have little chance of securing a mate. While females are generally the choosy sex due to greater reproductive investment, there are species in which sexual roles are reversed and females produce signals to attract males. For example, in the deep-snouted pipefish, Syngnathus typhle, females that produce a temporary striped pattern during the mating period are more attractive to males than unornamented females (Berglund et al. 1997).
Communication signals also play an important role in conflict resolution, including territory defense. When males are competing for access to females, the costs of engaging in physical combat can be very high; hence natural selection has favored the evolution of communication systems that allow males to honestly assess the fighting ability of their opponents without engaging in combat. Red deer, Cervus elaphus, exhibit such a complex signaling system. During the mating season, males strongly defend a group of females, yet fighting among males is relatively uncommon. Instead, males exchange signals indicative of fighting ability, including roaring and parallel walks. An altercation between two males most often escalates to a physical fight when individuals are closely matched in size, and the exchange of visual and acoustic signals is insufficient for determining which animal is most likely to win a fight (Clutton-Brock et al. 1979).
Communication signals are often critical for allowing animals to relocate and accurately identify their own young. In species that produce altricial young, adults regularly leave their offspring at refugia, such as a nest, to forage and gather resources. Upon returning, adults must identify their own offspring, which can be especially difficult in highly colonial species. Brazilian free-tailed bats, Tadarida brasiliensis, form cave colonies containing millions of bats; when females leave the cave each night to forage, they place their pup in a crèche that contains thousands of other young. When females return to the roost, they face the challenge of locating their own pups among thousands of others. Researchers originally thought that such a discriminatory task was impossible, and that females simply fed any pups that approached them, yet further work revealed that females find and nurse their own pup 83% of the time (McCracken 1984, Balcombe 1990). Females are able to make such fantastic discriminations using a combination of spatial memory, acoustic signaling, and chemical signaling. Specifically, pups produce individually-distinct “isolation calls”, which the mother can recognize and detect from a moderate distance. Upon closer inspection of a pup, females use scent to further confirm the pup’s identity.
Many animals rely heavily on communication systems to convey information about the environment to conspecifics, especially close relatives. A fantastic illustration comes from vervet monkeys, Chlorocebus pygerythrus, in which adults give alarm calls to warn colony members about the presence of a specific type of predator. This is especially valuable as it conveys the information needed to take appropriate actions given the characteristics of the predator (Figure 4). For example, emitting a “cough” call indicates the presence of an aerial predator, such as an eagle; colony members respond by seeking cover amongst vegetation on the ground (Seyfarth & Cheney 1980). Such an evasive reaction would not be appropriate if a terrestrial predator, such as a leopard, were approaching.
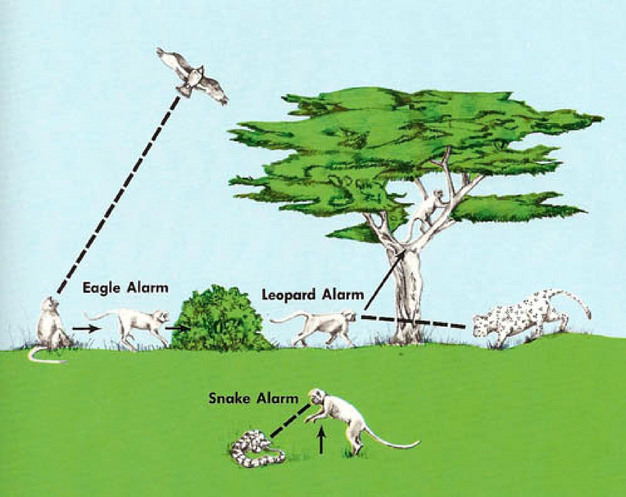
Figure 4
Vervet monkeys.
Many animals have sophisticated communication signals for facilitating integration of individuals into a group and maintaining group cohesion. In group-living species that form dominance hierarchies, communication is critical for maintaining ameliorative relationships between dominants and subordinates. In chimpanzees, lower-ranking individuals produce submissive displays toward higher-ranking individuals, such as crouching and emitting “pant-grunt” vocalizations. In turn, dominants produce reconciliatory signals that are indicative of low aggression. Communication systems also are important for coordinating group movements. Contact calls, which inform individuals about the location of groupmates that are not in visual range, are used by a wide variety of birds and mammals.
Overall, studying communication not only gives us insight into the inner worlds of animals, but also allows us to better answer important evolutionary questions. As an example, when two isolated populations exhibit divergence over time in the structure of signals use to attract mates, reproductive isolation can occur. This means that even if the populations converge again in the future, the distinct differences in critical communication signals may cause individuals to only select mates from their own population. For example, three species of lacewings that are closely related and look identical are actually reproductively isolated due to differences in the low-frequency songs produced by males; females respond much more readily to songs from their own species compared to songs from other species (Martinez, Wells & Henry 1992). A thorough understanding of animal communication systems can also be critical for making effective decisions about conservation of threatened and endangered species. As an example, recent research has focused on understanding how human-generated noise (from cars, trains, etc) can impact communication in a variety of animals (Rabin et al. 2003). As the field of animal communication continues to expand, we will learn more about information exchange in a wide variety of species and better understand the fantastic variety of signals we see animals produce in nature.
Glossary
Vomeronasal organ – auxiliary olfactory organ that detects chemosensory cues
Altricial – the state of being born in an immature state and relying exclusively on parental care for survival during early development
Refugia – areas that provide concealment from predators and/or protection from harsh environmental conditions
Conspecifics – organisms of the same species
References and Recommended Reading
Balcombe, J.P. Vocal recognition of pups by mother Mexican free-tailed bats, Tadarida brasiliensis mexicana. Animal Behaviour 39, 960-966 (1990).
Berglund, J., Rosenqvist G. and Bernet P. Ornamentation predicts reproductive success in female pipefish. Behavioral Ecology and Sociobiology 40, 145-150 (1997).
Clutton-Brock, T., Albon S., Gibson S. & Guinness F. The logical stag: Adaptive aspects of fighing in the red deer. Animal Behaviour 27, 211-225 (1979).
de Waal F.B.M. Food sharing and reciprocal obligations among chimpanzees. Journal of Human Evolution 18, 433–459 (1989).
Hauser, M. 1997. The Evolution of Communication. Cambridge, MA: MIT Press.
Lloyd, J.E. Aggressive mimicry in Photuris: signal repertoires by femmes fatales. Science 197, 452-453 (1975).
Marler, P. Characteristics of some animal calls. Nature 176, 6-8 (1955).
Martinez Well, M. & Henry C.S. The role of courtship songs in reproductive isolation among populations of green lacewings of the genus Chrysoperla. Evolution 46, 31-43 (1992).
McCracken, G.F. Communal nursing in Mexican free-tailed bat maternity colonies. Science 223, 1090-1091(1984).
Rabin, L.A., McCowan B., Hooper S.L & Owings D.H. Anthropogenic noise and its effect on animal communication: an interface between comparative psychology and conservation biology. International Journal of Comparative Psychology 16, 172-192 (2003).
Ryan M.J., Tuttle M.D., & Rand A.S. Sexual advertisement and bat predation in a neotropical frog. American Naturalist 119, 136–139 (1982).
Schneider, D. The sex attractant receptors of moths. Scientific American 231, 28-35 (1974).
Seyfarth, R.M., Cheney D.L. & Marler P. Monkey responses to three different alarm calls: Evidence for predator classification and semantic communication. Science 210, 801-803 (1980).
Smith, D. The role of the epaulets in the red-winged blackbird, (Agelaius phoeniceus) social system. Behaviour 41, 251-268 (1972).
Vehrencamp, S.L., Bradbury J.W., & Gibson R.M. The energetic cost of display in male sage grouse. Animal Behaviour 38, 885-896 (1989).
von der Emde, G. Electroreception. In D. H. Evans (ed.). The Physiology of Fishes, pp. 313-343. Boca Raton, FL: CRC Press (1998).
Wilson, E.O. Sociobiology: The New Synthesis. Cambridge, MA: Harvard University Press (1975).










