
Abstract
Epigenetic gene silencing induced by expanded repeats can cause diverse phenotypes ranging from severe growth defects in plants to genetic diseases such as Friedreich’s ataxia in humans. The molecular mechanisms underlying repeat expansion-induced epigenetic silencing remain largely unknown. Using a plant model with a temperature-sensitive phenotype, we have previously shown that expanded repeats can induce small RNAs, which in turn can lead to epigenetic silencing through the RNA-dependent DNA methylation pathway. Here, using a genetic suppressor screen and yeast two-hybrid assays, we identified novel components required for epigenetic silencing caused by expanded repeats. We show that FOURTH ULP GENE CLASS 1 (FUG1)—an uncharacterized SUMO protease with no known role in gene silencing—is required for epigenetic silencing caused by expanded repeats. In addition, we demonstrate that FUG1 physically interacts with ALFIN-LIKE 3 (AL3)—a histone reader that is known to bind to active histone mark H3K4me2/3. Loss of function of AL3 abolishes epigenetic silencing caused by expanded repeats. AL3 physically interacts with the chromodomain protein LIKE HETEROCHROMATIN 1 (LHP1)—known to be associated with the spread of the repressive histone mark H3K27me3 to cause repeat expansion-induced epigenetic silencing. Loss of any of these components suppresses repeat expansion-associated phenotypes coupled with an increase in IIL1 expression with the reversal of gene silencing and associated change in epigenetic marks. Our findings suggest that the FUG1–AL3–LHP1 module is essential to confer repeat expansion-associated epigenetic silencing and highlight the importance of post-translational modifiers and histone readers in epigenetic silencing.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


Data availability
All sequencing data are available at the National Center for Biotechnology Information short reads archive. The ChIP–seq reads are available in Bioproject ID PRJNA1080228, the small RNA sequence from fug1 is in Bioproject ID PRJNA1080246 and the RNA-seq data are in Bioproject ID PRJNA1080085. Source data are provided with this paper.
Code availability
The python script that we used to extract allele frequencies is available upon request to the corresponding authors.
References
Zhang, G., Wang, Y. & Andersen, E. C. Natural variation in C. elegans short tandem repeats. Genome Res. 32, 1852–1861 (2022).
Sawyer, L. A. et al. Natural variation in a Drosophila clock gene and temperature compensation. Science 278, 2117–2120 (1997).
Garg, P. et al. A phenome-wide association study identifies effects of copy-number variation of VNTRs and multicopy genes on multiple human traits. Am. J. Hum. Genet. 109, 1065–1076 (2022).
Barre, B. P. et al. Intragenic repeat expansion in the cell wall protein gene HPF1 controls yeast chronological aging. Genome Res. 30, 697–710 (2020).
Han, Z. et al. Natural variation underlies differences in ETHYLENE RESPONSE FACTOR17 activity in fruit peel degreening. Plant Physiol. 176, 2292–2304 (2018).
Press, M. O., McCoy, R. C., Hall, A. N., Akey, J. M. & Queitsch, C. Massive variation of short tandem repeats with functional consequences across strains of Arabidopsis thaliana. Genome Res. 28, 1169–1178 (2018).
Sureshkumar, S. et al. A genetic defect caused by a triplet repeat expansion in Arabidopsis thaliana. Science 323, 1060–1063 (2009).
Depienne, C. & Mandel, J. L. 30 years of repeat expansion disorders: what have we learned and what are the remaining challenges? Am. J. Hum. Genet. 108, 764–785 (2021).
Gall-Duncan, T., Sato, N., Yuen, R. K. C. & Pearson, C. E. Advancing genomic technologies and clinical awareness accelerates discovery of disease-associated tandem repeat sequences. Genome Res. 32, 1–27 (2022).
Rafehi, H. et al. An intronic GAA repeat expansion in FGF14 causes the autosomal-dominant adult-onset ataxia SCA50/ATX-FGF14. Am. J. Hum. Genet. 110, 105–119 (2023).
Pellerin, D. et al. Deep intronic FGF14 GAA repeat expansion in late-onset cerebellar ataxia. N. Engl. J. Med. 388, 128–141 (2023).
Erwin, G. S. et al. Recurrent repeat expansions in human cancer genomes. Nature https://doi.org/10.1038/s41586-022-05515-1 (2022).
Malik, I., Kelley, C. P., Wang, E. T. & Todd, P. K. Molecular mechanisms underlying nucleotide repeat expansion disorders. Nat. Rev. Mol. Cell Biol. 22, 589–607 (2021).
Finsterer, J. Phenotype and management of neurologic intronic repeat disorders (NIRDs). Rev. Neurol. https://doi.org/10.1016/j.neurol.2022.09.004 (2022).
Gottesfeld, J. M. Molecular mechanisms and therapeutics for the GAA.TTC expansion disease Friedreich ataxia. Neurotherapeutics 16, 1032–1049 (2019).
Eimer, H. et al. RNA-dependent epigenetic silencing directs transcriptional downregulation caused by intronic repeat expansions. Cell 174, 1095–1105 e1011 (2018).
Schneeberger, K. et al. SHOREmap: simultaneous mapping and mutation identification by deep sequencing. Nat. Methods 6, 550–551 (2009).
Castro, P. H. et al. Arabidopsis thaliana SPF1 and SPF2 are nuclear-located ULP2-like SUMO proteases that act downstream of SIZ1 in plant development. J. Exp. Bot. 69, 4633–4649 (2018).
Morrell, R. & Sadanandom, A. Dealing with stress: a review of plant SUMO proteases. Front. Plant Sci. 10, 1122 (2019).
Rosa, M. T. G. et al. Insights into the transcriptional and post-transcriptional regulation of the rice SUMOylation machinery and into the role of two rice SUMO proteases. BMC Plant Biol. 18, 349 (2018).
Wei, W. et al. The Alfin-like homeodomain finger protein AL5 suppresses multiple negative factors to confer abiotic stress tolerance in Arabidopsis. Plant J. 81, 871–883 (2015).
Lee, W. Y., Lee, D., Chung, W. I. & Kwon, C. S. Arabidopsis ING and Alfin1-like protein families localize to the nucleus and bind to H3K4me3/2 via plant homeodomain fingers. Plant J 58, 511–524 (2009).
Wysocka, J. et al. A PHD finger of NURF couples histone H3 lysine 4 trimethylation with chromatin remodelling. Nature 442, 86–90 (2006).
Miller, M. J., Barrett-Wilt, G. A., Hua, Z. & Vierstra, R. D. Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis. Proc. Natl Acad. Sci. USA 107, 16512–16517 (2010).
Augustine, R. C. & Vierstra, R. D. SUMOylation: re-wiring the plant nucleus during stress and development. Curr. Opin. Plant Biol. 45, 143–154 (2018).
Wen, D., Wu, J., Wang, L. & Fu, Z. SUMOylation promotes nuclear import and stabilization of polo-like kinase 1 to support its mitotic function. Cell Rep. 21, 2147–2159 (2017).
Park, I. S. et al. SUMOylation regulates nuclear localization and stability of TRAIP/RNF206. Biochem. Biophys. Res. Commun. 470, 881–887 (2016).
Zhao, Q. et al. GPS-SUMO: a tool for the prediction of sumoylation sites and SUMO-interaction motifs. Nucleic Acids Res. 42, W325–W330 (2014).
Beauclair, G., Bridier-Nahmias, A., Zagury, J. F., Saib, A. & Zamborlini, A. JASSA: a comprehensive tool for prediction of SUMOylation sites and SIMs. Bioinformatics 31, 3483–3491 (2015).
Zhao, S., Zhang, B., Yang, M., Zhu, J. & Li, H. Systematic profiling of histone readers in Arabidopsis thaliana. Cell Rep. 22, 1090–1102 (2018).
Molitor, A. M., Bu, Z., Yu, Y. & Shen, W. H. Arabidopsis AL PHD-PRC1 complexes promote seed germination through H3K4me3-to-H3K27me3 chromatin state switch in repression of seed developmental genes. PLoS Genet. 10, e1004091 (2014).
Romeo, K. et al. The SENP7 sumo-protease presents a module of two HP1 interaction motifs that locks HP1 protein at pericentric heterochromatin. Cell Rep. https://doi.org/10.1016/j.celrep.2015.01.004 (2015).
Maison, C. et al. The SUMO protease SENP7 is a critical component to ensure HP1 enrichment at pericentric heterochromatin. Nat. Struct. Mol. Biol. 19, 458–460 (2012).
Veluchamy, A. et al. LHP1 regulates H3K27me3 spreading and shapes the three-dimensional conformation of the Arabidopsis genome. PLoS ONE 11, e0158936 (2016).
Faivre, L. & Schubert, D. Facilitating transcriptional transitions: an overview of chromatin bivalency in plants. J. Exp. Bot. 74, 1770–1783 (2023).
Zhao, Z. et al. CRY2 interacts with CIS1 to regulate thermosensory flowering via FLM alternative splicing. Nat. Commun. 13, 7045 (2022).
Walter, M. et al. Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J. 40, 428–438 (2004).
Wu, F. H. et al. Tape–Arabidopsis sandwich—a simpler Arabidopsis protoplast isolation method. Plant Methods 5, 16 (2009).
Yoo, S. D., Cho, Y. H. & Sheen, J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat. Protoc. 2, 1565–1572 (2007).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Fletcher, S. J., Boden, M., Mitter, N. & Carroll, B. J. SCRAM: a pipeline for fast index-free small RNA read alignment and visualization. Bioinformatics https://doi.org/10.1093/bioinformatics/bty161 (2018).
Tasset, C. et al. POWERDRESS-mediated histone deacetylation is essential for thermomorphogenesis in Arabidopsis thaliana. PLoS Genet. 14, e1007280 (2018).
Zhang, Y. et al. Model-based analysis of ChIP-seq (MACS). Genome Biol. 9, R137, https://doi.org/10.1186/gb-2008-9-9-r137 (2008).
Liao, Y., Smyth, G. K. & Shi, W. The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 41, e108 (2013).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Conti, L., Kioumourtzoglou, D., O’Donnell, E., Dominy, P. & Sadanandom, A. OTS1 and OTS2 SUMO proteases link plant development and survival under salt stress. Plant Signal Behav. 4, 225–227 (2009).
Ewan, R. et al. Deubiquitinating enzymes AtUBP12 and AtUBP13 and their tobacco homologue NtUBP12 are negative regulators of plant immunity. New Phytol. 191, 92–106 (2011).
Bhagat, P. K., Roy, D. & Sadanandom, A. Expression, purification, and enzymatic analysis of plant SUMO proteases. Methods Mol. Biol. 2581, 109–119 (2023).
Weigel, D. & Glazebrook, J. Arabidopsis: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 2002).
Orosa, B. et al. SUMO conjugation to the pattern recognition receptor FLS2 triggers intracellular signalling in plant innate immunity. Nat. Commun. 9, 5185 (2018).
Schwab, R., Ossowski, S., Riester, M., Warthmann, N. & Weigel, D. Highly specific gene silencing by artificial microRNAs in Arabidopsis. Plant Cell 18, 1121–1133 (2006).
Clough, S. J. & Bent, A. F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743 (1998).
Pfaffl, M. W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29, e45 (2001).
Acknowledgements
We thank R. Clark (Utah, USA) for help with sequencing the fug1 mutant, A. Seleznev for help with bioinformatic analysis and H. Sako for initial characterization of suppressors. We thank J. Bowman, J. Coutts, Y. Guo, D. Smyth and L. Wong for discussions and comments on this manuscript. This work is supported by a Canadian Institute for Health Research grant PJT-178112 (E.R.), Department of Science and Technology – Science and Engineering Research Board Core grant (R.K.Y.), a National Health and Medical Research Council project grant APP1182090 (S.B. and S.S.), Australian Research Council Discovery Projects DP1095325 (S.B.) and DP190101818 (S.B.), and Australian Research Council Future Fellowships FT100100377 (S.B.), FT190100403 (S.S.), Australia–India Strategic Research Fund–Early and Mid-Career Fellowship (S.S.) and a Monash Larkins Fellowship (S.B.).
Author information
Authors and Affiliations
Contributions
Conceptualization: S.S. and S.B.; methodology: S.S., H.L., S.A., A.S., E.R. and S.B.; software: C.I.D., S.S. and S.B.; formal analysis: S.S., C.B., J.L., C.I.D., P.K.B., S.M., R.S., C.A., H.M.Y., N.S., G.F., P.T., A.S.Y., B.G.B., S.K., P.S., O.S.G. and S.B.; investigation: S.S., J.L., S.M., R.S., C.A., H.M.Y., N.S. and C.B.; writing—original draft: S.S.; writing—review and editing: S.S., A.S., S.A., H.L. and S.B.; visualization: S.S., S.A., H.M.Y. and S.B.; supervision: S.S., R.K.Y., S.A., A.S., H.L. and S.B.; project administration: S.S. and S.B.; funding acquisition: S.S. and S.B.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Plants thanks Jixian Zhai and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Genetic mapping of the suppressors of Bur-0 by SHOREMAP.
Allele frequency of EMS-type SNPs in the fraction of F2 pooled plants that display the suppression of the iil phenotype. a) 49-9 b) 61-7 and c) 57-3. High frequency alleles (>0.85) are coloured red and red crosses show the putative causal alleles.
Extended Data Fig. 2 fug1 mutant suppresses the temperature-dependent iil phenotype despite the presence of repeat expansion.
a) fug1 phenotype compared with Bur-0 at 23 °C and 27 °C. b) Suppression of the iil phenotype in fug1 is not due to the loss of a repeat expansion. Gels showing retention of the expanded repeat in fug1 plants compared with Bur-0 plants. The red boxes display the typical banding pattern of the repeat expansion. 1.5Kb band on the 1Kb plus (Invitrogen) is shown as the molecular marker. Each lane represents DNA analysed from individual plants with the Col-0 control in the first lane after the marker showing a non-expanded repeat. This is analysed multiple times with similar results.
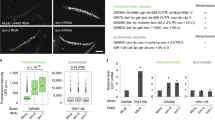
Extended Data Fig. 3 FUG1 is localised to the nucleus.
Representative maximum intensity projections (MIPs) of protoplasts of transgenic Bur-0 leaves harbouring either GFP (left) or GFP-FUG1 (right). Autofluorescence (purple) was separated using fluorescence life time. Scale bar=10 µM. The experiment is replicated more than twice with similar results.
Extended Data Fig. 4 Analysis of AL3 expression in transgenic lines.
a) Knocking down AL3 leads to increase in IIL1 expression. Independent plants representing multiple T2 lines (shown in X-axis) were analysed for IIL1 expression. Data were analysed through ANOVA and the statistical significance was tested with Tukey’s posthoc test. Different labels indicate significant differences (p < 0.05) in gene expression n = 5 in Bur-0, n = 4 in T2 Line 28 and n = 3 in all other T2 lines. b) Comparison of the expression levels of various fusion constructs of GFP-AL3. Primers against the GFP region allowed the measurement of transgenic AL3 levels in different backgrounds. Anlysis by ANOVA revealed that there were no significant differences in gene expression n = 3 in all samples except 35 S::GFP-AL3, where n = 6. Error bars represent standard errors of mean.
Extended Data Fig. 5 Perturbing the SUMOylation site of AL3 affects its sub-cellular localisation.
a) A schematic of the Fluorescence Lifetime Imaging (FLIM) assay using transformed protoplasts. Scale bar = 5 μm b) Representative maximum intensity projections showing the localisation of GFP or GFP-AL3 or AL3 harbouring K178R mutation (GFP-AL3-K178R) in Bur-0 and fug1 mutant backgrounds 2.5 μm. c) Box and whisker plots of nuclear:cytoplasm ratio of GFP (blue, n = 6 in both Bur-0 and fug1) or GFP-AL3 (cyan, n = 16 in Bur-0 and n = 24 in fug1) or GFP-AL3-K178R (orange, n = 8 in Bur-0 and n = 14 in fug1) in Bur-0 and fug1 mutant backgrounds. Box plots show the 50th percentile and the whiskers show the max and minimum with the mean shown by a line in the graph. Each dot represents the quantification from an individual protoplast expressing the corresponding transgene. Statistical comparisons were done with a two sided Student’s t-test. P-values: **<0.01, ***<0.001, ****<0.0001. The first lane of panel b and the left half (Bur) of panel c are also used in the Main Fig. 3(h & i). The protoplast experiments were carried out at least twice with similar results.
Extended Data Fig. 6 XVE::amiR-LHP1 lines show an increase in IIL1 levels upon estradiol induction.
a) Relative IIL1 expression levels. Comparison is made between Bur-0, empty vector controls as well as mock treated plants with that of estradiol treated samples in one-way ANOVA with the samples being separated as controls vs test. b) Comparison of IIL1 expression in plants that show the suppression of the iil phenotype and those that do not analysed together independent of the specific transgenic lines in one-way ANOVA c) Comparison of IIL1 expression in plants that show the suppression of the iil phenotype and those that do not from independent T2 lines. Expression analysis is from 2–3 independent plants as shown by individual data points on each of the bar graphs. The percentage of plants that show the suppression and the number of plants analysed are shown above each of the transgenic lines. The comparison of gene expression in this dataset is by two-sided Student’s T-test. Error bars represent standard errors of mean. p-values: *<0.05, **<0.01, ***<0.001, ****<0.0001.
Extended Data Fig. 7 Abundance of siRNAs that map to IIL1 locus in fug1 mutant.
Small RNAs that are typically found in Bur-0 at 27 °C (Fig. 2 of Eimer et al, Cell, 20184 were not observed in the fug 1 mutant. Small RNA profiles are generated as previously described4 using Small Complementary RnA Mapper (SCRAM)37. The genic region is shown in the bottom with the black boxes and lines representing exons and introns, respectively. Normalised coverage along with standard error shown as shadows is shown for different types of small RNAs as indicated by the colour code. Only small RNAs that mapped to the non-triplet repeat sequences of IIL1 are shown. Quantification of the sense and antisense small RNAs are shown in the positive and negative dimensions, respectively, along the y-axis. The green arrow indicates the position of the GAA/TTC repeat in the intron 3 of IIL1.
Extended Data Fig. 8 Effects of fug1 on gene silencing cannot be explained by changes in the RNA expression levels of the RdDM components.
A volcano plot depicting the changes in the RNA expression levels of genes in the RdDM pathway. Genes that show significant differential expression are marked in red. Differential expression was assessed by DESeq2 and only samples with an adjusted p-value after multiple correction (p-adj) are shown. An example for upregulated (AGO4) and downregulated (AGO9) are highlighted in the figure.
Supplementary information
Supplementary Information
Supplementary Tables 1–4.
Source data
Source Data Fig. 2
Unprocessed actin blot of the lower panel of Fig. 2c.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sureshkumar, S., Bandaranayake, C., Lv, J. et al. SUMO protease FUG1, histone reader AL3 and chromodomain protein LHP1 are integral to repeat expansion-induced gene silencing in Arabidopsis thaliana. Nat. Plants (2024). https://doi.org/10.1038/s41477-024-01672-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41477-024-01672-5
