
Abstract
In the US, high rates of preterm birth (PTB) and profound Black–White disparities in PTB have persisted for decades. This review focuses on the role of social determinants of health (SDH), with an emphasis on maternal stress, in PTB disparity and biological embedding. It covers: (1) PTB disparity in US Black women and possible contributors; (2) the role of SDH, highlighting maternal stress, in the persistent racial disparity of PTB; (3) epigenetics at the interface between genes and environment; (4) the role of the genome in modifying maternal stress–PTB associations; (5) recent advances in multi-omics studies of PTB; and (6) future perspectives on integrating multi-omics with SDH to elucidate the Black–White disparity in PTB. Available studies have indicated that neither environmental exposures nor genetics alone can adequately explain the Black–White PTB disparity. Preliminary yet promising findings of epigenetic and gene–environment interaction studies underscore the value of integrating SDH with multi-omics in prospective birth cohort studies, especially among high-risk Black women. In an era of rapid advancements in biomedical sciences and technologies and a growing number of prospective birth cohort studies, we have unprecedented opportunities to advance this field and finally address the long history of health disparities in PTB.
Impact
-
This review provides an overview of social determinants of health (SDH) with a focus on maternal stress and its role on Black–White disparity in preterm birth (PTB).
-
It summarizes the available literature on the interplay of maternal stress with key biological layers (e.g., individual genome and epigenome in response to environmental stressors) and significant knowledge gaps.
-
It offers perspectives that such knowledge may provide deeper insight into how SDH affects PTB and why some women are more vulnerable than others and underscores the critical need for integrating SDH with multi-omics in prospective birth cohort studies, especially among high-risk Black women.
Similar content being viewed by others


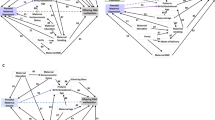
Overview: preterm birth (PTB) disparity in US Black women and possible contributors
Worldwide, about 11% of infants are born preterm (defined as birth before 37 weeks gestation) and 1 million die annually.1,2,3 In the US, there have been not only persistently high rates of PTB for decades but also a prolonged and profound racial disparity in PTB among Black women. In 2018, the rate of PTB among Black women (14%) was about 50% higher than the rate among White women (9%).3 Even more concerning is the finding of 40% excess risk for late PTB and 2.2 times higher risk for early PTB in Black women compared to White women.4 Early PTB is known to be associated with myriad perinatal and postnatal complications and sequelae.5,6,7,8 To date, the underlying mechanisms of PTB have remained elusive despite more than half a century of research. This has been partly explained as due to the fact that PTB is a complex trait determined by multiple environmental and genetic factors9 and because of the significant heterogeneity and complexity of biological pathways that could lead to PTB.10,11 This lack of progress after years of investigation underscores a critical need to go beyond traditional approaches to tackle PTB.
The earliest studies on PTB were primarily focused on uncovering socio-demographic, environmental, and clinical factors, and a number of them have been identified, including maternal race/ethnicity,12,13,14 age,15,16,17 education,18,19 income,20,21 place of birth,22 smoking,23,24,25 stress,26,27,28 (see section “Social determinants of health (SDH) and the role of maternal stress in the persistent Black–White disparity of PTB” below), social support,29,30 air pollution,31,32,33,34 and malnutrition.35,36 Two of the strongest risk factors appear to be a history of prior PTB37,38 and a woman’s own PTB outcome.39 Our early study showed that these two factors in combination greatly increased the risk of PTB or low birth weight.40 There are by now hundreds of studies, and reviews of those studies, pointing to the role of environmental and psychosocial exposures in PTB, though their findings and even their conclusions are still in some way limited by our incomplete understanding of the causes of PTB.
The field is confronted by persistent challenges. Foremost among these is that these identified factors still only explain a fraction of all PTB cases, and the identified associations vary by studies, populations, and individuals. At the same time, intervention trials focused on major risk factors aiming to reduce PTB have also yielded disappointing results. For most social-epidemiological studies, there has been an absence of biomarker data that would allow for further explorations of underlying biological mechanisms.
Over the past two decades, with rapid advancements in human genetics and high-throughput biotechnology, an increasing number of genome-wide association studies (GWAS), epigenome-wide association studies (EWAS), and other omics studies of PTB have been or are being conducted (see sections “Epigenetics: the interface of genes and environment in mediating the stress response,” “Role of the genome in modifying maternal stress–PTB associations,” and “Recent advances in multi-omics studies of PTB”).41,42,43,44,45,46,47,48,49,50,51,52 However, most GWAS have failed to yield promising results that can be replicated in other populations, including Black populations. The largest study to date identified some significant genes associated with PTB,43 but this study was limited to White populations. The role of genes in the continued disparities seen in Black populations still remains to be determined, though it has been suggested that genetic variation may not be the main cause of the Black–White disparity in PTB.53 The value of searching for gene-by-environment (G×E) interactions, which are typically overlooked in existing genetic studies, has been demonstrated by our studies.52,54,55,56 We have shown that maternal risk factors such as smoking,54 maternal pre-pregnancy obesity,55 and maternal perceived stress52 can interact with individual genetic variants to affect the risk of PTB. Such studies help to identify individuals at significantly higher risk of PTB in the presence of these risk factors. This is an important step forward for precision risk assessment and prevention as compared to a “one size fits all” approach.
In the following sections, we discuss the social determinants of PTB, with an emphasis on the role of maternal stress in PTB risk and the persistent Black–White disparity, as well as its biological embedding in the context of multi-omics. In this report, the terms African American and Black are used interchangeably, unless otherwise specified.
Social determinants of health (SDH) and the role of maternal stress in the persistent Black–White disparity of PTB
The ongoing search for modifiable factors that result in racial disparities in PTB has been broad in scope; this review will focus on SDH, as defined by the Centers for Disease Control.57 More specifically, SDH are “the economic and social conditions that influence individual and group differences in health status,”58 which influence an individual’s ability to engage in health promoting activities, and are themselves “shaped by the distribution of money, power, and resources at global, national, and local levels.”59,60,61 The growing list of SDH under study today is extensive and includes food security, housing stability, homelessness, violence exposure, structural racism, and immigration-related policies. While a comprehensive review of SDH and their roles in the Black–White disparity of PTB is beyond the scope and allowed space, here we primarily discuss one major component of SDH, maternal stress, and its relationships with PTB.
In general, stress, as defined by Cohen et al., is a process in which environmental demands exceed a person’s adaptive capacity to respond and results in changes that put a person at risk for disease.62 Key elements of stress include its timing, type, severity, and length of exposure, as well as the response to the stressor. Black populations have been shown to be disproportionally subject to psychosocial stressors and poor health outcomes, including PTB.63,64 One form of stress that consistently varies by race is the experience of discrimination and racism.65,66 Exposure to discrimination showed a twofold or higher risk for adverse birth outcomes related to residential segregation and neighborhood-level poverty.67 Since the release in 2004 of the report “Unequal treatment: confronting racial and ethnic disparities in health care” from the Institute of Medicine,68 an even greater focus on racism as a factor driving higher PTB rates in Black women has emerged. The report’s findings were clear—racism is one of the single most important factors in health disparities for African Americans as it relates to the provision of low-quality care and high burden of preventable causes of death including PTB. The relationship between racism and adverse birth outcomes may also be mediated by stress.69 These findings should not be surprising. However, many questions remain to be answered. Below we highlight a few.
There is a lack of consensus about the extent to which stress may account for the racial disparity in PTB.70 For example, a study by Grobman et al.69 demonstrated that Black women were more likely to have greater psychosocial burden across almost all domains compared to White women and that such racial/ethnic differences cannot be fully mitigated by income status. Such racial/ethnic disparity in psychosocial burden may mirror the disparity observed in birth outcomes, including PTB. In comparison, Almeida et al.,12 using data from the Pregnancy Risk Assessment Monitoring System, found that Black women had a higher risk of PTB relative to White women. While accounting for stress reduced the risk of PTB by 22%, this, however, did not fully explain the Black–White disparity in PTB. However, the study by Lu et al. did not support that maternal stress contributed significantly to racial disparities in PTB.71
Investigators working as far back as the 1940s have explored the association of maternal stress with birth outcomes.12,26,27,28,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92 However, after decades of studies such an association has not consistently been found. For example, some reported that mothers experiencing self-reported higher stress, stressful life events, and/or emotional disorders27,77,78,79,81,82,83,84,85,86,87,92,93,94 were at a higher risk of PTB, but others did not find such associations.71,80,88,90,91 It has been proposed that such inconsistency may be at least partly due to methodological variations in stress measurements across studies.28,80,87,93,94 Furthermore, a combination of different stressors, or stress plus other environmental exposures (i.e., smoking, air pollution, and heavy metal exposure),95 was found to have an even more profound impact on birth outcomes, including PTB.
Moreover, there is limited understanding of the biological embedding of SDH in PTB. Maternal stress during pregnancy can lead to a series of biochemical changes that may underlie the psychological and physiological consequences of maternal stress. The hypothalamic–pituitary–adrenal (HPA) axis is the principal endocrine system that is activated in response to stress. Specifically, maternal stress is thought to trigger norepinephrine and cortical release, activating placental corticotropin-releasing hormone gene expression, and leading to a cascade of events ending in PTB.96,97 Hyperactivity of the HPA axis in mothers and/or in the fetus was found to be associated with an altered risk of PTB,98,99 indicating a potential physiological link between stress exposure and risk of PTB. It is also likely that elevated maternal stress may contribute to PTB via inflammation/infection.100,101
Taken together, maternal stress is a socially based but biologically plausible risk factor for PTB. In the following sections, we summarize the current literature on the interplay of maternal stress with key biological layers (e.g., genome, epigenome, metabolome) in response to environmental stressors. Such knowledge may provide deeper insight into how maternal stress affects PTB and why some women are more vulnerable than others, to help move the field beyond studies focused on the identification of risk factors, and should also help to inform more targeted, precise, and effective interventions.
Epigenetics: the interface of genes and environment in mediating the stress response
Epigenetics—a mechanism for regulating gene expression without changes occurring in the DNA sequence—may represent a critical interface between individual genetic susceptibility and responses to environmental or psychosocial exposures.102,103 In contrast to the consistency of the genome, the epigenome is characterized as having dynamic and flexible changes in response to intra- and extra-cellular stimuli that can serve as modifiable biomarkers for environmental exposures.104 Epigenomic variations are largely established in utero, a period that is most sensitive to environmental perturbation and a critical time for the establishment of epigenetic variability.105,106,107,108 The major epigenetic mechanisms include DNA methylation (DNAm), histone modification, and non-coding RNAs (ncRNAs).109,110 DNAm, an addition of a methyl group to cytosines, occurs predominantly in cytosines located at 5’ of guanines (known as CpG dinucleotides). Although its function may vary, DNAm, when it occurs in promoter regions, generally is associated with gene silencing and repressing gene expression.111 Histone acetylation directly remodels chromatin rather than affecting messenger RNA (mRNA), which may affect nucleosome positioning, DNA wrapping, accessibility of chromatin to transcription factors, and regulate gene expression. ncRNAs could silence gene expression via RNA interference, which is commonly associated with posttranscriptional modification of mRNA. Among these mechanisms, DNAm has been studied most extensively in human studies because DNA is relatively stable compared to chromatin or RNA, and because recent technological advances make epigenome-wide DNAm profiling feasible in large cohorts.
As reviewed previously,112 a growing number of candidate-gene and epigenome-wide studies have showed that different forms and severity of maternal psychosocial stress have an influence on fetal DNAm (measured in cord blood or placental DNA),113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140 which may lead to an altered risk of PTB. Candidate-gene studies of this kind have mainly targeted specific genes involved in the human HPA axis.113,114,115,116,117,120,121,122,123,129,130,131,138 In particular, the NR3C1 gene, which encodes glucocorticoid receptors that mediate the stress response in humans, was found to have altered methylation levels at the promoter region in newborns whose mothers were exposed to stress and/or depression during pregnancy.113,114,115,116,121,131,138 Mulligan et al., in cord blood samples from 25 mother–newborn dyads, demonstrated a significant correlation between maternal stress, newborn methylation in the promoter region of the NR3C1 gene, and newborn birth weight, suggesting a potential role of NR3C1 DNAm in mediating the impact of prenatal stress exposure on birth outcomes.121 Maternal stress may also be associated with altered DNAm of other genes that are involved in the HPA axis, such as FKBP5,116,120,122,123 which encodes FK506-binding protein 51 that plays an important role in the negative feedback loop, OXTRs,139 which encode the receptors of oxytocin (OXT) that have stress-buffering effects, and HSD11B2 that encodes hydroxysteroid 11-beta dehydrogenase 2.123,130 It may also be associated with altered DNAm of other genes not in the HPA axis but that may be implicated in PTB such as IGF2132,133 and SLC6A4.119 Further studies to explore how these promising DNAm markers mediate the impact of maternal stress on the risk of PTB may contribute to our understanding of the biological mechanisms underlying PTB.
Several epigenome-wide associations with maternal stress have also been reported, although their findings await further validation and replication.124,125,126,127,128,135,136,137,140 Vangeel et al., by enrolling 22 versus 23 newborns who were exposed to the lowest or highest degree of maternal anxiety, respectively, identified and verified a differentiated methylated region (DMR) in the GABA-B receptor subunit 1 gene (GABBR1) in newborns that was associated with pregnancy anxiety. DNAm level of the GABBR1 gene was significantly associated with HPA axis response to a stressor.124 Burnst et al., in 207 subjects, investigated epigenome-wide placental DNAm in relation to maternal experiences of traumatic and non-traumatic stressors over her lifetime, which led to differential DNAm at 112 CpG sites. They also identified some significant pathways that play important roles in multiple physiological functions necessary for proper fetal development.125 Cardenas et al., in Project Viva, measured DNAm profiles in 479 infants at birth and found that newborns exposed to antidepressants in pregnancy had decreased DNAm levels in the gene body of ZNF575 (a gene involved in transcriptional regulation but with unknown specific functions), which was replicated in the Generation R study.126 Nemoda et al. performed genome-wide DNA methylation profiling in CD3+ T lymphocytes from 38 antepartum maternal and 44 neonatal cord blood samples via Illumina HumanMethylation 450K and reported that maternal depression was significantly associated with DNAm alteration at multiple CpG sites in newborns, most of which are involved in the immune system.127 Rijlaarsdam et al. conducted an epigenome-wide association meta-analysis of prenatal maternal stress, which, however, did not identify any Bonferroni-corrected DMRs associated with prenatal stress exposure, suggesting that there are no large effects of prenatal maternal stress exposure on neonatal DNA methylation.136
Maternal stress may also affect a mother’s own epigenetic profile, as supported by both animal models141,142 and human studies.120,143,144,145,146 Maternal FKBP5 methylation was inversely correlated with threat-based adverse childhood experiences and maternal posttraumatic stress disorder (PTSD) symptoms during pregnancy,120 which was independent of maternal FKBP5 rs136780 genotypes. Schechter et al. reported that maternal PTSD severity and parenting stress were negatively correlated with the mean percentage of methylation of the NR3C1 gene in mothers.147 A few epigenome-wide studies were also reported in mothers.127,148 Nemoda et al. identified no maternal CpG sites with altered DNAm levels in women exposed to depression.127 A recent study by Surkan et al. in the Boston Birth Cohort (BBC) showed that, although maternal perceived stress displayed no significant associations with maternal DNAm alterations, social support during pregnancy was significantly associated with maternal DNAm changes at multiple genes.148
Epigenetics is also posited as a potential mechanism driving racial disparities in PTB.70 First, the social construct of race and the propensity to use it as the means to determine how to treat others and develop policy makes it such that Blacks are more likely to experience stress and other adverse SDH than Whites,69 which could induce epigenetic changes (as described above) that may explain racial differences in PTB. Second, epigenetic levels at certain genes may vary across different ethnic populations, some of which may lead to differences in response to maternal stress and/or represent precursors for future disease risk. Findings from previous studies offer evidence for this. Salihu et al. studied umbilical cord blood DNAm of genes implicated in PTB from 22 Black neonates and 69 non-Black neonates and found differential DNAm in TNFAIP8 and PON1 genes among Black vs. non-Black infants.149 Soubry et al. reported a significant hypermethylation of the IGF2 H19 DMRs in newborns of Black mothers who reported use of anti-depressive drugs during pregnancy,150 while such associations were not observed in White mothers. Furthermore, differential risks for PTB were noted among recent African immigrants compared to U.S.-born Black women,22,151 which could not be explained by known risk factors.22 This finding may further indicate the role of acquired epigenetic inheritance in the underlying biology of prematurity, although further studies are needed.
As summarized above, although available research suggests that maternal stress can lead to epigenetic changes and that epigenetics may play a role in PTB etiology,152,153,154,155,156 these existing studies (especially EWAS) have had limited sample sizes and the data remain fragmented. Furthermore, the identification of stress-related DNAm signatures in mothers and newborns raise new questions about when and how these changes might occur and whether maternal stress affects fetal DNAm through changes in their own DNAm. There is also considerable interest in the possibility that environmental and psychosocial exposures result in epigenetic effects that can then be transmitted from one generation to the next, but so far there is no direct evidence for this in humans.157
Role of the genome in modifying maternal stress–PTB associations
Human development is known to be shaped by a complex interplay of the social environment with genetic potential, as are birth outcomes including PTB.158,159 It has been hypothesized that the inconsistent findings for the relationships between maternal stress exposure and birth outcomes, as reported previously,27,77,78,79,81,82,83,84,85,86,87,92,93,94 are at least partly due to differences in individual genetic susceptibility to stress. Boyce has proposed the “orchid vs. dandelion” theory,160 which suggests that certain genetic variants can increase a person’s susceptibility to stressors. This plausibility was further supported by previous studies that demonstrated the significant impact of the interaction between maternal genes and perceived stress on multiple child health outcomes, as discussed below.
Multiple genetic variants in the stress response pathways, including those in the HPA axis, may lead to individual differences in response to stress161,162 and then modify the relationships between stress exposure and different health outcomes, including birth outcomes. Increasing evidence has suggested the existence of gene × maternal stress interactions that may impact multiple child health issues, such as adolescent disruptive behavior,163 negative emotionality,164 child internalizing symptoms,165 childhood intelligence quotient,166 and bronchiolitis.167 A recent study showed that the HPA axis multi-locus genetic profile score, which reflects the additive risk of three candidate genes (CRHR1, FKBP5, and NR3C1) and maternal prenatal perceived stress, interacted to affect risk of adolescent depression.168
The existence of G×E interactions in PTB etiology has also been supported by studies from us and others, such as maternal obesity × gene interactions,55 maternal smoking × gene interactions,54,56,169 and bacterial vaginosis × gene interaction.170 However, a limited number of studies have been performed to identify the impact of gene × stress interactions on PTB or related birth outcomes. The study by Mparmpakas et al. suggested an interaction between maternal stress (or maternal negative attitude toward the pregnancy) and NR3C1 polymorphisms on fetal weight.171 It is believed that genome-wide analyses of gene × stress interactions may have the potential to identify novel pathways underlying the stress–PTB relationships. Our recent study in the BBC was the first to demonstrate a genome-wide significant PTPRD × stress interaction on the risk of spontaneous PTB in African Americans. In that study, Hong et al. performed genome-wide screening to identify the gene × stress interactions on the risk of spontaneous PTB (sPTB) in 1490 Black women.52 The authors reported that rs35331017, a T-allele insertion/deletion polymorphism in the PTPRD gene, was genome-wide significantly interacted with overall lifetime stress on the risk of sPTB (PG×E = 4.7 × 10−8): maternal lifetime stress was dose-responsively associated with an increased risk of sPTB in mothers carrying the II (insertion/insertion) genotype; but the opposite trend was observed in mothers carrying the heterozygous or DD (deletion/deletion) genotypes. This interaction was validated in both Black (PG×E = 0.088) and White mothers (PG×E = 0.023) from another independent cohort.52 These findings, if further confirmed, may provide new insight into individual susceptibility to stress-induced sPTB.
Recent advances in multi-omics studies of PTB
Besides genomics and epigenomics, an increasing number of other omics studies, including transcriptomics,172,173 microbiomics,174,175 metabolomics,176,177 and proteomics,178,179 have been performed in association with PTB. Overall, such single-omics studies, once again, could not fully capture the entire biological complexity of PTB. Recent significant technological advances and the rapidly decreasing costs of such high-throughput technologies have made it feasible to conduct multi-omics profiles in a single study cohort. An increasing number of analytical tools have also been developed to analyze and integrate multi-omics data. So far, a few reviews on multi-omics integration have been published to discuss its potential advantages over single-omics studies, analytical approaches and challenges, and its utility in clinical diagnosis and treatment.180,181,182
To our knowledge, only a very limited number of studies have performed multi-omics integration in PTB research, although with promising findings. Chien et al., in 10 full-term and 8 PTB infants, conducted integrative analyses of transcriptomics and proteomics. They found that 29 genes/proteins had consistently altered regulation in PTB. This study indicated that such dual-omics analyses can provide new insight into molecular mechanisms and identify candidate biomarkers associated with PTB.183 Chabrun et al., in 36 placental samples, performed combined analyses of methylomics and transcriptomics in association with intra-uterine growth restriction and related phenotypes, including PTB. They built machine learning models that had a high capacity for predicting PTB, with r2 of 0.83 between the predicted and the actual PTB score.184 Ghaemi et al., in 51 samples from 17 pregnant women who delivered at term, built multivariate predictive models for gestational age using the Elastic net algorithm to integrate the multi-omics datasets including transcriptomics, microbiomics, proteomics and metabolomics, which can significantly increase predictive power compared to models based on single-omics datasets.185 However, as with the other types of studies reviewed above, these currently available studies had limited samples sizes, and their findings call for replication in large populations.
Future perspective: integration of multi-omics with SDH to elucidate PTB disparity
Available studies to date have indicated that neither social–environmental risk factors nor genetics/epigenetics alone can adequately explain the persistent and striking Black–White PTB disparity. The preliminary yet promising findings of single-omics and multi-omics studies underscore the need to bring these pieces of puzzle together to gain a better and fuller understanding of the causes and underlying mechanisms driving PTB and racial disparities. The value of integrating SDH with multi-omics in prospective birth cohort studies lies in the following. From a scientific discovery and innovation perspective, this will ensure a strong foundation of basic science and methodology research in the field of SDH, including rigor and reproducibility. From a translational perspective, it will provide critically needed sensitive and objective evidence of SDH to inform public policy, social reform, health service organization and delivery, and clinical and public health programs.
As this review highlights, many gaps remain to achieve integration. First, there is no widely accepted methodology to measure SDH, including maternal stress. Research methodologies must account for the pervasive, chronic, and multidimensional experiences of interpersonal and structural racism throughout the life course. Second, the impact of SDH on PTB disparity is likely mediated by the joint effects of DNAm at multiple CpG sites and/or modified by numerous genetic factors, each of which only have a small effect size. The detection of such small effect markers in GWAS and EWAS requires very large sample sizes or innovative methodologies. Third, the potential involvement of both maternal and fetal genomes, epigenomes, and other omics (such as transcriptomics, proteomics, metabolomics, and microbiomics) underlying the pathogenesis of PTB requires future studies to include both maternal and fetal biospecimens for analyses. Fourth, although the importance of SDH and individual genetics/epigenetics in health and disease are well recognized, few have considered all these factors as well as other omics in the same study. A successful systems biology approach requires that multi-omics data be generated from the same set of samples to allow for data integration. Furthermore, few studies conducted to date have been longitudinal by design, which make them less likely to clarify temporal and causal relationships. Of greatest concern is that there has been a lack of multi-omics studies conducted among disadvantaged US minority populations including Black mothers and children who bear a disproportionally high burden of social adversities and disparate health outcomes.
We end this review by highlighting our aspiration to connect multi-level, multi-dimensional SDH data with multi-omics to better understand PTB and address health disparities in PTB. Given the growing recognition of the importance of SDH, the rapid advancement of biomedical sciences and technologies, and growing number of prospective birth cohort studies, we have unprecedented opportunities to advance this field both in terms of scientific discoveries and clinical and public health translation. Ultimately, these efforts may allow us to move beyond risk factor analysis to a deeper understanding of the underlying causes of the persistent disparity in PTB, finally leading to improved individual and population health for Black women and children and for all people.
References
Chawanpaiboon, S. et al. Global, regional, and national estimates of levels of preterm birth in 2014: a systematic review and modelling analysis. Lancet Glob. Health 7, e37–e46 (2019).
Blencowe, H. et al. National, regional, and worldwide estimates of preterm birth rates in the year 2010 with time trends since 1990 for selected countries: a systematic analysis and implications. Lancet 379, 2162–2172 (2012).
Centers for Disease Control and Prevetion. Preterm birth. https://www.cdc.gov/reproductivehealth/maternalinfanthealth/pretermbirth.htm (2019).
Martin, J. A., Hamilton, B. E., Osterman, M. J. K. & Driscoll, A. K. Births: final data for 2018. Natl Vital Stat. Rep. 68, 1–47 (2019).
Raghavan, R. et al. Preterm birth subtypes, placental pathology findings, and risk of neurodevelopmental disabilities during childhood. Placenta 83, 17–25 (2019).
He, H. et al. Preterm birth with childhood asthma: the role of degree of prematurity and asthma definitions. Am. J. Respir. Crit. Care Med. 192, 520–523 (2015).
Carr, H., Cnattingius, S., Granath, F., Ludvigsson, J. F. & Edstedt Bonamy, A. K. Preterm birth and risk of heart failure up to early adulthood. J. Am. Coll. Cardiol. 69, 2634–2642 (2017).
Barfield, W. D. Public health implications of very preterm birth. Clin. Perinatol. 45, 565–577 (2018).
Hodgson, E. J. & Lockwood, C. J. Preterm birth: a complex disease. Preterm Birth Prev. Manag. 1, 8–16 (2010).
Goldenberg, R. L., Culhane, J. F., Iams, J. D. & Romero, R. Epidemiology and causes of preterm birth. Lancet 371, 75–84 (2008).
Brou, L. et al. Dysregulated biomarkers induce distinct pathways in preterm birth. BJOG 119, 458–473 (2012).
Almeida, J., Becares, L., Erbetta, K., Bettegowda, V. R. & Ahluwalia, I. B. Racial/ethnic inequities in low birth weight and preterm birth: the role of multiple forms of stress. Matern. Child Health J. 22, 1154–1163 (2018).
Culhane, J. F. & Goldenberg, R. L. Racial disparities in preterm birth. Semin. Perinatol. 35, 234–239 (2011).
Tsai, H. J. et al. Association of genetic ancestry with preterm delivery and related traits among African American mothers. Am. J. Obstet. Gynecol. 201, 94.e1–94.e10 (2009).
Schempf, A. H., Branum, A. M., Lukacs, S. L. & Schoendorf, K. C. Maternal age and parity-associated risks of preterm birth: differences by race/ethnicity. Paediatr. Perinat. Epidemiol. 21, 34–43 (2007).
Keiser, A. M. et al. Risks of preterm birth among non-Hispanic black and non-Hispanic white women: effect modification by maternal age. Paediatr. Perinat. Epidemiol. 33, 346–356 (2019).
Aldous, M. B. & Edmonson, M. B. Maternal age at first childbirth and risk of low birth weight and preterm delivery in Washington State. JAMA 270, 2574–2577 (1993).
El-Sayed, A. M. & Galea, S. Temporal changes in socioeconomic influences on health: maternal education and preterm birth. Am. J. Public Health 102, 1715–1721 (2012).
Jansen, P. W. et al. Explaining educational inequalities in preterm birth: the generation R study. Arch. Dis. Child. Fetal Neonatal Ed. 94, F28–F34 (2009).
Olson, M. E., Diekema, D., Elliott, B. A. & Renier, C. M. Impact of income and income inequality on infant health outcomes in the United States. Pediatrics 126, 1165–1173 (2010).
DeFranco, E. A., Lian, M., Muglia, L. A. & Schootman, M. Area-level poverty and preterm birth risk: a population-based multilevel analysis. BMC Public Health 8, 316 (2008).
Elo, I. T., Vang, Z. & Culhane, J. F. Variation in birth outcomes by mother’s country of birth among non-Hispanic black women in the United States. Matern. Child Health J. 18, 2371–2381 (2014).
van den Berg, G., van Eijsden, M., Vrijkotte, T. G. & Gemke, R. J. Educational inequalities in perinatal outcomes: the mediating effect of smoking and environmental tobacco exposure. PLoS ONE 7, e37002 (2012).
Kyrklund-Blomberg, N. B., Granath, F. & Cnattingius, S. Maternal smoking and causes of very preterm birth. Acta Obstet. Gynecol. Scand. 84, 572–577 (2005).
Soneji, S. & Beltran-Sanchez, H. Association of maternal cigarette smoking and smoking cessation with preterm birth. JAMA Netw. Open 2, e192514 (2019).
Wadhwa, P. D., Entringer, S., Buss, C. & Lu, M. C. The contribution of maternal stress to preterm birth: issues and considerations. Clin. Perinatol. 38, 351–384 (2011).
Gilles, M. et al. Maternal hypothalamus-pituitary-adrenal (HPA) system activity and stress during pregnancy: effects on gestational age and infant’s anthropometric measures at birth. Psychoneuroendocrinology 94, 152–161 (2018).
Hoffman, M. C., Mazzoni, S. E., Wagner, B. D., Laudenslager, M. L. & Ross, R. G. Measures of maternal stress and mood in relation to preterm birth. Obstet. Gynecol. 127, 545–552 (2016).
Hetherington, E. et al. Preterm birth and social support during pregnancy: a systematic review and meta-analysis. Paediatr. Perinat. Epidemiol. 29, 523–535 (2015).
Surkan, P. J. et al. Paternal involvement and support and risk of preterm birth: findings from the Boston birth cohort. J. Psychosom. Obstet. Gynaecol. 40, 48–56 (2019).
Li, Q. et al. Effect of airborne particulate matter of 2.5 μm or less on preterm birth: a national birth cohort study in China. Environ. Int. 121, 1128–1136 (2018).
Wang, Y. Y. et al. Association of long-term exposure to airborne particulate matter of 1 μm or less with preterm birth in China. JAMA Pediatr. 172, e174872 (2018).
Guo, L. Q. et al. Ambient air pollution and adverse birth outcomes: a systematic review and meta-analysis. J. Zhejiang Univ. Sci. B 20, 238–252 (2019).
Bekkar, B., Pacheco, S., Basu, R. & DeNicola, N. Association of air pollution and heat exposure with preterm birth, low birth weight, and stillbirth in the US: a systematic review. JAMA Netw. Open 3, e208243 (2020).
Mason, J. B. et al. Opportunities for improving maternal nutrition and birth outcomes: synthesis of country experiences. Food Nutr. Bull. 33, S104–S137 (2012).
Olapeju, B. et al. Maternal postpartum plasma folate status and preterm birth in a high-risk US population. Public Health Nutr. 22, 1281–1291 (2019).
Goldenberg, R. L. & Culhane, J. F. Prepregnancy health status and the risk of preterm delivery. Arch. Pediatr. Adolesc. Med. 159, 89–90 (2005).
Kistka, Z. A. et al. Racial disparity in the frequency of recurrence of preterm birth. Am. J. Obstet. Gynecol. 196, 131.e1–6 (2007).
Wilcox, A. J., Skjaerven, R. & Lie, R. T. Familial patterns of preterm delivery: maternal and fetal contributions. Am. J. Epidemiol. 167, 474–479 (2008).
Wang, X., Zuckerman, B., Coffman, G. A. & Corwin, M. J. Familial aggregation of low birth weight among whites and blacks in the United States. N. Engl. J. Med. 333, 1744–1749 (1995).
Uzun, A., Dewan, A. T., Istrail, S. & Padbury, J. F. Pathway-based genetic analysis of preterm birth. Genomics 101, 163–170 (2013).
Burris, H. H., Baccarelli, A. A., Wright, R. O. & Wright, R. J. Epigenetics: linking social and environmental exposures to preterm birth. Pediatr. Res. 79, 136–140 (2016).
Zhang, G. et al. Genetic associations with gestational duration and spontaneous preterm birth. N. Engl. J. Med. 377, 1156–1167 (2017).
Parets, S. E., Conneely, K. N., Kilaru, V., Menon, R. & Smith, A. K. DNA methylation provides insight into intergenerational risk for preterm birth in African Americans. Epigenetics 10, 784–792 (2015).
Knight, A. K. & Smith, A. K. Epigenetic biomarkers of preterm birth and its risk factors. Genes 7, 15 (2016).
Beaumont, R. N. et al. Genome-wide association study of offspring birth weight in 86 577 women identifies five novel loci and highlights maternal genetic effects that are independent of fetal genetics. Hum. Mol. Genet. 27, 742–756 (2018).
Liu, X. et al. Variants in the fetal genome near pro-inflammatory cytokine genes on 2q13 associate with gestational duration. Nat. Commun. 10, 3927 (2019).
Modi, B. P. et al. Discovery of rare ancestry-specific variants in the fetal genome that confer risk of preterm premature rupture of membranes (PPROM) and preterm birth. BMC Med. Genet. 19, 181 (2018).
York, T. P. et al. Replicated umbilical cord blood DNA methylation loci associated with gestational age at birth. Epigenetics 15, 1243–1258 (2020).
Rappoport, N. et al. A genome-wide association study identifies only two ancestry specific variants associated with spontaneous preterm birth. Sci. Rep. 8, 226 (2018).
Stevenson, D. K. et al. The contributions of genetics to premature birth. Pediatr. Res. 85, 416–417 (2019).
Hong, X. et al. Genome-wide association study identifies a novel maternal gene x stress interaction associated with spontaneous preterm birth. Pediatr. Res. https://doi.org/10.1038/s41390-020-1093-1 (2020).
Burris, H. H. et al. Racial disparities in preterm birth in USA: a biosensor of physical and social environmental exposures. Arch. Dis. Child. 104, 931–935 (2019).
Wang, X. et al. Maternal cigarette smoking, metabolic gene polymorphism, and infant birth weight. JAMA 287, 195–202 (2002).
Hong, X. et al. Genome-wide approach identifies a novel gene-maternal pre-pregnancy BMI interaction on preterm birth. Nat. Commun. 8, 15608 (2017).
Tsai, H. J. et al. Role of African ancestry and gene-environment interactions in predicting preterm birth. Obstet. Gynecol. 118, 1081–1089 (2011).
Centers for Disease Control and Prevention. Social determinants of health: know what affects health. https://www.cdc.gov/socialdeterminants/index.htm (2020).
Braveman, P. & Gottlieb, L. The social determinants of health: it’s time to consider the causes of the causes. Public Health Rep. 129(Suppl 2), 19–31 (2014).
Preda, A. & Voigt, K. The social determinants of health: why should we care? Am. J. Bioeth. 15, 25–36 (2015).
Braveman, P. & Gruskin, S. Defining equity in health. J. Epidemiol. Community Health 57, 254–258 (2003).
Brennan Ramirez, L. K., Baker, E. A., & Metzler, M. Promoting Health Equity: A Resource to Help Communities Address Social Determinants of Health (U.S. Department of Health and Human Services, Centers for Disease Control and Prevention, Atlanta, 2008).
Cohen, S., Kessler, R. C. & Underwood, L. G. Measuring Stress: A Guide for Health and Social Scientists (Oxford University Press, New York, 1995).
Rosenberg, L., Palmer, J. R., Wise, L. A., Horton, N. J. & Corwin, M. J. Perceptions of racial discrimination and the risk of preterm birth. Epidemiology 13, 646–652 (2002).
Mustillo, S. et al. Self-reported experiences of racial discrimination and Black-White differences in preterm and low-birthweight deliveries: the CARDIA study. Am. J. Public Health 94, 2125–2131 (2004).
Mendez, D. D., Hogan, V. K. & Culhane, J. F. Institutional racism, neighborhood factors, stress, and preterm birth. Ethn. Health 19, 479–499 (2014).
Rankin, K. M., David, R. J. & Collins, J. W. Jr. African American women’s exposure to interpersonal racial discrimination in public settings and preterm birth: the effect of coping behaviors. Ethn. Dis. 21, 370–376 (2011).
Mehra, R., Boyd, L. M. & Ickovics, J. R. Racial residential segregation and adverse birth outcomes: a systematic review and meta-analysis. Soc. Sci. Med. 191, 237–250 (2017).
Nelson, A. Unequal treatment: confronting racial and ethnic disparities in health care. J. Natl Med. Assoc. 94, 666–668 (2002).
Grobman, W. A. et al. Racial/ethnic disparities in measures of self-reported psychosocial states and traits during pregnancy. Am. J. Perinatol. 33, 1426–1432 (2016).
Kramer, M. R., Hogue, C. J., Dunlop, A. L. & Menon, R. Preconceptional stress and racial disparities in preterm birth: an overview. Acta Obstet. Gynecol. Scand. 90, 1307–1316 (2011).
Lu, M. C. & Chen, B. Racial and ethnic disparities in preterm birth: the role of stressful life events. Am. J. Obstet. Gynecol. 191, 691–699 (2004).
Istvan, J. Stress, anxiety, and birth outcomes: a critical review of the evidence. Psychol. Bull. 100, 331–348 (1986).
Hobel, C. J. Stress and preterm birth. Clin. Obstet. Gynecol. 47, 856–880 (2004).
Dunkel Schetter, C. Psychological science on pregnancy: stress processes, biopsychosocial models, and emerging research issues. Annu. Rev. Psychol. 62, 531–558 (2011).
Beydoun, H. & Saftlas, A. F. Physical and mental health outcomes of prenatal maternal stress in human and animal studies: a review of recent evidence. Paediatr. Perinat. Epidemiol. 22, 438–466 (2008).
Paarlberg, K. M., Vingerhoets, A. J., Passchier, J., Dekker, G. A. & Van Geijn, H. P. Psychosocial factors and pregnancy outcome: a review with emphasis on methodological issues. J. Psychosom. Res. 39, 563–595 (1995).
Alder, J., Fink, N., Bitzer, J., Hosli, I. & Holzgreve, W. Depression and anxiety during pregnancy: a risk factor for obstetric, fetal and neonatal outcome? A critical review of the literature. J. Matern. Fetal Neonatal Med. 20, 189–209 (2007).
Shah, P. S., Shah, J. & Knowledge Synthesis Group on Determinants of Preterm/LBW Births Maternal exposure to domestic violence and pregnancy and birth outcomes: a systematic review and meta-analyses. J. Womens Health 19, 2017–2031 (2010).
Straub, H., Adams, M., Kim, J. J. & Silver, R. K. Antenatal depressive symptoms increase the likelihood of preterm birth. Am. J. Obstet. Gynecol. 207, 329.e1–329.e4 (2012).
Abeysena, C. & Jayawardana, P. & Seneviratne, R. de A. Effect of psychosocial stress and physical activity on preterm birth: a cohort study. J. Obstet. Gynaecol. Res. 36, 260–267 (2010).
Kim, D. R. et al. Elevated risk of adverse obstetric outcomes in pregnant women with depression. Arch. Womens Ment. Health 16, 475–482 (2013).
Kramer, M. S. et al. Stress pathways to spontaneous preterm birth: the role of stressors, psychological distress, and stress hormones. Am. J. Epidemiol. 169, 1319–1326 (2009).
Wadhwa, P. D., Sandman, C. A., Porto, M., Dunkel-Schetter, C. & Garite, T. J. The association between prenatal stress and infant birth weight and gestational age at birth: a prospective investigation. Am. J. Obstet. Gynecol. 169, 858–865 (1993).
Lobel, M. et al. Pregnancy-specific stress, prenatal health behaviors, and birth outcomes. Health Psychol. 27, 604–615 (2008).
Dole, N. et al. Maternal stress and preterm birth. Am. J. Epidemiol. 157, 14–24 (2003).
Kitai, T. et al. A comparison of maternal and neonatal outcomes of pregnancy with mental disorders: results of an analysis using propensity score-based weighting. Arch. Gynecol. Obstet. 290, 883–889 (2014).
Hedegaard, M., Henriksen, T. B., Secher, N. J., Hatch, M. C. & Sabroe, S. Do stressful life events affect duration of gestation and risk of preterm delivery? Epidemiology 7, 339–345 (1996).
Nordeng, H. et al. Pregnancy outcome after exposure to antidepressants and the role of maternal depression: results from the Norwegian Mother and Child Cohort Study. J. Clin. Psychopharmacol. 32, 186–194 (2012).
Yonkers, K. A. et al. Depression and serotonin reuptake inhibitor treatment as risk factors for preterm birth. Epidemiology 23, 677–685 (2012).
Suri, R. et al. Effects of antenatal depression and antidepressant treatment on gestational age at birth and risk of preterm birth. Am. J. Psychiatry 164, 1206–1213 (2007).
Andersson, L., Sundstrom-Poromaa, I., Wulff, M., Astrom, M. & Bixo, M. Neonatal outcome following maternal antenatal depression and anxiety: a population-based study. Am. J. Epidemiol. 159, 872–881 (2004).
Weber, K. A. et al. Periconceptional stressors and social support and risk for adverse birth outcomes. BMC Pregnancy Childbirth 20, 487 (2020).
Rondo, P. H. et al. Maternal psychological stress and distress as predictors of low birth weight, prematurity and intrauterine growth retardation. Eur. J. Clin. Nutr. 57, 266–272 (2003).
Chen, M. J., Grobman, W. A., Gollan, J. K. & Borders, A. E. The use of psychosocial stress scales in preterm birth research. Am. J. Obstet. Gynecol. 205, 402–434 (2011).
Cory-Slechta, D. A., Virgolini, M. B., Thiruchelvam, M., Weston, D. D. & Bauter, M. R. Maternal stress modulates the effects of developmental lead exposure. Environ. Health Perspect. 112, 717–730 (2004).
Hobel, C. J., Dunkel-Schetter, C., Roesch, S. C., Castro, L. C. & Arora, C. P. Maternal plasma corticotropin-releasing hormone associated with stress at 20 weeks’ gestation in pregnancies ending in preterm delivery. Am. J. Obstet. Gynecol. 180, S257–S263 (1999).
Hobel, C. J., Dunkel-Schetter, C. & Roesch, S. Maternal stress as a signal to the fetus. Prenat. Neonat. Med. 3, 116–120 (1998).
Gelaye, B. et al. Chronic HPA activity in mothers with preterm delivery: a pilot nested case-control study. J. Neonatal Perinatal Med. 13, 313–321 (2020).
Stirrat, L. I. et al. Altered maternal hypothalamic-pituitary-adrenal axis activity in obese pregnancy is associated with macrosomia and prolonged pregnancy. Pregnancy Hypertens. 4, 238 (2014).
Coussons-Read, M. E., Okun, M. L. & Nettles, C. D. Psychosocial stress increases inflammatory markers and alters cytokine production across pregnancy. Brain Behav. Immun. 21, 343–350 (2007).
Christian, L. M., Franco, A., Glaser, R. & Iams, J. D. Depressive symptoms are associated with elevated serum proinflammatory cytokines among pregnant women. Brain Behav. Immun. 23, 750–754 (2009).
Holland, N. Future of environmental research in the age of epigenomics and exposomics. Rev. Environ. Health 32, 45–54 (2017).
Feinberg, A. P. Phenotypic plasticity and the epigenetics of human disease. Nature 447, 433–440 (2007).
Kanherkar, R. R., Bhatia-Dey, N. & Csoka, A. B. Epigenetics across the human lifespan. Front. Cell Dev. Biol. 2, 49 (2014).
Tang, W. Y. & Ho, S. M. Epigenetic reprogramming and imprinting in origins of disease. Rev. Endocr. Metab. Disord. 8, 173–182 (2007).
Radford, E. J. et al. In utero effects. In utero undernourishment perturbs the adult sperm methylome and intergenerational metabolism. Science 345, 1255903 (2014).
Cook, J. D., Davis, B. J., Goewey, J. A., Berry, T. D. & Walker, C. L. Identification of a sensitive period for developmental programming that increases risk for uterine leiomyoma in Eker rats. Reprod. Sci. 14, 121–136 (2007).
Heijmans, B. T. et al. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl Acad. Sci. USA 105, 17046–17049 (2008).
Egger, G., Liang, G., Aparicio, A. & Jones, P. A. Epigenetics in human disease and prospects for epigenetic therapy. Nature 429, 457–463 (2004).
Zhang, L., Lu, Q. & Chang, C. Epigenetics in health and disease. Adv. Exp. Med. Biol. 1253, 3–55 (2020).
Deaton, A. M. et al. Cell type-specific DNA methylation at intragenic CpG islands in the immune system. Genome Res. 21, 1074–1086 (2011).
Nowak, A. L., Anderson, C. M., Mackos, A. R., Neiman, E. & Gillespie, S. L. Stress during pregnancy and epigenetic modifications to offspring DNA: a systematic review of associations and implications for preterm birth. J. Perinat. Neonatal Nurs. 34, 134–145 (2020).
Oberlander, T. F. et al. Prenatal exposure to maternal depression, neonatal methylation of human glucocorticoid receptor gene (NR3C1) and infant cortisol stress responses. Epigenetics 3, 97–106 (2008).
Palma-Gudiel, H., Cordova-Palomera, A., Eixarch, E., Deuschle, M. & Fananas, L. Maternal psychosocial stress during pregnancy alters the epigenetic signature of the glucocorticoid receptor gene promoter in their offspring: a meta-analysis. Epigenetics 10, 893–902 (2015).
Conradt, E., Lester, B. M., Appleton, A. A., Armstrong, D. A. & Marsit, C. J. The roles of DNA methylation of NR3C1 and 11beta-HSD2 and exposure to maternal mood disorder in utero on newborn neurobehavior. Epigenetics 8, 1321–1329 (2013).
Kertes, D. A. et al. Prenatal maternal stress predicts methylation of genes regulating the hypothalamic-pituitary-adrenocortical system in mothers and newborns in the Democratic Republic of Congo. Child Dev. 87, 61–72 (2016).
Cecil, C. A. et al. Environmental risk, oxytocin receptor gene (OXTR) methylation and youth callous-unemotional traits: a 13-year longitudinal study. Mol. Psychiatry 19, 1071–1077 (2014).
Kertes, D. A. et al. BNDF methylation in mothers and newborns is associated with maternal exposure to war trauma. Clin. Epigenetics 9, 68 (2017).
Devlin, A. M., Brain, U., Austin, J. & Oberlander, T. F. Prenatal exposure to maternal depressed mood and the MTHFR C677T variant affect SLC6A4 methylation in infants at birth. PLoS ONE 5, e12201 (2010).
Grasso, D. J. et al. Adverse childhood experiences, posttraumatic stress, and FKBP5 methylation patterns in postpartum women and their newborn infants. Psychoneuroendocrinology 114, 104604 (2020).
Mulligan, C. J., D’Errico, N. C., Stees, J. & Hughes, D. A. Methylation changes at NR3C1 in newborns associate with maternal prenatal stress exposure and newborn birth weight. Epigenetics 7, 853–857 (2012).
Duis, J. et al. Effect of genotype and maternal affective disorder on intronic methylation of FK506 binding protein 5 in cord blood DNA. Front. Genet. 9, 648 (2018).
Monk, C. et al. Distress during pregnancy: epigenetic regulation of placenta glucocorticoid-related genes and fetal neurobehavior. Am. J. Psychiatry 173, 705–713 (2016).
Vangeel, E. B. et al. Newborn genome-wide DNA methylation in association with pregnancy anxiety reveals a potential role for GABBR1. Clin. Epigenetics 9, 107 (2017).
Brunst, K. J. et al. Cumulative lifetime maternal stress and epigenome-wide placental DNA methylation in the PRISM cohort. Epigenetics 13, 665–681 (2018).
Cardenas, A. et al. Prenatal maternal antidepressants, anxiety, and depression and offspring DNA methylation: epigenome-wide associations at birth and persistence into early childhood. Clin. Epigenetics 11, 56 (2019).
Nemoda, Z. et al. Maternal depression is associated with DNA methylation changes in cord blood T lymphocytes and adult hippocampi. Transl. Psychiatry 5, e545 (2015).
Santos, H. P. Jr et al. Epigenome-wide DNA methylation in placentas from preterm infants: association with maternal socioeconomic status. Epigenetics 14, 751–765 (2019).
Mansell, T. et al. Maternal mental well-being during pregnancy and glucocorticoid receptor gene promoter methylation in the neonate. Dev. Psychopathol. 28, 1421–1430 (2016).
Stroud, L. R. et al. Prenatal major depressive disorder, placenta glucocorticoid and serotonergic signaling, and infant cortisol response. Psychosom. Med. 78, 979–990 (2016).
Hompes, T. et al. Investigating the influence of maternal cortisol and emotional state during pregnancy on the DNA methylation status of the glucocorticoid receptor gene (NR3C1) promoter region in cord blood. J. Psychiatr. Res. 47, 880–891 (2013).
Mansell, T. et al. The effects of maternal anxiety during pregnancy on IGF2/H19 methylation in cord blood. Transl. Psychiatry 6, e765 (2016).
Liu, Y. et al. Depression in pregnancy, infant birth weight and DNA methylation of imprint regulatory elements. Epigenetics 7, 735–746 (2012).
Gurnot, C. et al. Prenatal antidepressant exposure associated with CYP2E1 DNA methylation change in neonates. Epigenetics 10, 361–372 (2015).
Nieratschker, V. et al. MORC1 exhibits cross-species differential methylation in association with early life stress as well as genome-wide association with MDD. Transl. Psychiatry 4, e429 (2014).
Rijlaarsdam, J. et al. An epigenome-wide association meta-analysis of prenatal maternal stress in neonates: a model approach for replication. Epigenetics 11, 140–149 (2016).
Schroeder, J. W. et al. DNA methylation in neonates born to women receiving psychiatric care. Epigenetics 7, 409–414 (2012).
Cao-Lei, L. et al. DNA methylation signatures triggered by prenatal maternal stress exposure to a natural disaster: Project Ice Storm. PLoS ONE 9, e107653 (2014).
Unternaehrer, E. et al. Maternal adversities during pregnancy and cord blood oxytocin receptor (OXTR) DNA methylation. Soc. Cogn. Affect. Neurosci. 11, 1460–1470 (2016).
Non, A. L., Binder, A. M., Kubzansky, L. D. & Michels, K. B. Genome-wide DNA methylation in neonates exposed to maternal depression, anxiety, or SSRI medication during pregnancy. Epigenetics 9, 964–972 (2014).
Chertkow-Deutsher, Y., Cohen, H., Klein, E. & Ben-Shachar, D. DNA methylation in vulnerability to post-traumatic stress in rats: evidence for the role of the post-synaptic density protein Dlgap2. Int. J. Neuropsychopharmacol. 13, 347–359 (2010).
Cohen, H. & Zohar, J. An animal model of posttraumatic stress disorder: the use of cut-off behavioral criteria. Ann. NY Acad. Sci. 1032, 167–178 (2004).
Uddin, M. et al. Epigenetic and immune function profiles associated with posttraumatic stress disorder. Proc. Natl Acad. Sci. USA 107, 9470–9475 (2010).
Santos, H. P. Jr. et al. Discrimination exposure and DNA methylation of stress-related genes in Latina mothers. Psychoneuroendocrinology 98, 131–138 (2018).
Wright, M. L. et al. Parenting stress and DNA methylation among African Americans in the InterGEN Study. J. Clin. Transl. Sci. 1, 328–333 (2017).
Rodney, N. C. & Mulligan, C. J. A biocultural study of the effects of maternal stress on mother and newborn health in the Democratic Republic of Congo. Am. J. Phys. Anthropol. 155, 200–209 (2014).
Schechter, D. S. et al. Methylation of NR3C1 is related to maternal PTSD, parenting stress and maternal medial prefrontal cortical activity in response to child separation among mothers with histories of violence exposure. Front. Psychol. 6, 690 (2015).
Surkan, P. J. et al. Can social support during pregnancy affect maternal DNA methylation? Findings from a cohort of African-Americans. Pediatr. Res. 88, 131–138 (2020).
Salihu, H. M. et al. Racial differences in DNA-methylation of CpG sites within preterm-promoting genes and gene variants. Matern Child Health J. 20, 1680–1687 (2016).
Soubry, A. et al. The effects of depression and use of antidepressive medicines during pregnancy on the methylation status of the IGF2 imprinted control regions in the offspring. Clin. Epigenetics 3, 2 (2011).
Elsayed, A., Amutah-Onukagha, N. N., Navin, L., Gittens-Williams, L. & Janevic, T. Impact of immigration and duration of residence in US on length of gestation among Black women in Newark, New Jersey. J. Immigr. Minor Health 21, 1095–1101 (2019).
Lee, H. et al. DNA methylation shows genome-wide association of NFIX, RAPGEF2 and MSRB3 with gestational age at birth. Int. J. Epidemiol. 41, 188–199 (2012).
Schroeder, J. W. et al. Neonatal DNA methylation patterns associate with gestational age. Epigenetics 6, 1498–1504 (2011).
Behnia, F. et al. Fetal DNA methylation of autism spectrum disorders candidate genes: association with spontaneous preterm birth. Am. J. Obstet. Gynecol. 212, 533.e1–533.e9 (2015).
Sparrow, S. et al. Epigenomic profiling of preterm infants reveals DNA methylation differences at sites associated with neural function. Transl. Psychiatry 6, e716 (2016).
Hong, X. et al. Genome-wide DNA methylation associations with spontaneous preterm birth in US Blacks: findings in maternal and cord blood samples. Epigenetics 13, 163–172 (2018).
Grossniklaus, U., Kelly, W. G., Ferguson-Smith, A. C., Pembrey, M. & Lindquist, S. Transgenerational epigenetic inheritance: how important is it? Nat. Rev. Genet. 14, 228–235 (2013).
Strauss, J. F. 3rd et al. Spontaneous preterm birth: advances toward the discovery of genetic predisposition. Am. J. Obstet. Gynecol. 218, 294.e2–314.e2 (2018).
Dolan, S. M. Genetic and environmental contributions to racial disparities in preterm birth. Mt. Sinai J. Med. 77, 160–165 (2010).
Boyce, T. W. The Orchid and the Dandelion: Why Some Children Struggle and How All Can Thrive (Knopf, New York, 2019).
Kumsta, R. et al. Sex specific associations between common glucocorticoid receptor gene variants and hypothalamus-pituitary-adrenal axis responses to psychosocial stress. Biol. Psychiatry 62, 863–869 (2007).
Gerritsen, L. et al. HPA axis genes, and their interaction with childhood maltreatment, are related to cortisol levels and stress-related phenotypes. Neuropsychopharmacology 42, 2446–2455 (2017).
Massey, S. H. et al. Does MAOA increase susceptibility to prenatal stress in young children? Neurotoxicol. Teratol. 61, 82–91 (2017).
Green, C. G. et al. Prenatal maternal depression and child serotonin transporter linked polymorphic region (5-HTTLPR) and dopamine receptor D4 (DRD4) genotype predict negative emotionality from 3 to 36 months. Dev. Psychopathol. 29, 901–917 (2017).
O’Donnell, K. J., Glover, V., Holbrook, J. D. & O’Connor, T. G. Maternal prenatal anxiety and child brain-derived neurotrophic factor (BDNF) genotype: effects on internalizing symptoms from 4 to 15 years of age. Dev. Psychopathol. 26, 1255–1266 (2014).
Lamb, Y. N. et al. Perceived stress during pregnancy and the catechol-O-methyltransferase (COMT) rs165599 polymorphism impacts on childhood IQ. Cognition 132, 461–470 (2014).
Lee, E. et al. The effect of perinatal anxiety on bronchiolitis is influenced by polymorphisms in ROS-related genes. BMC Pulm. Med. 14, 154 (2014).
McKenna, B. G., Hammen, C. & Brennan, P. A. HPA-axis multilocus genetic profile score moderates the association between maternal prenatal perceived stress and offspring depression in early adulthood. Dev. Psychopathol. https://doi.org/10.1017/S0954579419001639 (2020).
Zheng, X. et al. Association of maternal CNVs in GSTT1/GSTT2 with smoking, preterm delivery, and low birth weight. Front. Genet. 4, 196 (2013).
Gomez, L. M. et al. Evidence of a gene-environment interaction that predisposes to spontaneous preterm birth: a role for asymptomatic bacterial vaginosis and DNA variants in genes that control the inflammatory response. Am. J. Obstet. Gynecol. 202, 386.e1–386.e6 (2010).
Mparmpakas, D. et al. Differential expression of placental glucocorticoid receptors and growth arrest-specific transcript 5 in term and preterm pregnancies: evidence for involvement of maternal stress. Obstet. Gynecol. Int. 2014, 239278 (2014).
Yadama, A. P., Mirzakhani, H., McElrath, T. F., Litonjua, A. A. & Weiss, S. T. Transcriptome analysis of early pregnancy vitamin D status and spontaneous preterm birth. PLoS ONE 15, e0227193 (2020).
Pereyra, S., Sosa, C., Bertoni, B. & Sapiro, R. Transcriptomic analysis of fetal membranes reveals pathways involved in preterm birth. BMC Med. Genomics 12, 53 (2019).
Bayar, E., Bennett, P. R., Chan, D., Sykes, L. & MacIntyre, D. A. The pregnancy microbiome and preterm birth. Semin. Immunopathol. 42, 487–499 (2020).
Fettweis, J. M. et al. The vaginal microbiome and preterm birth. Nat. Med. 25, 1012–1021 (2019).
Ernst, M. et al. Gestational age-dependent development of the neonatal metabolome. Pediatr. Res. https://doi.org/10.1038/s41390-020-01149-z (2020).
Menon, R. et al. Amniotic fluid metabolomic analysis in spontaneous preterm birth. Reprod. Sci. 21, 791–803 (2014).
Pan, J., Tian, X., Huang, H. & Zhong, N. Proteomic study of fetal membrane: inflammation-triggered proteolysis of extracellular matrix may present a pathogenic pathway for spontaneous preterm birth. Front. Physiol. 11, 800 (2020).
Parry, S. et al. Cervicovaginal fluid proteomic analysis to identify potential biomarkers for preterm birth. Am. J. Obstet. Gynecol. 222, 493.e1–493.e13 (2020).
Karczewski, K. J. & Snyder, M. P. Integrative omics for health and disease. Nat. Rev. Genet. 19, 299–310 (2018).
Chu, S. H. et al. Integration of metabolomic and other omics data in population-based study designs: an epidemiological perspective. Metabolites 9, 117 (2019).
Hasin, Y., Seldin, M. & Lusis, A. Multi-omics approaches to disease. Genome Biol. 18, 83 (2017).
Chien, C. W. et al. Transcriptomic and proteomic profiling of human mesenchymal stem cell derived from umbilical cord in the study of preterm birth. Proteomics Clin. Appl. 14, e1900024 (2020).
Chabrun, F. et al. Data-mining approach on transcriptomics and methylomics placental analysis highlights genes in fetal growth restriction. Front. Genet. 10, 1292 (2019).
Ghaemi, M. S. et al. Multiomics modeling of the immunome, transcriptome, microbiome, proteome and metabolome adaptations during human pregnancy. Bioinformatics 35, 95–103 (2019).
Acknowledgements
This work is supported in part by the grants from the National Institute of Health (R03HD096136, R21HD085556, 2R01HD041702, R01HD086013, R01HD098232, R01ES031272, R21AI154233, and R01ES031521) and the Johns Hopkins Population Center (NICHD R24HD042854).
Author information
Authors and Affiliations
Contributions
All the authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; drafted the article or revised it critically for important intellectual content; and gave final approval of the version to be published.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Patient consent
Not applicable.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hong, X., Bartell, T.R. & Wang, X. Gaining a deeper understanding of social determinants of preterm birth by integrating multi-omics data. Pediatr Res 89, 336–343 (2021). https://doi.org/10.1038/s41390-020-01266-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41390-020-01266-9
This article is cited by
-
Data-driven insights can transform women’s reproductive health
npj Women's Health (2024)
-
Association between very advanced maternal age women with gestational diabetes mellitus and the risks of adverse infant outcomes: a cohort study from the NVSS 2014–2019
BMC Pregnancy and Childbirth (2023)
-
Pregnant in a Pandemic: Mental Wellbeing and Associated Healthy Behaviors Among Pregnant People in California During COVID-19
Maternal and Child Health Journal (2023)
