
Abstract
What changes in brain function cause the debilitating symptoms of depression? Can we use the answers to this question to invent more effective, faster acting antidepressant drug therapies? This review provides an overview and update of the converging human and preclinical evidence supporting the hypothesis that changes in the function of excitatory synapses impair the function of the circuits they are embedded in to give rise to the pathological changes in mood, hedonic state, and thought processes that characterize depression. The review also highlights complementary human and preclinical findings that classical and novel antidepressant drugs relieve the symptoms of depression by restoring the functions of these same synapses and circuits. These findings offer a useful path forward for designing better antidepressant compounds.
Similar content being viewed by others
Introduction
Depression is a leading cause of mortality and morbidity, affecting ~5% of the US population. Its many devastating symptoms, including persistent pathological deficits in mood, responses to rewarding stimuli (i.e., anhedonia), motivation, and cognition, as well as a greatly increased risk of suicide, make it essential to find effective treatments. Most widely used therapeutics, which potentiate monoaminergic neurotransmission, take 3–8 weeks to relieve symptoms, complicating treatment optimization and delaying relief, and are ineffective in one third of patients. Any individual patient may present with a range of possible symptoms, longitudinal time-course, and co-morbidities, as well responsiveness to antidepressants. It is hoped that a better understanding of the underlying neurobiological deficits will allow for a more effective personalized, targeted treatment plan [1, 2].
The etiology of depression involves both genetic and environmental factors [3], with the latter accounting for roughly 60% of risk [4]. Interestingly, pathway analyses from genome wide association studies have implicated many genes involved in synaptic function, axonal projections, and neurogenesis [5], although each contributes only a small amount to overall risk. A common environmental factor that increases the likelihood of depressive episodes is stress. Depressed patients report more stressful life events than non-depressed subjects, including physical illness, troubled family relationships, and financial difficulty [6]. Excessive rumination is common in depression and represents a uniquely human form of psychosocial stress. Early life adversity and traumatic childhood experiences are also a strong predictor of whom is at risk for depression, undoubtedly by triggering life-long lasting reprogramming events during particularly sensitive developmental windows [7], including in cortico-mesolimbic reward circuits [8]. Many of these long-lasting changes are likely to be mediated by epigenetic and transgenerational mechanisms that are beyond the scope of this review [9, 10]. Inflammation, often implicated in the genesis of depression, may also increase the susceptibility of synapses and neurons to the deleterious effects of stress [11].
Although acute stress is adaptive, persistently elevated levels of stress are maladaptive. In this review, I will summarize the abundant evidence that chronic stress damages brain function, to a significant extent, through its deleterious effects on excitatory synapses, thereby impairing the circuits underlying the brain’s responses to rewarding and aversive stimuli, cognitive demand, and mood. I will also discuss evidence that a critical mechanism underlying antidepressant efficacy is restoration of the function of stress-damaged synapses, particularly within these same reward circuits. This topic has been reviewed extensively [12,13,14,15,16]. My goal is to provide an up-to-date summary of the evidence of synaptic dysfunction in depression and to discuss the impact of this cellular pathology in the context of circuit-level and systems-level interactions that together regulate mood, hedonic state and other symptom domains characterizing depression.
Stress, symptoms of depression, and the circuitry of reward and emotion
The nature of the changes produced in the brain by chronic stress that promote depression in susceptible individuals remains unknown, but one of the most well-defined brain hallmarks of chronic hyperactivation of the HPA axis and chronic elevation of glucocorticoids is damage to excitatory synapses [16]. Glucocorticoids normally regulate neuronal survival, excitability, proliferation, metabolism, and memory but persistently high glucocorticoid levels may promote depression by impairing these processes [17].
The range of behaviors altered in depression suggests that multiple synaptic circuits are adversely affected, with no one synapse or circuit likely to be uniquely responsible for generating the entirety of the depressed phenotype. We can speculate that different phenotypic symptom clusters may reflect anatomical differences in the circuits compromised in these patient groups [18]. The nucleus accumbens (NAc) is a critical link in cortico-mesolimbic reward circuits, integrating information from the prefrontal cortices (PFC), amygdala, and hippocampus to control the release of dopamine from the ventral tegmental area (VTA) back to the telencephalon [19,20,21,22,23]. There are also circuits involving the lateral habenula that display a relatively selective processing of negative valence stimuli and feed forward to the VTA. There is a great deal of evidence of dysfunction in both cortico-mesolimbic and habenular circuits in preclinical stress-based models of anhedonic states, as will be reviewed below, with VTA activation and dopamine release subject to bidirectional regulation by both rewarding and aversive stimuli. Dysregulation of dopamine release in motivating responses to rewarding and aversive stimuli, as well as cognitive demand, is a plausible mechanistic cause that can explain a range of depression symptoms [24,25,26,27,28,29].
Virtually all preclinical studies of utility for understanding depression involve the production of a depression-like neurobehavioral state by exposure of laboratory animals to various forms of chronic stress. Given the links between human depression and stressful life events, it is generally agreed that such models have a high degree of etiological construct validity, and many researchers consider stress-based models to have the greatest utility for depression research. Chronic stress-based models in rodents come in several varieties, but the common factor is repeated (typically daily) exposure to psychological and physiological stress beyond a period lasting for several weeks [30,31,32]. Physical stressors include mild unpredictable stressors (a rotating sequence of cage tilting, wet bedding, forced swim, etc) or several hours of physical restraint. One potential challenge is overcoming the habituation to a single stressor after repeated exposure [33]. Psychosocial stressors include social defeat stress and social isolation. Early life stressors include fragmentated maternal care and social isolation [34]. Chronic administration of exogenous corticosteroids (CORT) also mimics many behavioral and physiological aspects of chronic stress paradigms [35, 36].
These chronic stress models induce depressive-like behavioral states, resembling the symptom domains of human depression, and alter the putative underlying brain circuits in parallel with functional imaging findings in depressed humans [18, 37,38,39]. Importantly, these preclinical changes are reversed by chronic, but not acute, administration of classical monoaminergic antidepressants, such as selective serotonin reuptake inhibitors (SSRIs) [36, 40,41,42,43,44], presumably analogous to the delay for the therapeutic response to SSRIs in humans. They also respond quickly to acute administration of fast-acting antidepressant compounds like ketamine [45]. These models thus demonstrate some degree of predictive validity. Limitations of stress-based models include the induction of several anxiety-related behaviors, perhaps mirroring the co-morbidity and overlap of anxiety disorders and depression and some of their respective symptoms.
All chronic stress protocols produce behavioral changes with face validity to the symptom domains of human depression. Diagnosis of major depressive disorder (MDD) in humans requires two cardinal symptoms, persistent depression of mood and persistent anhedonia, or loss of pleasure in all activities that should be pleasurable. In addition, one or more other symptoms may occur, including impaired cognition, concentration, and attention; dysregulation of sleep; changes in appetite; fatigue; feelings of worthlessness; and suicidal thinking. Many of these are impossible to address in laboratory animals, particularly since detection of most human symptoms is traditionally dependent upon self-reporting by the patient. Tests of hedonic state and cognition, however, are readily tested in rodents in a variety of reward related and cognitive tasks. Impairment in these tasks is consistently observed after chronic stress and is clearly maladaptive. Other tests, with less obvious correspondence with human depression symptoms include various measures of ‘behavioral despair,’ such as the forced swim test (FST) and the tail suspension test. Somewhat easier to interpret are the learned helplessness tasks, in which repeated delivery of an inescapable negative stimulus, such as a foot shock, changes animals’ behavior such that they will not escape even when given an opportunity. Although widely used and sensitive to known and novel antidepressants, their utility and interpretation is less clear [46] and their use is discouraged by the National Institute of Mental Health. Because they have been widely used, I will nevertheless discuss them in this review. It is to be hoped that better, more readily translatable, tests of reward behaviors will become more widely used in preclinical studies [47,48,49] in order to improve the selection of antidepressant compounds to take forward to clinical development.
An advantage of using tests of hedonic state and cognition is that the circuits underlying these behaviors are well known [50,51,52,53,54,55]. We can thus direct our preclinical search for depression relevant causative mechanisms and antidepressant relevant responses to brain regions in which cellular and molecular descriptions of pathology and treatment can be understood in terms of the behavioral changes they are likely to cause.
A grossly oversimplified overview of these circuits is that the prefrontal cortices and hippocampus are devoted to detecting and decoding specific features of stimuli and the environment, including the social environment, to guide decision-making and ensure adaptive, flexible, and predictive responses to rewarding stimuli, whereas the lateral hypothalamus-lateral habenula circuits predominantly encode features of stimuli with negative emotional valence. Both of these networks are then integrated in the NAc and VTA to regulate the release of dopamine. The feedback release of dopamine in these same regions than helps to reinforce and sculpt subsequent responses to rewarding and aversive stimuli.
The hippocampus
An impaired ability to think and concentrate, which encompasses deficits in memory, executive function, and attention, is one of the major symptom categories used to diagnose MDD and contributes significantly to the overall disability associated with depression [56, 57]. The hippocampus is well known to be essential in declarative and spatial memory formation from both animal studies and humans. It is also known to be an important regulator of the hypothalamic-pituitary axis, mediating negative feedback of stress hormone secretion.
The role of the hippocampus in reward related behaviors is becoming increasingly apparent [58]. Numerous changes in the electrophysiological responses of hippocampal neurons related to various aspects of reward have been described, including an increase in firing rates as animals approach a reward location [59, 60]. This is thought to result because the learning of reward locations results in the reorganization of place fields so that areas near those locations become overrepresented [61]. Hippocampal ensemble firing patterns also help animals correctly predict which direction they need to move in order to reach a chosen distant goal [62]. Importantly, receipt of a reward triggers an increase in the rate of spike-wave ripples [63], which are known to enhance place field stability, and may thereby promote learning of reward locations [64].
There are also strong, and psychologically impactful, interactions between memory and emotions [65]. Excessive rumination on memories of emotionally significant negative events from the past is positively correlated with incidence of depression and anxiety [66]. Dysfunction of the hippocampal–prefrontal circuits that underlie extinction of emotional memories is postulated to contribute to excessive negative rumination [67]. Proper ventral hippocampus-to-mPFC signaling is also necessary to recall social memories [68].
McEwen and colleagues first discovered that chronic stress and administration of exogenous corticosteroids lead to neuronal death in the hippocampus in rodents, as well as atrophy of the distal pyramidal cell dendrites and loss of dendritic spines [69,70,71,72,73,74]. These effects are consistent in all principal cell types and across multiple forms of physical and social stress [75,76,77].
Hippocampal atrophy is also one of the most consistent anatomical findings in imaging studies of human depression [78,79,80]. Hippocampal volume loss in depressed patients is correlated with memory dysfunction [81]. As in animal models, human hippocampal atrophy can result solely from elevated corticosteroid levels. Selective atrophy of the hippocampus is observed in patients with Cushing’s Syndrome and other diseases with chronically elevated cortisol levels, and these morphological changes are reversed by treatments that lower cortisol levels [82]. These data suggest that chronic stress, either from traumatic life events or psychosocial stressors, like excessive negative rumination, is the cause of the hippocampal atrophy in depression. There is also evidence that early life trauma can cause lifelong decreases in hippocampal volume [83], suggesting that preexisting hippocampal atrophy may predispose some people to the onset of depression later in life. Treatment with conventional antidepressants diminishes the atrophy in human depression [84], including recent longitudinal studies in previously drug naïve patients [85, 86], suggesting that the anatomical changes may underlie functional improvements.
How might stress and elevated corticosteroids produce this hippocampal dendritic atrophy and volume loss? Preclinical studies have shown that chronic stress and exogenous corticosteroid administration decrease the expression of several neurotrophic factors and their receptors, notably brain derived neurotrophic factor (BDNF) and its receptor tropomyosin receptor kinase B (trkB)(for review see: [87]. Furthermore, hippocampal and prefrontal cortical BDNF and trkB levels are decreased in postmortem tissue taken from suicide victims compared to tissue obtained from non-suicidal control brains [88]. The dentate gyrus is a unique source of ongoing neurogenesis in the adult brain, at least in nonhuman animals, and there is preclinical evidence that the birth and maturation of new granule cells may play a role in depression and antidepressant drug actions [89]. Preclinical studies have shown that the genesis of new neurons in the dentate gyrus from stem cells is impaired after chronic stress and exogenous corticosteroid administration [90,91,92].
The link between these phenomena and the genesis of depression was considerably strengthened with the finding that conventional antidepressants promote BDNF expression [87, 93] and neurogenesis [94,95,96]. A neurotrophic hypothesis of depression has been proposed, which posits that a decrease in neurotrophic factor signaling as the root of dysgenesis of dentate granule cells and atrophy of dendrites and synapses in depressed brain [87, 97, 98]. This hypothesis is complementary to the suggestion that stimulation of neurotrophic signaling and neurogenesis by antidepressants explains their therapeutic actions [99,100,101]. The explanatory power of neurogenesis in depression and antidepressant drug effects is complicated by the fact that neurogenesis is only observed in the dentate gyrus, putting a severe limit on the possible depression symptom domains it can impact, and by recent findings that neurogenesis may be very limited or absent in humans [102]. Nevertheless, it is particularly exciting that several slowly and rapidly acting antidepressant compounds, including fluoxetine and other classical antidepressants, and ketamine and its metabolites, have recently been shown to bind directly to the trkB receptor and thereby promote its activation by BDNF [103].
Beyond dendritic atrophy, there are dramatic alterations in synaptic structure and function in animal models based on chronic stress [104], including altered synaptic currents [105], deficits in synaptic plasticity in CA1 and DG [106], and impaired local circuit function [107]. In the hippocampus, several types of chronic stress cause a loss of distal apical dendritic branches in CA3 and CA1 pyramidal cells and a loss in the size and number of dendritic spines [71, 74, 108]. Nevertheless, the evidence of stress-induced changes in excitatory synaptic transmission in hippocampal pyramidal cells is somewhat mixed. In this regard, it is important to recognize that the effects of acute stress are generally adaptive and very different from the maladaptive consequences of chronic stress [104]. In area CA3, chronic stress enhances synaptic currents mediated by NMDARs but has no effect on AMPARs [109]. In contrast, chronic stress has no effect on basal synaptic strength at Schaffer collateral (SC) synapses in area CA1, although it does impair SC-CA1 long-term potentiation (LTP) [106, 110]. In contrast, at the temporoammonic (TA) synapses formed by entorhinal cortical afferents to the distal dendrites of CA1 pyramidal cells, where atrophy is most apparent, AMPAR-mediated basal synaptic transmission is impaired by chronic stress or exogenous corticosteroid administration, but LTP is unaffected [36, 110]. The functional consequence of stress-weakened TA-CA1 synaptic transmission is an impairment of spatial memory consolidation [36, 110]. Chronic social defeat stress also induces dendritic spine atrophy in CA1 pyramidal cells [111]. There may also be lasting, life-long consequences of early life chronic stress. Chronic adolescent stress in female rats caused a decrease in hippocampal SC-CA1 synaptic strength in adults, accompanied by impaired cognitive flexibility [112].
The mechanisms underlying the ability of chronic stress and exogenous corticosteroid administration to decrease excitatory synaptic function are presumably related to anatomical loss of dendritic spines and decreased expression of AMPARs. For example, GluA1 expression is decreased by 50% in the distal dendrites of area CA1 by chronic stress and glucocorticoid elevation [36, 110]. It is possible that these changes in glutamate expression and function, if occurring at multiple brain regions within reward circuits, are sufficient to cause depression-relevant behavioral phenotypes. Global deletion of the gene for the GluA1 subunit of the AMPAR, for example, results in a depression-like phenotype in the learned helplessness test [113]. Mice in which the GluA1 gene cannot be phosphorylated by calcium/calmodulin-dependent kinase (CaMK), thereby impairing their insertion into synapses, display altered behavior in the sucrose preference and novelty suppressed feeding tests, identical to those produced by chronic stress [44].
It has also been suggested that the ability of allostatic load to affect excitatory synaptic function may influence susceptibility to depression. In the chronic social defeat model, for example, stress susceptible mice had larger decreases in GluA1 expression in area CA1, compared to resilient mice [114]. The authors further examined genetic polymorphisms in genes encoding GluA1 subunits and found a single nucleotide polymorphism that correlated with vulnerability to stress. How this compares to human GluA1 gene SNPs was not described, but this evidence converges with human data implicating AMPARs [115]. Gene clusters associated with excitatory synaptic function are also commonly identified in genetic screens in suicide and depression [116,117,118] and in preclinical studies [119].
On a network level, chronic stress deceases the spatial tuning of hippocampal cell discharges, as well as the power of gamma oscillations in local field potentials [120]. The power of gamma oscillations contributes to working memory capacity, is reduced in patients with MDD, and is normalized with symptomatic remission [121]. GABAergic inhibition is also affected by chronic stress [122] and may contribute to the altered balance of excitation and inhibition as well as defects in ensemble activity. These results suggest that chronic stress-induced changes in synaptic function impair local network function and thereby impair the neural computations underlying cognitive processing. Together these data make a strong argument that disruption of hippocampal glutamatergic transmission, specifically AMPAR function, accompanies and may cause the depression-like behavioral state seen in preclinical models.
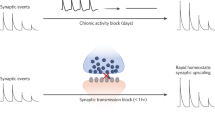
A common feature of all known antidepressants and many antidepressant treatments under development is their ability to reverse the effects of stress on synaptic structure and function, although they may produce this effect through various mechanisms. Serotonin increases phosphorylation of AMPA receptors and increases their delivery to the cell surface [123], an effect that is mimicked by the tricyclic antidepressant imipramine [124] and by chronic administration of the SSRI fluoxetine [125]. Chronic, but not acute, administration of conventional antidepressants also reverses stress-induced dendritic atrophy in rodents [126].
We have observed that chronic, but not acute, administration of fluoxetine restores the strength of stress-weakened TA-CA1 excitatory synapses, concomitant with the recovery of normal hedonic behavior [44, 120]. We observed that this restoration of synaptic strength and hedonic behavior was absent in mice with genetically modified GluA1 subunits that are incapable of being phosphorylated by calcium/calmodulin-dependent protein kinase (CaMK). We concluded that the antidepressant actions of SSRIs are mediated, in part, by a rapid insertion of AMPARs into synapses, due to 5HT1BR-dependent activation of CaMK and phosphorylation of GluA1 subunits at serine 831, a mechanism shared with canonical NMDAR-dependent long-term potentiation, as well as a slower restoration in GluA1 expression levels. Similarly, fluoxetine normalizes stress-induced changes in dendritic spine and glutamate receptor expression in the amygdala [127]. Fluoxetine also produces rapid disinhibition in hippocampal brain slices [128], which could also contribute to a net increase in excitation in vivo.
Restoration of stress-impaired excitatory synaptic function also appears to be a fundamental mechanism underlying the actions of known fast acting antidepressant drugs and potential therapeutic compounds. Extensive experience with human patients has established that ketamine exerts rapid, robust antidepressant, antianhedonic, and antisuicidal actions within two hours of administration that are sustained for at least seven days in patients with major depression following a single infusion or intranasal administration [129,130,131,132,133]. Ketamine’s antidepressant effects compared to placebo controls are further supported by several thorough meta-analyses [134,135,136]. Ketamine also exerts strong antidepressant-like actions in preclinical models, including restoration of reward behavior in chronically stressed animals, promotion of dendritic spine formation, restoration of synaptic strength, and promotion of expression of pro-synaptic genes [45]. Three classes of mechanism have been proposed to account for these therapeutic actions on the basis of preclinical work, all of which involve plasticity of excitatory synapses.
The first hypothesis might be called the ‘glutamate surge’ or ‘activity-dependent plasticity’ model. Based on early microdialysis measurements of increased extracellular glutamate release after a single administration of a therapeutic dose of ketamine [137], it is now known that ketamine produces increased synchronous neuronal activity in the prefrontal cortex and hippocampus, particularly in the gamma frequency band (30–80 Hz) in preclinical studies and in humans [138,139,140,141]. These oscillations are likely to result from disinhibition produced by a selective block of NMDARs on GABAergic interneurons [142, 143]. These high frequency oscillations lead to activation of endogenous activity-dependent synaptic strengthening mechanisms [144], including (paradoxically) NMDA-receptor dependent LTP, increases in dendritic spine number [145], and promotion of BDNF-trkB signaling [146], as we and others have speculated [13, 14, 147, 148].
Indeed, this potential mechanism may be shared by almost all other compounds exerting rapid antidepressant actions in clinical and/or pre-clinical studies, such as AZD6765 [149], mGluR2/3 antagonists [150], alpha5 subunit-selective GABAAR-NAMs [151, 152], and the ketamine metabolite (2 R,6 R)-HNK [153], all of which enhance EEG power in the gamma frequency band.
The second explanation for ketamine’s actions on excitatory synapses is the synaptic scaling hypothesis [154]. This hypothesis posits that ketamine restores stress-impaired excitatory synapses by blocking a subset of NMDARs that are preferentially activated by spontaneous glutamate release. When these receptors are inhibited, postsynaptic Ca2+ influx is also inhibited, decreasing the activity of the calcium/calmodulin-dependent protein kinase and eukaryotic elongation factor 2 kinase (eEF2K). Activated eEF2K phosphorylates and inhibits eEF2, an elongation factor that promotes global translation of mRNAs. Reduced eEF2K activity thus de-suppresses eEF2 and thereby promotes protein translation. Among the proteins whose synthesis is promoted are GluA1 receptors and BDNF [155,156,157,158]. As a result, synaptic AMPAR expression is increased [154, 159]. As predicted, acute inhibition of eEF2K increases mEPSC amplitude in cultured hippocampal cells and occludes the actions of ketamine on mEPSC amplitude [159]. This hypothesis cannot be readily applied to other fast acting antidepressant compounds that do not target NMDARs. It is also difficult to reconcile with findings that other NMDAR antagonists that do not appear to have antidepressant activity. Finally, it was observed that five days of treatment with the monoaminergic antidepressant citalopram failed to produce antidepressant behavioral effects in mice but did decrease phosphorylation of eEF2 levels [160]. It remains to be determined whether reduced eEF2 phosphorylation is necessary and sufficient for the actions of rapidly acting antidepressants other than ketamine.
Lastly, there is an exciting recent hypothesis that ketamine may exert its antidepressant actions not by blocking NMDARs, but rather by its neuroactive metabolites, particularly the hydroxynorketamines (HNKs). Zanos et al. [153] reported that the behavioral actions of ketamine in mice could be eliminated by deuterating the molecule to greatly reduce its rate of hydrolysis, suggesting that ketamine’s inhibition of NMDARs is not sufficient for its antidepressant actions. Direct administration of HNKs mimicked the behavioral actions of ketamine in mice, promoted EEG gamma oscillations, promoted expression of AMPARs and BDNF, and decreased phosphorylation of eEF2 levels in the hippocampus, providing plenty of mechanisms that could be triggered independently from NMDAR inhibition. Acute application of HNKs to hippocampal brain slices also produces a direct potentiation of SC-CA1 synaptic transmission [153], mediated by an increase in the probability of presynaptic glutamate release [161].
Perhaps the most direct evidence of the critical role that plasticity of excitatory synaptic transmission plays in the treatment of depression comes from preclinical studies of positive allosteric AMPAR modulators (i.e., AMPAkines) [162]. Antidepressant relevant effects reported for AMPAkines include decreased immobility in the FST, promotion of BDNF expression, amplification of ketamine effects, and increased neurogenesis in the dentate gyrus [163,164,165,166]. One AMPAkine failed to restore sucrose preference in chronically stressed mice, however [165].
The prefrontal cortices
Prefrontal cortical regions act in concert with the hippocampus, to which they are coupled by direct synaptic projections. For simplicity in this review, I will refer to the dorsal and ventral prefrontal cortices, the infralimbic cortex, and the orbitofrontal cortex as prefrontal cortex (PFC), but it is important to recognize that there is strong evidence that their functions are not identical and may display sex differences [167].
An essential adaptive skill of all animals is to learn to associate a rewarding experience with the sensory cues and actions in the environment that can be used to predict their future occurrence. This learning skill allows us to make optimal decisions and take appropriate approach and consummatory actions. In order to do this, we need an internal representation of the environment and of expected rewards that are based on prior experiences and predictive cues. These cognitive maps of rewards underlie all goal-directed behaviors, providing an adaptive benefit of permitting flexible decision-making optimized for both interoceptive state and environmental availability. Similarly, it is essential to have experience-dependent, up-datable knowledge about where danger lurks or what foods are unpalatable in the environment. There is excellent evidence that these positively and negatively valenced functions take place within the PFC [168,169,170].
It is becoming increasingly apparent that not only is the physical environment represented within the prefrontal cortices, but also the social environment. Our own experiences tell us that the social environment may be even more dynamic than the physical environment and certainly more arbitrary. Included in the social environment are various features such as cooperativity, competition, and social affiliation that are strongly correlated with the triggers and symptoms of depression. Loneliness, for example, is caused by perceived social isolation and absence of companionship and is associated with increased risk for depression and suicide [171]. On the other hand, social support is one of the strongest predictors of resilience and increasing social support can provide a beneficial component of effective therapeutic approaches [172].
The cellular mechanisms underlying the PFC’s ability to perform these functions are recently becoming clearer. It appears that the PFC acts as a choice option comparator during reward-guided decision making. Electrophysiological activity in the PFC encodes potential choice options and computes their comparison in the currently relevant environment. PFC output then turns these computations into actual actions [170]. In the human brain, activity increases as cues and stimuli become relevant to decisions, with the magnitude of this increase directly proportional to the outcome and reliability of the decision [173]. Modeling studies suggest that these comparisons are performed by two competing populations of neurons, each representing one option, which compete in a winner-take-all manner for choice via mutual recurrent excitatory synapses and reward-dependent synaptic plasticity [174, 175]. Deficits in PFC synaptic excitation and imbalances between excitation and inhibition would have obvious impact on these calculations.
The PFC is also an important site at which cognitive evaluations, such as the controllability of a stressor [176] or the valence of sensory stimuli [177], can influence affect and reward directly. It also regulates the negative ruminative thinking that can indirectly have an adverse impact on cognition and mood [178]. For example, cognitive control of responses to emotionally laden stimuli activates lateral regions of the PFC in human subjects [179].
Measured with fMRI, the PFC of depressed patients displays evidence of reduced volume, activity, and connectivity. Activity in the human PFC is increased by ‘happy’ stimuli to a greater extent than it is for ‘sad’ stimuli and these responses are altered in depressed and anhedonic patients [180,181,182], consistent with alterations in PFC circuit function. Hypoactivation of these regions is observed in MDD patients when performing tasks that require ignoring negative valence distracters [183]. Similarly, Liston et al. [184] tested heathy subjects in a PFC-dependent attention-shifting task before and one month after exposing them to psychosocial stress. Compared to a group of unstressed control subjects, stress selectively impaired subjects’ attention and disrupted functional connectivity within a frontoparietal cortical network. Comparable effects are seen in a decision-making task, in which stress favors habitual choices.
In animal models, chronic stress impairs attentional set-shifting tasks that are mediated by the PFC and decision making in reward-based tasks [185], mimicking the human deficits seen in MDD patients [184]. Chronic stress also results in a ca. 20% reduction in the length of apical dendrites in pyramidal neurons in the PFC and a ca. 30% decrease in the density of axospinous synapses on apical dendrites, along with corresponding decreases in the expression of synaptic genes [186,187,188,189]. Longitudinal two-photon microscopic imaging of fluorescently labeled dendritic spines in the PFC revealed that chronic elevation of CORT leads to an increase in dendritic spine turnover and an abnormal loss of stable spines that had been formed early in life [190].
Loss of dendritic spines suggests a loss or dysfunction of excitatory synapses. Indeed, consistent with spine loss, chronic stress decreases the synaptic excitation of pyramidal cells in the PFC [191]. Both AMPAR- and NMDAR- components of the EPSP were equally affected, unlike in the NAc [192, 193] and hippocampus [110]. In parallel to these physiological findings, expression of GluA1 and GluN1 proteins was decreased due to ubiquitination and proteosomal degradation. There was no change in expression of the postsynaptic density protein PSD95, however. Consistent with the decrease in excitation, expression of several immediate early genes and genes associated with synaptic function are decreased in the PFC of depressed humans and in rodent models [194, 195].
There is strong evidence that treatments that reverse the symptoms of depression also reverse deficits in the PFC. Stress induced decision-making deficits in MDD patients are reversible: after one month of reduced stress, subjects were indistinguishable from controls [196]. Treatment of MDD patients with classical monoaminergic antidepressants also restores normal activation of the PFC in attention tasks and restores the volume of the PFC [197,198,199].
There is also strong preclinical evidence that increases in PFC activity mimic the actions of antidepressants. For example, optogenetic stimulation of the mPFC in a bursting pattern, not unlike that occurring during ketamine-induced gamma oscillations, in mice subjected to chronic social defeat drives changes in immediate early gene expression like those seen in the PFC of human patients medicated with antidepressants and has an antidepressant-like effect in sucrose and social interaction preference tests [194]. These findings are currently being translated into novel clinical therapies in which invasive deep brain stimulation or noninvasive transcranial magnetic stimulation targeted to the PFC or its descending output pathways is used to treat patients with depression [200, 201].
Ketamine and other fast-acting antidepressant candidates all produce a rapid restoration of dendritic spine density in the PFC in stressed and unstressed animals with the same rapid time course as its behavioral actions [145, 187, 202, 203], implying the restoration of synaptic and circuit function, too. Considerable progress has been made in identifying the cellular signaling mechanisms underlying these effects. One strong candidate is the activation of mammalian target of rapamycin (mTOR), a central regulator of cellular growth [202]. Ketamine triggers not only its activation, but also the activation of many of its downstream targets, within minutes-to-hours, in unstressed rats. Furthermore, co-administration of the mTOR inhibitor rapamycin prevented the induction of dendritic spines by ketamine, as well as ketamine’s actions in the FST and other behaviors [202]. Use of synaptic vesicle PET ligands has recently revealed that ketamine produces similar synapse-promoting effects in the PFC in human depression [204].
Immunohistochemical analysis of interneurons and transcripts of interneuron-associated proteins are decreased within the PFC and anterior cingulate cortex in human MDD brain tissue and in chronically stressed rats, suggesting that deficits in GABA levels may contribute to the symptoms of depression [205,206,207,208]. Somatostatin- and parvalbumin-expressing GABAergic interneurons are particularly affected in chronically stressed rats [209]. There is also evidence that the antidepressant effects of ketamine occur via inhibition of GABA interneurons. As in the hippocampus, ketamine inhibits the discharge of interneurons in the PFC and reduces spontaneous IPSCs in pyramidal cells acutely [210], while restoring inhibition at later timepoints corresponding to its behavioral actions [211]. Furthermore, knock-down and deletion of critical ketamine-sensitive NMDARs from either somatostatin- or parvalbumin-expressing interneurons alters baseline behaviors in the FST and other tasks and eliminates the ability of ketamine to change these behaviors in unstressed mice [210]. The effects of the deletion on the ability of ketamine to produce electrophysiological disinhibition were not tested, unfortunately. Similarly, chemogenetic inhibition of either somatostatin- and parvalbumin-expressing interneurons in the mPFC mimicked the effects produced by antidepressants in the FST and other tasks, suggesting that transient inhibition of GABAergic interneurons contributes to the actions of rapidly acting antidepressants [212].
As discussed above, inhibition of interneurons may promote the kinds of high frequency oscillatory activity in pyramidal cells that underlies gamma oscillations, as seen in human and rodent EEGs, and, in turn, promotes endogenous activity-dependent mechanisms that strengthen stress-impaired excitatory synapses, such as LTP and BDNF-trkB signaling. Indeed, ketamine and GABAA receptor negative allosteric modulators share an ability to promote gamma oscillations in the PFC at doses that reverse the behavioral consequences of chronic stress [152, 153]. Many of these forms of activity-dependent plasticity are likely to be NMDAR-dependent, as supported by the antidepressant-like actions of the NMDAR positive allosteric modulator GLYX-13 (rapastinel) in preclinical models [213]. Ketamine’s disinhibitory action in dendrites also promotes dendritic Ca2+ influx [214], providing an additional mechanism to drive endogenous activity-dependent synaptic strengthening mechanisms.
The nucleus accumbens
The nucleus accumbens in the ventral striatum is an essential integrator of input pathways from the prefrontal cortex, hippocampus, and amygdala and is critical for determining motivating and reinforcing responses to rewarding stimuli and avoidance responses to aversive stimuli [215,216,217]. The NAc is divided into two histologically distinct subregions, the core and the shell, which have unique patterns of inputs and outputs [218, 219] and are thought to serve distinct, but similar functions [177]. The NAc consists primarily of GABAergic medium spiny neurons (MSNs), which can be distinguished into two primary populations based on their largely non-overlapping expression of dopamine receptors. As originally described in the dorsal striatum, MSNs expressing D1-type dopamine receptors, canonically considered the ‘direct’ pathway, project to and inhibit GABAergic interneurons within the VTA that, in turn, inhibit VTA dopamine releasing cells. Activation of D1-expressing MSNs thereby promotes activation of dopamine projections back to the forebrain by disinhibiting them [220]. In contrast, MSNs expressing D2-type dopamine receptors, canonically considered the ‘indirect’ pathway, project to GABAergic cells within the ventral pallidum, that then project to the interneurons within the VTA. Activation of D2-expressing MSNs thereby inhibits activation of dopamine projections back to the forebrain by disinhibiting the interneurons that silence them. More recent findings have established that these neat divisions do not apply as simplistically to the ventral striatum [221, 222]. Nevertheless, activity in D1 MSNs is more likely to be associated with motivation, reward, and positive responses [193, 223,224,225] and resilience to stress [226]. Conversely, a decrease in the ability of D1 MSNs to drive dopamine release from the VTA, due to decreased excitation and/or increased activity of D2 MSNs, may contribute to the loss of the rewarding properties of normally rewarding stimuli characterizing anhedonia and lack of motivation in the depressed state.
Activation of inputs to the NAc from the hippocampus and PFC is rewarding and reinforcing of rewarding stimuli [227, 228]. Excitatory synapses in the NAc in these pathways display activity-dependent plasticity. Much like canonical hippocampal LTP, high frequency stimulation of hippocampal afferents to the NAc elicits a rapid and persistent NMDAR-triggered, Ca2+- and CaMK-dependent increase in postsynaptic AMPAR responses in D1 and D2 MSNs [193]. Dopamine receptor activation is not required for this potentiation but may be modulatory under physiological conditions [229]. Activity-dependent LTP in vivo was both necessary and sufficient to induce a conditioned place preference lasting for at least 24hrs [193]. Silencing hippocampal inputs to the NAc prevented formation of a place preference following social interactions, as assayed one day later, but did not prevent the rewarding properties of the social interaction itself. Similarly, establishment of cocaine place preferences is accompanied by a potentiation of hippocampal-NAc synaptic excitation [230]. Conversely, weakening inputs to the NAc can reverse previously established reward responses to addictive drugs [231, 232].
In response to chronic stress, rodent NAc MSNs display an array of plastic changes that act in concert to alter forebrain dopaminergic release from the VTA and, thereby, alter the responses to rewarding and aversive stimuli [233]. As such they are heavily implicated as central players in the genesis of depression. Susceptibility to chronic social defeat stress is accompanied by atrophy of MSN dendrites, specifically in D1 MSNs and not D2 MSNs, and a corresponding loss of dendritic spines [226, 234, 235]. Consistent with the anatomical findings, the frequency of miniature EPSCs is decreased without a change in amplitude. Similarly, evoked AMPA receptor-mediated excitatory synaptic currents are reduced by chronic stress in D1, but not D2, MSNs [236], including in response to afferents from the hippocampus [193]. These decreases in the excitation of D1 MSNs may be sufficient, in and of themselves, to mediate an anhedonic loss of reward behavior. Lim et al. [236] demonstrated that preventing stress-induced AMPA receptor loss in D1 MSNs was sufficient to prevent stress-induced loss of hedonic responses in sucrose preference and cocaine-induced place preference (although not the FST).
Chronic stress also impairs hippocampal – NAc LTP in D1 MSNs, but not D2 MSNs. This differential effect suggests a possible stress-induced biasing of NAc responses from D1 MSN towards D2 MSN activation, hence favoring negative valence responses over positive valence. Neuromodulators, including dopamine itself, serotonin, and enkephalins are also likely to play a significant role in gating synaptic strength and the balance between D1 and D2 MSN activity [217, 233, 237]. The intrinsic excitability of NAc MSNs is also affected by stress. Susceptibility to social defeat stress is accompanied by an increase in input resistance and firing in response to depolarizing currents pulses in D1 MSNs, but not D2 MSNs [226, 234].
Although receiving less attention than cortical plasticity, there is evidence that known and novel potential antidepressant compounds exert some of their therapeutic benefits via actions within the NAc. Chronic, but not acute, administration of fluoxetine to stressed mice restores the strength of hippocampal excitatory synapses in D1 MSNs and their ability to undergo LTP, concomitant with restoration of sucrose preference [193]. Consistent with these synaptic findings, the probability of action potential firing of neurons in the NAc in response to stimulation of hippocampal afferents is increased by escitalopram administration [238]. The net effect of these synaptic changes is to restore the function of the NAc- VTA circuit. Chronic treatment with escitalopram restores the ability of rewarding stimuli to trigger DA release within the NAc in chronically stressed rats [239]. Almost nothing is known about the mechanisms underlying the restorative actions of SSRIs in the NAc.
Ketamine also has effects on NAc synaptic function, including a direct potentiation of PFC-NAc synapses. Ketamine also restores the ability to induce LTP in the hippocampus-accumbens pathway in stressed animals [240]. Restoration of synaptic function is accompanied by restoration of circuit function. Ketamine restores a stress-induced decrease in VTA dopaminergic cell firing [240] and increases extracellular levels of dopamine in the nucleus accumbens [241]. The mechanisms underlying these effects remain unknown. Like the effects of ketamine in the cortex, ketamine increases the power of high-frequency oscillations of local field potentials in the NAc in unanesthetized mice [242], suggesting a potential role for activity-dependent synapse strengthening mechanisms. Given the lack of local excitatory interactions within the NAc, it is likely that this high frequency synchronized activity is projected onto the NAc via afferents from the PFC and hippocampus. As elsewhere in the brain, elevated BDNF – trkB signaling may be crucial within these mesolimbic circuits [243]. Finally, local infusion of ketamine into the NAc reversed stress-induced decreases in the phosphorylation of GluA1 AMPARs at serines 831 and 845, which is known to promote their insertion into synapses, suggesting a direct mechanism for the potentiation of NAc synapses [244].
Increases in resolution and increased use of reward-based tasks that correlate with preclinical studies have enhanced the ability to resolve the NAc in human functional imaging studies and allowed for new insights into the role its plasticity plays in reward deficits in human depression. There is evidence that the NAc is activated by reward anticipation and both motivational and hedonic aspects of reward processing [245]. Defects in hippocampal – NAc coupling have been observed to correlate with reward processing deficits [246]. Unmedicated MDD patients show deficits in reward learning, reduced responses in the ventral striatum for positive prediction errors in a reward task, and reduced NAc- VTA connectivity [247, 248]. There is limited evidence of the effect of antidepressants on these measures of reward circuit function, but it has been shown that improvements in self-reported depression severity and negative affective bias are improved with chronic SSRI administration in depressed patients and are strongly correlated with increases in NAc activity and NAc- PFC connectivity [249]. Similar increases in NAc-PFC connectivity correlate with reduced symptom severity and quality-of-life measurements in SSRI-responsive patients compared to unresponsive controls [250]. More studies are clearly needed. In particular, direct before-and-after comparisons of patients receiving fast-acting antidepressants would be of great interest. For example, Sterpinich et al. [251] observed that ketamine increased responses in a reward task and caused an accompanying increase in activity in the orbitofrontal cortex, ventral striatum, and the ventral tegmental area. Importantly, these effects persisted for up to a week after a single ketamine administration, strongly supporting an important role for plasticity in the cortico-mesolimbic circuitry in the genesis and treatment of depression.
The lateral habenula
The habenula is an epithalamic nucleus comprised of medial and lateral subregions. The lateral habenula (LHb) is another key node in regulation of reward behaviors that has been implicated in the genesis of depressive symptoms and reward dysregulation [252]. It integrates glutamatergic inputs from the lateral hypothalamus [253] and from the anterior cingulate and medial prefrontal cortices [254]. Excitatory synapses onto LHb cells are characterized by a high ratio of AMPAR- to NMDAR-mediated synaptic responses [255], indicative of low levels of NMDAR expression. LHb neurons also receive feed-forward and feedback GABAergic inhibition [253, 255]. The glutamatergic neurons of the LHb have three primary targets, the dopaminergic cells of the VTA, the serotonergic neurons of the nucleus raphe, and the GABAergic neurons of the rostromedial tegmental nucleus (RMTg). RMTg neurons, in turn, project to the VTA and are largely responsible for the predominantly inhibitory action of LHb projections onto VTA dopaminergic neurons [256].
Activity of LHb neurons is generally correlated with aversive and negative affective behaviors. LHb neurons fire in response to both aversive stimuli and the absence of an expected reward [257,258,259,260]. Conversely, reward delivery decreases LHb discharge [257]. Optogenetic activation of LHb neurons via projections from the lateral hypothalamic nucleus (LH) triggers avoidance and escape behaviors, whereas optogenetic or pharmacological inhibition of LHb cell discharge impairs avoidance and escape in response to aversive stimuli [258,259,260]. With regard to behaviors typically used to study depression and antidepressant drugs, acute physical restraint, foot shock, tail suspension, forced swimming, and social defeat are all accompanied by strong LHb activation [258, 260]. The consequences of LHb activity are largely mediated by its output to the RMTg [261]. Importantly, activation of LHb inputs to the RMTg reduced motivation to work for a rewarding sucrose solution in an operant task, without reducing its rewarding hedonic properties [262].
There have now been several demonstrations that chronic stress results in changes in LHb discharge and synaptic function. Mechanisms include both changes in the intrinsic excitability of LHb neurons and plasticity of their synapses. Stress-induced increases in firing are attributed to decreased function of G-protein coupled K+ channels in LHb neurons [263]. Increased burst discharges [264] are attributed to dysregulation of the astrocytic K+ channels that regulate extracellular K+ concentrations, thereby promoting hyperpolarization of LHb neurons [265], although this hyperpolarization was not shown. Increased numbers of burst-firing LHb neurons were observed in vitro and in vivo in both a rat strain with congenital learned helplessness and in wild type rats after chronic restraint stress, although no overall change in membrane potential was reported [266]. Bursting in LHb cells is mediated by T-type voltage-dependent Ca2+ channels and NMDA receptors. Surprisingly, bursting was largely unaffected by AMPAR antagonists, suggesting that synaptic activity does not drive bursting. Optogenetically induced increases in burst firing in LHb neurons increased mobility in the FST and decreased sucrose preference [266], like the effects of chronic stress.
Chronic stress potentiates excitatory transmission onto LHb cells in parallel with altered excitability. An increased probability of presynaptic glutamate release in the LHb accompanies learned helplessness and chronic restraint stress [255, 260]. Some forms of chronic stress also increase the amplitude of spontaneous excitatory synaptic responses [260]. Selective activity-dependent potentiation of synapses formed by axons of LH neurons with LHb neurons promoted LHb discharge and was sufficient to induce mimic the behavioral effects of chronic stress in the forced swim and sucrose preference tests [260, 267].
These behavioral results, although demanding to demonstrate, can be predicted from previous evidence that LHb firing is aversive. So, does the increase in LHb bursting drive behavioral changes in response to chronic stress? Lesioning or pharmacologically silencing the LHb can normalize learned helplessness responses [266, 268], including inhibition of NMDAR-mediated LHb bursting [264]. Evidence that chemo- or optogenetic silencing of LHb neurons can prevent pathological behaviors in a chronic stress paradigm is lacking. Viral knockdown of beta-CaMK, the enzyme underlying the stress-induced presynaptic potentiation of LHb excitatory synapses, prevented forced swim and learned helplessness behaviors in congenitally learned helpless rats [267]. Whether prevention of activity-dependent potentiation of LH-LHb synapses would block behavioral changes in response to chronic stress remains to be demonstrated. Nevertheless, the evidence is plentiful and strong that the LHb is an important mediator of aversive behavioral responses via its outputs to the VTA and that its behavior is profoundly increased in a variety of chronic stress conditions used to model depression relevant behaviors in rodents (for excellent further reviews, see: [252, 269, 270]).
Future research directions: putting it all together
The work cited above, describing results from both preclinical and human studies, provides compelling evidence, first, that prolonged stress, both physical and psychosocial, affects the structure, protein composition, and function of excitatory synapses in every area of the brain devoted to the processing of reward related stimuli and cognitive, learning, and memory processes. These plastic changes appear maladaptive. Excitatory synapses generally become reduced in number and weaker in areas devoted to directing attention and making decisions about cues and stimuli and stronger in the pathways processing negative valence stimuli. These synapses are the working elements of the circuits they are embedded in and hence are could be the cause of significant changes in the function of those circuits. In general, the synaptic changes result in a weakening of pathways that promote dopamine release by activating cells in the VTA, such as the direct and indirect weakening of excitatory drive of D1 MSNs in the NAc, and potentiation of pathways inhibiting dopamine secretion, such as those within the LH-LHb-RMTg.
These concerted changes in synapses in response to chronic stress have been found in essentially every circuit in every reward pathway, providing a mechanistic explanation for the wide range of symptoms that together constitute the clinical definition of depression. As summarized above, there is strong evidence from imaging and functional connectivity studies of human patients to suggest that analogous changes occur in human MDD. Of course, not all patients display the same combination of symptoms. Perhaps the relative prevalence of individual symptoms in a given MDD patient arises from the relative degree to which any particular synapses and circuits are affected in that patient. Considering emotional dysregulation [271], for example, an overactive LHb circuit may favor an increase in negative affect. In contrast, hypoactive PFC circuits might be more heavily affected in patients with more prevalent cognitive and decision-making symptoms. A better understanding of the function of these reward circuits in preclinical models and human subjects and their dysfunction in depression may thus help to distinguish between subtypes of depression [2]. Because not all circuits and symptoms may respond in the same manner to all antidepressants, a better understanding may also guide the choice of antidepressant therapies to employ. For example, increased negative affective bias in depressed subjects was shown to respond rapidly to a norepinephrine reuptake inhibitor, even though patients reported no overall improvement in their overall depression [272]. The distributed nature of these changes also complicates experimental efforts to identify a single synapse or circuit as the unique ‘cause’ of a given behavioral deficit, given their highly interconnected nature. Driving the LHb may overpower counterbalancing NAc inputs to the VTA, for example, regardless of whether they are fully functional or dysfunctional.
An underlying assumption of all preclinical models is that the forms of chronic stress we produce experimentally in our animals provides a realistic model of human depression. On the one hand, the evidence of the etiological relevance of stressful life events to human depression, either acutely or during critical developmental windows, is very strong. On the other hand, environmental factors are believed to account for only 60% of risk for MDD. Preclinical studies are largely done on experimental animals with a relatively homogenous genetic background, obscuring the important contributions of genetics to susceptibility and resilience. Given the likelihood that overall genetic risk results from small differences in large numbers of genes, each with small contributions to overall risk, this blind spot represents a particular short coming in our current understanding and significant experimental challenge for future progress.
Supporting the hypothesis that synaptic and circuit dysfunction is a major cause of depression, there is increasing evidence that restoration of the function of synapses and circuits is a common mechanism of action underlying a wide range of known and novel antidepressants. Those antidepressants that restore synaptic strength slowly, such as SSRIs, restore synaptic strength slowly, whereas fast acting antidepressants do so rapidly. Further supporting this hypothesis of antidepressant action, these substances do not necessarily share the same mechanisms of action. Although there is overlap, some favor activity-dependent synaptic strengthening mechanisms, such as ketamine and ketamine metabolites, whereas others, such as SSRIs, favor neurotrophic growth factor signaling. Nevertheless, all strengthen synapse function. The search for novel antidepressants should be focused on a search for compounds that share this mechanism of action. For example, we hypothesized that negative allosteric modulators of GABAA receptors containing alpha5 subunits would promote a transient, ketamine-like increase in high frequency, synchronized oscillations in the PFC and hippocampus and would therefore produce a ketamine-like rapid antidepressant response. Subsequent preclinical experiments supported this hypothesis [152, 273].
One of the most exciting developments in the treatment of depression and other psychiatric disorders has been the recent findings that a single administration of psilocybin produces a rapid and persistent improvement in self-reported depression symptoms in patients resistant to conventional antidepressants (e.g., [274]). There is also robust preclinical evidence that psilocybin can restore both reward behaviors and synapse structure and function. In the hippocampus, stress-impaired reward behaviors are restored to baseline levels 24 h after a single administration of psilocybin [275], exactly as seen with other fast-acting antidepressants. Taking hippocampal brain slices from these same mice revealed that psilocybin also restored the strength of the stress-sensitive TA-CA1 synapses, concurrent with the restoration of reward behavior, again much like other fast-acting antidepressants. Similarly, in the PFC, psilocybin administration to unstressed mice leads to an increase in the net formation of new dendritic spines in pyramidal cells, persisting for over one month, which is accompanied by an increase in the frequency of spontaneous mEPSCs [276]. These preclinical findings are paralleled by human functional imaging findings that psychedelics change connectivity in reward- and emotion-relevant brain regions in a persistent manner [277]. Surprisingly, both studies suggested that the actions of psilocybin were independent of the serotonin 2 A receptor that mediates the mind-altering properties of psychedelics, raising the possibility of dissociating the ‘trip’ from the antidepressant benefits of psychedelics. Because of the expense and time-consuming nature of current psychedelic therapies, involving multiple clinic visits and a full day with one or more therapists, this could considerably lower barriers to widespread use. An active current area of research is to define the expression mechanisms underlying the persistent increases in synaptic strength. Many of the same processes described above are likely to be involved, including neurotrophin signaling and enzymatic cascades promoting receptor trafficking and process outgrowth [278]. It will be of great interest to further identify and refine the beneficial properties of these powerful compounds and determine whether their promise is mediated by promoting plasticity of synapses and reward circuits.
References
Drysdale AT, Grosenick L, Downar J, Dunlop K, Mansouri F, Meng Y, et al. Resting-state connectivity biomarkers define neurophysiological subtypes of depression. Nat Med. 2017;23:28–38.
Buch AM, Liston C. Dissecting diagnostic heterogeneity in depression by integrating neuroimaging and genetics. Neuropsychopharmacol. 2021;46:156–75.
Nestler EJ, Barrot M, DiLeone RJ, Eisch AJ, Gold SJ, Monteggia LM. Neurobiology of depression. Neuron. 2002;34:13–25.
Fava M, Kendler KS. Major depressive disorder. Neuron. 2000;28:335–41.
Wray NR, Ripke S, Mattheisen M, Trzaskowski M, Byrne EM, Abdellaoui A, et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat Genet. 2018;50:668–81.
Billings AG, Cronkite RC, Moos RH. Social-environmental factors in unipolar depression: comparisons of depressed patients and nondepressed controls. J Abnorm Psychol. 1983;92:119–33.
Bale TL. Epigenetic and transgenerational reprogramming of brain development. Nat Rev Neurosci. 2015;16:332–44.
Hanson JL, Williams AV, Bangasser DA, Peña CJ. Impact of early life stress on reward circuit function and regulation. Front Psychiatry. 2021;12:744690.
Nestler EJ. Role of the brain’s reward circuitry in depression: transcriptional mechanisms. Int Rev Neurobiol. 2015;124:151–70.
Morgan CP, Chan JC, Bale TL. Driving the next generation: paternal lifetime experiences transmitted via extracellular vesicles and their small RNA cargo. Biol Psychiatry. 2019;85:164–71.
de Pablos RM, Villarán RF, Argüelles S, Herrera AJ, Venero JL, Ayala A, et al. Stress increases vulnerability to inflammation in the rat prefrontal cortex. J Neurosci. 2006;26:5709–19.
Sanacora G, Treccani G, Popoli M. Towards a glutamate hypothesis of depression: an emerging frontier of neuropsychopharmacology for mood disorders. Neuropharmacol. 2012;62:63–77.
Thompson SM, Kallarackal AJ, Kvarta MD, Van Dyke AM, LeGates TA, Cai X. An excitatory synapse hypothesis of depression. Trends Neurosci. 2015;38:279–94.
Gould TD, Zarate CA Jr, Thompson SM. Molecular pharmacology and neurobiology of rapid-acting antidepressants. Annu Rev Pharm Toxicol. 2019;59:213–36.
Duman RS, Sanacora G, Krystal JH. Altered connectivity in depression: GABA and glutamate neurotransmitter deficits and reversal by novel treatments. Neuron. 2019;102:75–90.
Sanacora G, Yan Z, Popoli M. The stressed synapse 2.0: pathophysiological mechanisms in stress-related neuropsychiatric disorders. Nat Rev Neurosci. 2022;23:86–103.
Holsboer F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacol. 2000;23:477–501.
Xia CH, Ma Z, Ciric R, Gu S, Betzel RF, Kaczkurkin AN, et al. Linked dimensions of psychopathology and connectivity in functional brain networks. Nat Commun. 2018;9:3003.
Nestler EJ, Carlezon WA. The mesolimbic dopamine reward circuit in depression. Biol Psychiatry. 2006;59:1151–9.
Tye KM, Mirzabekov JJ, Warden MR, Ferenczi EA, Tsai HC, Finkelstein J, et al. Dopamine neurons modulate neural encoding and expression of depression-related behaviour. Nature. 2013;493:537–41.
Burke CJ, Dreher J-C, Seymour B, Tobler PN. State-dependent value representation: evidence from the striatum. Front Neurosci. 2014;8:1481–9.
Polter AM, Kauer JA. Stress and VTA synapses: implications for addiction and depression. Eur J Neurosci. 2014;39:1179–88.
Grace AA. Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat Rev Neurosci. 2016;17:524–32.
Floresco SB, Magyar O. Mesocortical dopamine modulation of executive functions: beyond working memory. Psychopharmacol. 2006;188:567–85.
Treadway MT, Zald DH. Reconsidering anhedonia in depression: lessons from translational neuroscience. Neurosci Biobehav Rev. 2011;35:537–55.
Schultz W. Neuronal reward and decision signals: from theories to data. Physiol Rev. 2015;95:853–951.
Berridge KC. Evolving concepts of emotion and motivation. Front Psychol. 2018;9:1647.
Thiele A, Bellgrove MA. Neuromodulation of Attention. Neuron. 2018;97:769–85.
Walton ME, Bouret S. What is the relationship between dopamine and effort? Trends Neurosci. 2019;42:79–91.
Krishnan V, Nestler EJ. Animal models of depression: molecular perspectives. Curr Top Behav Neurosci. 2011;7:121–47.
Willner P. The chronic mild stress (CMS) model of depression: history, evaluation and usage. Neurobiol Stress. 2017;6:78–93.
Lopez J, Bagot RC. Defining valid chronic stress models for depression with female rodents. Biol Psychiatry. 2021;90:226–35.
Herman JP. Neural control of chronic stress adaptation. Front Behav Neurosci. 2013;7:61.
Birnie MT, Kooiker CL, Short AK, Bolton JL, Chen Y, Baram TZ. Plasticity of the reward circuitry after early-life adversity: mechanisms and significance. Biol Psychiatry. 2020;87:875–84.
Gourley SL, Kiraly DD, Howell JL, Olausson P, Jane R, Taylor JR. Acute hippocampal BDNF restores motivational and forced swim performance after corticosterone. J Neurosci. 2009;64:884–90.
Kvarta MD, Bradbrook KE, Dantrassy HM, Bailey AM, Thompson SM. Corticosterone mediates the synaptic and behavioral effects of chronic stress at rat hippocampal temporoammonic synapses. J Neurophysiol. 2015;114:1713–24.
Strekalova T, Steinbusch HW. Measuring behavior in mice with chronic stress depression paradigm. Prog Neuropsychopharmacol Biol Psychiatry. 2010;34:348–61.
Lammel S, Tye KM, Warden MR. Progress in understanding mood disorders: optogenetic dissection of neural circuits. Genes Brain Behav. 2014;13:38–51.
Price RB, Gates K, Kraynak TE, Thase ME, Siegle GJ. Data-driven subgroups in depression derived from directed functional connectivity paths at rest. Neuropsychopharmacol. 2017;42:2623–32.
Papp M, Moryl E, Willner P. Pharmacological validation of the chronic mild stress model of depression. Eur J Pharm. 1996;296:129–36.
Grippo AJ, Beltz TG, Weiss RM, Johnson AK. The effects of chronic fluoxetine treatment on chronic mild stress-induced cardiovascular changes and anhedonia. Biol Psychiatry. 2006;59:309–16.
Bondi CO, Rodriguez G, Gould GG, Frazer A, Morilak DA. Chronic unpredictable stress induces a cognitive deficit and anxiety-like behavior in rats that is prevented by chronic antidepressant drug treatment. Neuropsychopharmacol. 2008;33:320–31.
Gourley SL, Taylor JR. Recapitulation and reversal of a persistent depression-like syndrome in rodents. Curr Protoc Neurosci. 2009, Chapter 9, 9; Unit 9.32.
Cai X, Kallarackal AJ, Kvarta MD, Goluskin S, Gaylor K, Bailey AM, et al. Local potentiation of excitatory synapses by serotonin and its alteration in rodent models of depression. Nat Neurosci. 2013;16:464–72.
Zanos P, Thompson SM, Duman RS, Zarate CA, Gould TD. Convergent mechanisms underlying rapid antidepressant action. CNS Drugs. 2018;32:197–227.
Bale TL, Abel T, Akil H, Carlezon WA Jr, Moghaddam B, Nestler EJ, et al. The critical importance of basic animal research for neuropsychiatric disorders. Neuropsychopharmacol. 2019;44:1349–53.
Treadway MT, Buckholtz JW, Schwartzman AN, Lambert WE, Zald DH. Worth the ‘EEfRT’? The effort expenditure for rewards task as an objective measure of motivation and anhedonia. PLoS One. 2009;4:e6598.
Lewis LR, Benn A, Dwyer DM, Robinson ESJ. Affective biases and their interaction with other reward-related deficits in rodent models of psychiatric disorders. Behav Brain Res. 2019;372:112051.
Pizzagalli DA. Toward a better understanding of the mechanisms and pathophysiology of anhedonia: are we ready for translation? Am J Psychiatry. 2022;179:458–69.
Sesack SR, Grace AA. Cortico-basal ganglia reward network: microcircuitry. Neuropsychopharmacol. 2010;2010:27–47.
Russo SJ, Nestler EJ. The brain reward circuitry in mood disorders. Nat Rev Neurosci. 2013;14:609–25.
Lammel S, Lim BK, Malenka RC. Reward and aversion in a heterogeneous midbrain dopamine system. Neuropharmacol. 2014a;76:351–9.
Belujon P, Grace AA. Dopamine system dysregulation in major depressive disorders. Int J Neuropsychopharmacol. 2017;20:1036–46.
Tye KM. Neural circuit motifs in valence processing. Neuron. 2018;100:436–52.
Berridge KC. Affective valence in the brain: modules or modes? Nat Rev Neurosci. 2019;20:225–34.
Ottowitz WE, Dougherty DD, Savage CR. The neural network for abnormalities of attention and executive function in major depressive disorder: implications for application of the medical disease model to psychiatric disorders. Harv Rev Psychiatry. 2002;10:86–99.
Frodl T, Schaub A, Banac S, Charypar M, Jäger M, Kümmler P, et al. Reduced hippocampal volume correlates with executive dysfunctioning in major depression. J Psych Neurosci. 2006;31:316–23.
Sosa M, Giocomo LM. Navigating for reward. Nat Rev Neurosci. 2021;22:472–87.
Eichenbaum H, Kuperstein M, Fagan A, Nagode J. Cue- sampling and goal- approach correlates of hippocampal unit- activity in rats performing an odor- discrimination task. J Neurosci. 1987;7:716–32.
Nyberg N, Duvelle É, Barry C, Spiers HJ. Spatial goal coding in the hippocampal formation. Neuron. 2022;110:394–422.
Dupret D, O’Neill J, Pleydell-Bouverie B, Csicsvari J. The reorganization and reactivation of hippocampal maps predict spatial memory performance. Nat Neurosci. 2010;13:995–1002.
Wikenheiser AM, Redish AD. Hippocampal theta sequences reflect current goals. Nat Neurosci. 2015;18:289–94.
Singer AC, Frank LM. Rewarded outcomes enhance reactivation of experience in the hippocampus. Neuron. 2009;64:910–21.
Roux L, Hu B, Eichler R, Stark E, Buzsaki G. Sharp wave ripples during learning stabilize the hippocampal spatial map. Nat Neurosci. 2017;20:845–53.
Engen HG, Anderson MC. Memory control: a fundamental mechanism of emotion regulation. Trends Cogn Sci. 2018;22:982–95.
McEvoy PM, Watson H, Watkins ER, Nathan P. The relationship between worry, rumination, and comorbidity: evidence for repetitive negative thinking as a transdiagnostic construct. J Affect Disord. 2013;151:313–20.
Anderson MC, Floresco SB. Prefrontal-hippocampal interactions supporting the extinction of emotional memories: the retrieval stopping model. Neuropsychopharmacology. 2022;47:180–95.
Phillips ML, Robinson HA, Pozzo-Miller L. Ventral hippocampal projections to the medial prefrontal cortex regulate social memory. Elife. 2019;8:e44182.
Sapolsky R, Krey L, McEwen BS. Prolonged glucocorticoid exposure reduces hippocampal neuron number: implications for aging. J Neurosci. 1985;5:1222–27.
Woolley C, Gould E, McEwen BS. Exposure to excess glucocorticoids alters dendritic morphology of adult hippocampal pyramidal neurons. Brain Res. 1990;531:225–31.
Watanabe Y, Gould E, McEwen BS. Stress induces atrophy of apical dendrites of hippocampal CA3 pyramidal neurons. Brain Res. 1992;588:341–5.
Sunanda, Rao MS, Raju TR. Effect of chronic restraint stress on dendritic spines and excrescences of hippocampal CA3 pyramidal neurons-a quantitative study. Brain Res. 1995;694:312–7.
Hajszan T, Dow A, Warner-Schmidt JL, Szigeti-Buck K, Sallam NL, Parducz A, et al. Remodeling of hippocampal spine synapses in the rat learned helplessness model of depression. Biol Psychiatry. 2009;65:392–400.
Magariños AM, McEwen BS, Flügge G, Fuchs E. Chronic psychosocial stress causes apical dendritic atrophy of hippocampal CA3 pyramidal neurons in subordinate tree shrews. J Neurosci. 1996;16:3534–40.
Silva-Gómez AB, Rojas D, Juárez I, Flores G. Decreased dendritic spine density on prefrontal cortical and hippocampal pyramidal neurons in postweaning social isolation rats. Brain Res. 2003;983:128–36.
Donohue HS, Gabbott PL, Davies HA, Rodríguez JJ, Cordero MI, Sandi C, et al. Chronic restraint stress induces changes in synapse morphology in stratum lacunosum-moleculare CA1 rat hippocampus: a stereological and three-dimensional ultrastructural study. Neuroscience. 2006;140:597–606.
Maras PM, Molet J, Chen Y, Rice C, Ji SG, Solodkin A, et al. Preferential loss of dorsal-hippocampus synapses underlies memory impairments provoked by short, multimodal stress. Mol Psychiatry. 2014;19:811–22.
Sheline YI, Wang PW, Gado MH, Csernansky JG, Vannier MW. Hippocampal atrophy in recurrent major depression. Proc Natl Acad Sci. 1996;93:3908–13.
Sheline YI, Sanghavi M, Mintun MA, Gado MH. Depression duration but not age predicts hippocampal volume loss in medically healthy women with recurrent major depression. J Neurosci. 1999;19:5034–43.
Videbech P, Ravnkilde B. Hippocampal volume and depression: a meta-analysis of MRI studies. Am J Psychiatry. 2004;161:1957–66.
Hickie I, Naismith S, Ward PB, Turner K, Scott E, Mitchell, et al. Reduced hippocampal volumes and memory loss in patients with early- and late-onset depression. Br J Psychiatry. 2005;186:197–202.
Starkman MN, Giordani B, Gebarski SS, Berent S, Schork MA, Schteingart DE. Decrease in cortisol reverses human hippocampal atrophy following treatment of Cushing’s disease. Biol Psychiatry. 1999;46:1595–602.
Opel N, Redlich R, Zwanzger P, Grotegerd D, Arolt V, Heindel W, et al. Hippocampal atrophy in major depression: a function of childhood maltreatment rather than diagnosis? Neuropsychopharmacol. 2014;39:2723–31.
Sheline YI, Gado MH, Kraemer HC. Untreated depression and hippocampal volume loss. Am J Psychiatry. 2003;160:1516–8.
Katsuki A, Watanabe K, Nguyen L, Otsuka Y, Igata R, Ikenouchi A, et al. Structural changes in hippocampal subfields in patients with continuous remission of drug-naive major depressive disorder. Int J Mol Sci. 2020;21:3032.
Zarate-Garza PP, Ortega-Balderas JA, Ontiveros-Sanchez de la Barquera JA, Lugo-Guillen RA, Marfil-Rivera A, Quiroga-Garza A, et al. Hippocampal volume as treatment predictor in antidepressant naïve patients with major depressive disorder. J Psychiatr Res. 2021;140:323–8.
Duman RS, Monteggia LM. A neurotrophic model for stress-related mood disorders. Biol Psychiatry. 2006;59:1116–27.
Dwivedi Y, Rizavi HS, Conley RR, Roberts RC, Tamminga CA, Pandey GN. Altered gene expression of brain-derived neurotrophic factor and receptor tyrosine kinase B in postmortem brain of suicide subjects. Arch Gen Psychiatry. 2003;60:804–15.
Tartt AN, Mariani MB, Hen R, Mann JJ, Boldrini M Dysregulation of adult hippocampal neuroplasticity in major depression: pathogenesis and therapeutic implications. Mol Psychiatry. 2022;27:2689-99; https://doi.org/10.1038/s41380-022-01520-y.
Gould E, McEwen BS, Tanapat P, Galea LA, Fuchs E. Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation. J Neurosci. 1997;17:2492–8.
Tanapat P, Galea LA, Gould E. Stress inhibits the proliferation of granule cell precursors in the developing dentate gyrus. Int J Dev Neurosci. 1998;16:235–9.
Joëls M, Karst H, Alfarez D, Heine VM, Qin Y, van Riel E, et al. Effects of chronic stress on structure and cell function in rat hippocampus and hypothalamus. Stress 2004;7:221–31.
Nibuya M, Morinobu S, Duman RS. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J Neurosci. 1995;15:7539–47.
Malberg J, Eisch AJ, Nestler EJ, Duman RS. Chronic antidepressant treatment increases neurogenesis in adult hippocampus. J Neurosci. 2000;20:9104–10.
Malberg J, Duman RS. Cell proliferation in adult hippocampus is decreased by inescapable stress: Reversal by fluoxetine treatment. Neuropsychopharmacol. 2003;28:1562–71.
Malberg JE, Hen R, Madsen TM. Adult neurogenesis and antidepressant treatment: the surprise finding by Ron Duman and the field 20 years later. Biol Psychiatry. 2021;90:96–101.
Duman RS, Li N. A neurotrophic hypothesis of depression: role of synaptogenesis in the actions of NMDA receptor antagonists. Philos Trans R Soc Lond B Biol Sci. 2012;367:2475–84.
Castrén E, Monteggia LM. Brain-derived neurotrophic factor signaling in depression and antidepressant action. Biol Psychiatry. 2021;90:128–36.
Dranovsky A, Hen R. Hippocampal neurogenesis: regulation by stress and antidepressants. Biol Psychiatry. 2006;59:1136–43.
Sahay A, Hen R. Adult hippocampal neurogenesis in depression. Nat Neurosci. 2007;10:1110–5.
Anacker C, Zunszain PA, Cattaneo A, Carvalho LA, Garabedian MJ, Thuret S, et al. Antidepressants increase human hippocampal neurogenesis by activating the glucocorticoid receptor. Mol Psychiatry. 2011;16:738–50.
Franjic D, Skarica M, Ma S, Arellano JI, Tebbenkamp ATN, Choi J, et al. Transcriptomic taxonomy and neurogenic trajectories of adult human, macaque, and pig hippocampal and entorhinal cells. Neuron. 2022;110:452–69.
Casarotto PC, Girych M, Fred SM, Kovaleva V, Moliner R, Enkavi G, et al. Antidepressant drugs act by directly binding to TRKB neurotrophin receptors. Cell. 2021;184:1299–313.
Krugers HJ, Hoogenraad CC, Groc L. Stress hormones and AMPA plasticity and memory. Nat Rev Neurosci. 2010;11:675–81.
Karst H, Joëls M. Effect of chronic stress on synaptic currents in rat hippocampal dentate gyrus neurons. J Neurophysiol. 2003;89:625–33.
Alfarez DN, Joëls M, Krugers HJ. Chronic unpredictable stress impairs long-term potentiation in rat hippocampal CA1 area and dentate gyrus in vitro. Eur J Neurosci. 2003;17:1928–34.
Airan RD, Meltzer LA, Roy M, Gong Y, Chen H, Deisseroth K. High-speed imaging reveals neurophysiological links to behavior in an animal model of depression. Science. 2007;317:819–23.
Sousa N, Lukoyanov NV, Madeira MD, Almeida OFX, Paula-Barbosa MM. Reorganization of the morphology of hippocampal neurites and synapses after stress-induced damage correlates with behavioral improvement. Neuroscience. 2000;97:253–66.
Kole MHP, Swan L, Fuchs E. The antidepressant tianeptine persistently modulates glutamate receptor currents of the hippocampal CA3 commissural associational synapse in chronically stressed rats. Eur J Neurosci. 2002;16:807–16.
Kallarackal AJ, Kvarta MD, Cammarata E, Jaberi L, Cai X, Bailey AM, et al. Chronic stress induces a selective decrease in AMPA receptor-mediated synaptic excitation at hippocampal temporoammonic-CA1 synapses. J Neurosci. 2013;33:15669–74.
Iñiguez SD, Aubry A, Riggs LM, Alipio JB, Zanca RM, Flores-Ramirez, et al. Social defeat stress induces depression-like behavior and alters spine morphology in the hippocampus of adolescent male C57BL/6 mice. Neurobiol Stress. 2016;5:54–64.
Hyer MM, Shaw GA, Goswamee P, Dyer SK, Burns CM, Soriano E, et al. Chronic adolescent stress causes sustained impairment of cognitive flexibility and hippocampal synaptic strength in female rats. Neurobiol Stress. 2021;14:100303.
Chourbaji S, Vogt MA, Fumagalli F, Sohr R, Frasca A, Brandwein C, et al. AMPA receptor subunit 1 (GluR-A) knockout mice model the glutamate hypothesis of depression. FASEB J. 2008;22:3129–34.
Schmidt MV, Trümbach D, Weber P, Wagner K, Scharf SH, Liebl C, et al. Individual stress vulnerability is predicted by short-term memory and AMPA receptor subunit ratio in the hippocampus. J Neurosci. 2010;30:16949–58.
Freed WJ, Dillon-Carter O, Kleinman JE. Properties of [3H]AMPA binding in postmortem human brain from psychotic subjects and controls: increases in caudate nucleus associated with suicide. Exp Neurol. 1993;121:48–56.
Sequeira A, Mamdani F, Ernst C, Vawter MP, Bunney WE, Lebel V, et al. Global brain gene expression analysis links glutamatergic and GABAergic alterations to suicide and major depression. PLoS One. 2009;4:e6585.
Sokolowski M, Wasserman J, Wasserman D. Gene-level associations in suicide attempter families show overrepresentation of synaptic genes and genes differentially expressed in brain development. Am J Med Genet B Neuropsychiatr Genet. 2018;177:774–84.
Kimbrel NA, Ashley-Koch AE, Qin XJ, Lindquist JH, Garrett ME, Dennis MF, et al. A genome-wide association study of suicide attempts in the million veterans program identifies evidence of pan-ancestry and ancestry-specific risk loci. Mol Psychiatry. 2022;27:2264–72; https://doi.org/10.1038/s41380-022-01472-3.
Kang HJ, Voleti B, Hajszan T, Rajkowska G, Stockmeier CA, Licznerski P, et al. Decreased expression of synapse-related genes and loss of synapses in major depressive disorder. Nat Med. 2012;18:1413–7.
Tomar A, Polygalov D, McHugh TJ. Differential impact of acute and chronic stress on CA1 spatial coding and gamma oscillations. Front Behav Neurosci. 2021;15:710725.
Arikan MK, Metin B, Tarhan N. EEG gamma synchronization is associated with response to paroxetine treatment. J Affect Disord. 2018;235:114–6.
Alaiyed S, McCann M, Mahajan G, Rajkowska G, Stockmeier CA, Kellar KJ, et al. Venlafaxine stimulates an MMP-9-dependent increase in excitatory/inhibitory balance in a stress model of depression. J Neurosci. 2020;40:4418–31.
Andreetta F, Carboni L, Grafton G, Jeggo R, Whyment AD, van den Top M, et al. Hippocampal 5-HT7 receptors signal phosphorylation of the GluA1 subunit to facilitate AMPA receptor mediated-neurotransmission in vitro and in vivo. Br J Pharm. 2016;173:1438–51.
Du J, Gray NA, Falke CA, Chen W, Yuan P, Szabo ST, et al. Modulation of synaptic plasticity by antimanic agents: the role of AMPA glutamate receptor subunit 1 synaptic expression. J Neurosci. 2004;24:6578–89.
Martinez-Turrillas R, Frechilla D, Del Rio J. Chronic antidepressant treatment increases the membrane expression of AMPA receptors in rat hippocampus. Neuropharmacol. 2002;43:1230–7.
Bessa JM, Ferreira D, Melo I, Marques F, Cerqueira JJ, Palha JA, et al. The mood-improving actions of antidepressants do not depend on neurogenesis but are associated with neuronal remodeling. Mol Psychiatry. 2009;14:764–73.
Yi ES, Oh S, Lee JK, Leem YH. Chronic stress-induced dendritic reorganization and abundance of synaptosomal PKA-dependent CP-AMPA receptor in the basolateral amygdala in a mouse model of depression. Biochem Biophys Res Commun. 2017;486:671–8.
Méndez P, Pazienti A, Szabo G, Bacci A. Direct alteration of a specific inhibitory circuit of the hippocampus by antidepressants. J Neurosci. 2012;32:16616–28.
Berman RM, Cappiello A, Anand A, Oren DA, Heninger GR, Charney DS, et al. Antidepressant effects of ketamine in depressed patients. Biol Psychiatry. 2000;47:351–4.
Zarate CA Jr, Singh JB, Carlson PJ, Brutsche NE, Ameli R, Luckenbaugh DA, et al. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch Gen Psychiatry. 2006;63:856–64.
DiazGranados N, Ibrahim LA, Brutsche NE, Ameli R, Henter ID, Luckenbaugh DA, et al. Rapid resolution of suicidal ideation after a single infusion of an N-methyl-D-aspartate antagonist in patients with treatment-resistant major depressive disorder. J Clin Psychiatry. 2010;71:1605–11.
Lapidus KA, Levitch CF, Perez AM, Brallier JW, Parides MK, Soleimani L, et al. A randomized controlled trial of intranasal ketamine in major depressive disorder. Biol Psychiatry 2014;76:970–6. 1014
Lally N, Nugent AC, Luckenbaugh DA, Ameli R, Roiser JP, Zarate CA. Anti-anhedonic effect of ketamine and its neural correlates in treatment-resistant bipolar depression. Transl Psychiatry. 2014;4:e469.
Newport DJ, Carpenter LL, McDonald WM, Potash JB, Tohen M, Nemeroff CB. Ketamine and other NMDA antagonists: early clinical trials and possible mechanisms in depression. Am J Psychiatry. 2015;172:950–66.
Wilkinson ST, Ballard ED, Bloch MH, Mathew SJ, Murrough JW, Feder A, et al. The effect of a single dose of intravenous ketamine on suicidal ideation: a systematic review and individual participant data meta-analysis. Am J Psychiatry. 2018;175:150–8.
Conley AA, Norwood AEQ, Hatvany TC, Griffith JD, Barber KE. Efficacy of ketamine for major depressive episodes at 2, 4, and 6-weeks post-treatment: A meta-analysis. Psychopharmacol (Berl). 2021;238:1737–52.
Moghaddam B, Adams B, Verma A, Daly D. Activation of glutamatergic neurotransmission by ketamine: a novel step in the pathway from NMDA receptor blockade to dopaminergic and cognitive disruptions associated with the prefrontal cortex. J Neurosci. 1997;17:2921–7.
Hong LE, Summerfelt A, Buchanan RW, O’Donnell P, Thaker GK, Weiler MA, et al. Gamma and delta neural oscillations and association with clinical symptoms under subanesthetic ketamine. Neuropsychopharmacology 2010;35:632–40.
Caixeta FV, Cornélio AM, Scheffer-Teixeira R, Ribeiro S, Tort AB. Ketamine alters oscillatory coupling in the hippocampus. Sci Rep. 2013;3:2348.
Nagy D, Stoiljkovic M, Menniti FS, Hajós M. Differential effects of an NR2B NAM and ketamine on synaptic potentiation and gamma synchrony: relevance to rapid-onset antidepressant efficacy. Neuropsychopharmacol. 2016;41:1486–94.
Farmer CA, Gilbert JR, Moaddel R, George J, Adeojo L, Lovett J, et al. Ketamine metabolites, clinical response, and gamma power in a randomized, placebo-controlled, crossover trial for treatment-resistant major depression. Neuropsychopharmacol. 2020;45:1398–404.
Homayoun H, Moghaddam B. NMDA receptor hypofunction produces opposite effects on prefrontal cortex interneurons and pyramidal neurons. J Neurosci. 2007;27:11496–500.
Widman AJ, McMahon LL. Disinhibition of CA1 pyramidal cells by low-dose ketamine and other antagonists with rapid antidepressant efficacy. Proc Natl Acad Sci. 2018;115:E3007–E3016.
Buzsáki G, Wang X-J. Mechanisms of gamma oscillations. Ann Rev Neurosci. 2012;35:203–25.
Moda-Sava RN, Murdock MH, Parekh PK, Fetcho RN, Huang BS, Huynh TN, et al. Sustained rescue of prefrontal circuit dysfunction by antidepressant-induced spine formation. Science. 2019;364:eaat8078.
Garcia LS, Comim CM, Valvassori SS, Réus GZ, Barbosa LM, Andreazza AC, et al. Acute administration of ketamine induces antidepressant-like effects in the forced swimming test and increases BDNF levels in the rat hippocampus. Prog Neuropsychopharmacol Biol Psychiatry. 2008;32:140–4.
Du J, Machado-Vieira R, Maeng S, Martinowich K, Manji HK, Zarate CA Jr. Enhancing AMPA to NMDA throughput as a convergent mechanism for antidepressant action. Drug Disco Today Ther Strateg. 2006;3:519–26.
Maeng S, Zarate CA Jr, Du J, Schloesser RJ, McCammon J, Chen G, et al. Cellular mechanisms underlying the antidepressant effects of ketamine: role of alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptors. Biol Psychiatry. 2008;63:349–52.
Sanacora G, Smith MA, Pathak S, Su HL, Boeijinga PH, McCarthy DJ, et al. Lanicemine: a low-trapping NMDA channel blocker produces sustained antidepressant efficacy with minimal psychotomimetic adverse effects. Mol Psychiatry. 2014;19:978–85.
Ahnaou A, Ver Donck L, Drinkenburg WH. Blockade of the metabotropic glutamate (mGluR2) modulates arousal through vigilance states transitions: evidence from sleep-wake EEG in rodents. Behav Brain Res. 2014;270:56–67.
Zanos P, Nelson ME, Highland JN, Krimmel SR, Georgiou P, Gould TD, et al. A negative allosteric modulator for α5 subunit-containing GABA receptors exerts a rapid and persistent antidepressant-like action without the side effects of the NMDA receptor antagonist ketamine in mice. eNeuro. 2017; 4:ENEURO.0285-16.2017.
Troppoli TA, Zanos P, Georgiou P, Gould TD, Rudolph U, Thompson SM. Negative allosteric modulation of gamma-aminobutyric acid A receptors at α5 subunit-containing benzodiazepine sites reverses stress-induced anhedonia and weakened synaptic function in mice. Biol Psychiatry. 2022;92:216–26.
Zanos P, Moaddel R, Morris PJ, Georgiou P, Fischell J, Elmer GI, et al. NMDAR inhibition-independent antidepressant actions of ketamine metabolites. Nature. 2016;533:481–6.
Kavalali ET, Monteggia LM. Targeting homeostatic synaptic plasticity for treatment of mood disorders. Neuron 2020;106:715–26.
Sutton MA, Taylor AM, Ito HT, Pham A, Schuman EM. Postsynaptic decoding of neural activity: eEF2 as a biochemical sensor coupling miniature synaptic transmission to local protein synthesis. Neuron. 2007;55:648–61.
Autry AE, Adachi M, Nosyreva E, Na ES, Los MF, Cheng PF, et al. NMDA receptor blockade at rest triggers rapid behavioural antidepressant responses. Nature. 2011;475:91–5.
Nosyreva E, Szabla K, Autry AE, Ryazanov AG, Monteggia LM, Kavalali ET. Acute suppression of spontaneous neurotransmission drives synaptic potentiation. J Neurosci. 2013;33:6990–7002.
Adaikkan C, Taha E, Barrera I, David O, Rosenblum K. Calcium/calmodulin-dependent protein kinase II and eukaryotic elongation factor 2 kinase pathways mediate the antidepressant action of ketamine. Biol Psychiatry. 2018;84:65–75.
Suzuki K, Kim JW, Nosyreva E, Kavalali ET, Monteggia LM. Convergence of distinct signaling pathways on synaptic scaling to trigger rapid antidepressant action. Cell Rep. 2021;37:109918.
Opal MD, Klenotich SC, Morais M, Bessa J, Winkle J, Doukas D, et al. Serotonin 2C receptor antagonists induce fast-onset antidepressant effects. Mol Psychiatry. 2014;19:1106–14.
Riggs LM, Aracava Y, Zanos P, Fischell J, Albuquerque EX, Pereira EFR, et al. (2R,6R)-hydroxynorketamine rapidly potentiates hippocampal glutamatergic transmission through a synapse-specific presynaptic mechanism. Neuropsychopharmacol. 2020;45:426–36.
Kadriu B, Musazzi L, Johnston JN, Kalynchuk LE, Caruncho HJ, Popoli M, et al. Positive AMPA receptor modulation in the treatment of neuropsychiatric disorders: A long and winding road. Drug Disco Today. 2021;26:2816–38.
Li X, Tizzano JP, Griffey K, Clay M, Lindstrom T, Skolnick P. Antidepressant-like actions of an AMPA receptor potentiator (LY392098). Neuropharmacol. 2001;40:1028–33.
Bai F, Bergeron M, Nelson DL. Chronic AMPA receptor potentiator (LY451646) treatment increases cell proliferation in adult rat hippocampus. Neuropharmacol. 2003;44:1013–21.
Farley S, Apazoglou K, Witkin JM, Giros B, Tzavara ET. Antidepressant-like effects of an AMPA receptor potentiator under a chronic mild stress paradigm. Int J Neuropsychopharmacol. 2010;13:1207–18.
Gordillo-Salas M, Pascual-Antón R, Ren J, Greer J, Adell A. Antidepressant-like effects of CX717, a positive allosteric modulator of AMPA receptors. Mol Neurobiol. 2020;57:3498–507.
Wallace T, Myers B. Effects of Biological Sex and Stress Exposure on Ventromedial Prefrontal Regulation of Mood-Related Behaviors. Front Behav Neurosci. 2021;15:737960.
Wilson RC, Takahashi YK, Schoenbaum G, Niv Y. Orbitofrontal cortex as a cognitive map of task space. Neuron. 2014;81:267–79.
Howard JD, Kahnt T. To be specific: The role of orbitofrontal cortex in signaling reward identity. Behav Neurosci. 2021;135:210–7.
Klein-Flügge MC, Bongioanni A, Rushworth MFS. Medial and orbital frontal cortex in decision-making and flexible behavior, Neuron 2022; https://doi.org/10.1016/j.neuron.2022.05.022
McClelland H, Evans JJ, Nowland R, Ferguson E, O’Connor RC. Loneliness as a predictor of suicidal ideation and behaviour: a systematic review and meta-analysis of prospective studies. J Affect Disord. 2020;274:880–96.
Pearce E, Myles-Hooton P, Johnson S, Hards E, Olsen S, Clisu D, et al. Loneliness as an active ingredient in preventing or alleviating youth anxiety and depression: a critical interpretative synthesis incorporating principles from rapid realist reviews. Transl Psychiatry. 2021;11:628.
Grueschow M, Polania R, Hare TA, Ruff CC. Automatic versus choice-dependent value representations in the human brain. Neuron. 2015;85:874–85.
Wang XJ. Decision making in recurrent. Neuronal Circuits Neuron. 2008;60:215–34.
Wong KF, Wang XJ. A recurrent network mechanism of time integration in perceptual decisions. J Neurosci 2006;26:1314–28.
Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF. Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci. 2005;8:365–71.
Berridge KC, Kringelbach ML. Pleasure systems in the brain. Neuron. 2015;86:646–64.
Lewis EJ, Blanco I, Raila H, Joormann J. Does repetitive negative thinking affect attention? Differential effects of worry and rumination on attention to emotional stimuli. Emotion. 2019;19:1450–62.
Ochsner K, Gross JJ. The cognitive control of emotion. Trends Cogn Sci. 2005;9:242–9.
Keedwell PA, Andrew C, Williams SC, Brammer MJ, Phillips ML. The neural correlates of anhedonia in major depressive disorder. Biol Psychiatry. 2005;58:843–53.
Cheng W, Rolls ET, Qiu J, Liu W, Tang Y, Huang CC, et al. Medial reward and lateral non-reward orbitofrontal cortex circuits change in opposite directions in depression. Brain. 2016;139:3296–309.
Green IW, Pizzagalli DA, Admon R, Kumar P. Anhedonia modulates the effects of positive mood induction on reward-related brain activation. Neuroimage. 2019;193:115–25.
Fales CL, Barch DM, Rundle MM, Mintun MA, Snyder AZ, Cohen JD, et al. Altered emotional interference processing in affective and cognitive-control brain circuitry in major depression. Biol Psychiatry. 2008;63:377–84.
Liston C, McEwen BS, Casey BJ. Psychosocial stress reversibly disrupts prefrontal processing and attentional control. Proc Natl Acad Sci. 2009;106:912–7.
Dias-Ferreira E, Sousa JC, Melo I, Morgado P, Mesquita AR, Cerqueira JJ, et al. Chronic stress causes frontostriatal reorganization and affects decision-making. Science. 2009;325:621–5.
Radley JJ, Sisti HM, Hao J, Rocher AB, McCall T, Hof PR, et al. Chronic behavioral stress induces apical dendritic reorganization in pyramidal neurons of the medial prefrontal cortex. Neuroscience. 2004;125:1–6.
Radley JJ, Rocher AB, Miller M, Janssen WG, Liston C, Hof PR, et al. Repeated stress induces dendritic spine loss in the rat medial prefrontal cortex. Cereb Cortex. 2006;16:313–20.
Liston C, Miller MM, Goldwater DS, Radley JJ, Rocher AB, Hof PR, et al. Stress-induced alterations in prefrontal cortical dendritic morphology predict selective impairments in perceptual attentional set-shifting. J Neurosci. 2006;26:7870–4.
Li N, Liu RJ, Dwyer JM, Banasr M, Lee B, Son H, et al. Glutamate N-methyl-D-aspartate receptor antagonists rapidly reverse behavioral and synaptic deficits caused by chronic stress exposure. Biol Psychiatry. 2011a;69:754–61.
Liston C, Gan WB. Glucocorticoids are critical regulators of dendritic spine development and plasticity in vivo. Proc Natl Acad Sci. 2011;108:16074–9.
Yuen EY, Wei J, Liu W, Zhong P, Li X, Yan Z. Repeated Stress Causes Cognitive Impairment by Suppressing Glutamate Receptor Expression and Function in Prefrontal Cortex. Neuron. 2012;73:962–77.
Kim U, Lee T. Topography of descending projections from anterior insular and medial prefrontal regions to the lateral habenula of the epithalamus in the rat. Eur J Neurosci. 2012;35:1253–69.
LeGates TA, Kvarta MD, Tooley JR, Francis TC, Lobo MK, Creed MC, et al. Reward behaviour is regulated by the strength of hippocampus-nucleus accumbens synapses. Nature. 2018;564:258–62.
Covington HE 3rd, Lobo MK, Maze I, Vialou V, Hyman JM, Zaman S, et al. Antidepressant effect of optogenetic stimulation of the medial prefrontal cortex. J Neurosci. 2010;30:16082–90.
Newton DF, Oh H, Shukla R, Misquitta K, Fee C, Banasr M, et al. Chronic stress induces coordinated cortical microcircuit cell-type transcriptomic changes consistent with altered information processing. Biol Psychiatry. 2022;91:798–809.
Soares JM, Sampaio A, Ferreira LM, Santos NC, Marques F, Palha JA, et al. Stress-induced changes in human decision-making are reversible. Transl Psychiatry. 2012;2:e131.
Fales CL, Barch DM, Rundle MM, Mintun MA, Mathews J, Snyder AZ, et al. Antidepressant treatment normalizes hypoactivity in dorsolateral prefrontal cortex during emotional interference processing in major depression. J Affect Disord. 2009;112:206–11.
Phillips ML, Chase HW, Sheline YI, Etkin A, Almeida JR, Deckersbach T, et al. Identifying predictors, moderators, and mediators of antidepressant response in major depressive disorder: neuroimaging approaches. Am J Psychiatry. 2015;172:124–38.
Fonseka TM, MacQueen GM, Kennedy SH. Neuroimaging biomarkers as predictors of treatment outcome in major depressive disorder. J Affect Disord. 2018;233:21–35.
Fitzgerald PB, Hoy K, McQueen S, Maller JJ, Herring S, Segrave R, et al. A randomized trial of rTMS targeted with MRI based neuro-navigation in treatment-resistant depression. Neuropsychopharmacol. 2009;34:1255–62.
Rudebeck PH, Rich EL, Mayberg HS. From bed to bench side: Reverse translation to optimize neuromodulation for mood disorders. Proc Natl Acad Sci. 2019;116:26288–96.
Li N, Lee B, Liu RJ, Banasr M, Dwyer JM, Iwata M, et al. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science. 2010;329:959–64.
Phoumthipphavong V, Barthas F, Hassett S, Kwan AC. Longitudinal effects of ketamine on dendritic architecture in vivo in the mouse medial frontal cortex. eNeuro. 2016; 3:ENEURO.0133-15.2016.
Holmes SE, Finnema SJ, Naganawa M, DellaGioia N, Holden D, Fowles K, et al. Imaging the effect of ketamine on synaptic density (SV2A) in the living brain. Mol Psychiatry. 2022;27:2273–81. https://doi.org/10.1038/s41380-022-01465-2
Rajkowska G, O’Dwyer G, Teleki Z, Stockmeier CA, Miguel-Hidalgo JJ. GABAergic neurons immunoreactive for calcium binding proteins are reduced in the prefrontal cortex in major depression. Neuropsychopharmacol. 2007;32:471–82.
Sibille E, Morris H, Kota R, Lewis DA. GABA-related transcripts in the dorsolateral prefrontal cortex in mood disorders. J Neuropsychopharmacol. 2011;14:721–34.
Tripp A, Oh H, Guilloux J-P, Martinowich K, Lewis DA, Sibille E. Brain-derived neurotrophic factor signaling and subgenual anterior cingulate cortex dysfunction in major depressive disorder. Am J Psychiatry. 2012;169:1194–202.
Girgenti MJ, Wohleb ES, Mehta S, Ghosal S, Fogaca MV, Duman RS. Prefrontal cortex interneurons display dynamic sex-specific stress-induced transcriptomes. Transl Psychiatry. 2019;9:292.
Banasr M, Lepack A, Fee C, Duric V, Maldonado-Aviles J, DiLeone R, et al. Characterization of GABAergic marker expression in the chronic unpredictable stress model of depression. Chronic Stress (Thousand Oaks). 2017;1:2470547017720459.
Gerhard DM, Pothula S, Liu RJ, Wu M, Li XY, Girgenti MJ, et al. GABA interneurons are the cellular trigger for ketamine’s rapid antidepressant actions. J Clin Invest. 2020;130:1336–49.
Ghosal S, Duman CH, Liu RJ, Wu M, Terwilliger R, Girgenti MJ, et al. Ketamine rapidly reverses stress-induced impairments in GABAergic transmission in the prefrontal cortex in male rodents. Neurobiol Dis. 2020;134:104669.
Fogaça MV, Wu M, Li C, Li XY, Picciotto MR, Duman RS. Inhibition of GABA interneurons in the mPFC is sufficient and necessary for rapid antidepressant responses. Mol Psychiatry. 2021;26:3277–91.
Liu RJ, Duman C, Kato T, Hare B, Lopresto D, Bang E, et al. GLYX-13 produces rapid antidepressant responses with key synaptic and behavioral effects distinct from ketamine. Neuropsychopharmacol. 2017;42:1231–42.
Ali F, Gerhard DM, Sweasy K, Pothula S, Pittenger C, Duman RS, et al. Ketamine disinhibits dendrites and enhances calcium signals in prefrontal dendritic spines. Nat Commun. 2020;11:72.
Volkow ND, Wise RA, Baler R. The dopamine motive system: implications for drug and food addiction. Nat Rev Neurosci. 2017;18:741–52.
Floresco SB. The nucleus accumbens: an interface between cognition, emotion, and action. Annu Rev Psychol. 2015;66:25–52.
Bayassi-Jakowicka M, Lietzau G, Czuba E, Steliga A, Waśkow M, Kowiański P. Neuroplasticity and multilevel system of connections determine the integrative role of nucleus accumbens in the brain reward system. Int J Mol Sci. 2021;22:9806.
Heimer L, Zahm DS, Churchill L, Kalivas PW, Wohltmann C. Specificity in the projection patterns of accumbal core and shell in the rat. Neurosci. 1991;41:89–125.
Brog JS, Salyapongse A, Deutch AY, Zahm DS. The patterns of afferent innervation of the core and shell in the “accumbens” part of the rat ventral striatum: immunohistochemical detection of retrogradely transported fluoro-gold. J Comp Neurol. 1993;338:255–78.
Watabe-Uchida M, Eshel N, Uchida N. Neural circuitry of reward prediction error. Annu Rev Neurosci. 2017;40:373–94.
Kupchik YM, Brown RM, Heinsbroek JA, Lobo MK, Schwartz DJ, Kalivas PW. Coding the direct/indirect pathways by D1 and D2 receptors is not valid for accumbens projections. Nat Neurosci. 2015;18:1230–2.
Soares-Cunha C, de Vasconcelos N, Coimbra B, Domingues AV, Silva JM, Loureiro-Campos E, et al. Nucleus accumbens medium spiny neurons subtypes signal both reward and aversion. Mol Psychiatry 2019;25:3241–55.218.
Kravitz AV, Tye LD, Kreitzer AC. Distinct roles for direct and indirect pathway striatal neurons in reinforcement. Nat Neurosci. 2012;15:816–8.
Tai LH, Lee AM, Benavidez N, Bonci A, Wilbrecht L. Transient stimulation of distinct subpopulations of striatal neurons mimics changes in action value. Nat Neurosci. 2012;15:1281–9.
Wu H, Kakusa B, Neuner S, Christoffel DJ, Heifets BD, Malenka RC, et al. Local accumbens in vivo imaging during deep brain stimulation reveals a strategy-dependent amelioration of hedonic feeding. Proc Natl Acad Sci. 2022;119:e2109269118.
Francis TC, Chandra R, Friend DM, Finkel E, Dayrit G, Miranda J, et al. Nucleus accumbens medium spiny neuron subtypes mediate depression-related outcomes to social defeat stress. Biol Psychiatry. 2015;77:212–22.
Britt JP, Benaliouad F, McDevitt RA, Stuber GD, Wise RA, Bonci A. Synaptic and behavioral profile of multiple glutamatergic inputs to the nucleus accumbens. Neuron. 2012;76:790–803.
Otis JM, Namboodiri VM, Matan AM, Voets ES, Mohorn EP, Kosyk O, et al. Prefrontal cortex output circuits guide reward seeking through divergent cue encoding. Nature. 2017;543:103–7.
Yagishita S, Hayashi-Takagi A, Ellis-Davies GC, Urakubo H, Ishii S, Kasai H. A critical time window for dopamine actions on the structural plasticity of dendritic spines. Science. 2014;345:1616–20.
Sjulson L, Peyrache A, Cumpelik A, Cassataro D, Buzsáki G. Cocaine place conditioning strengthens location-specific hippocampal coupling to the nucleus accumbens. Neuron. 2018;98:926–34.
Ma YY, Lee BR, Wang X, Guo C, Liu L, Cui R, et al. Bidirectional modulation of incubation of cocaine craving by silent synapse-based remodeling of prefrontal cortex to accumbens projections. Neuron. 2014;83:1453–67.
Creed M, Pascoli VJ, Lüscher C. Addiction therapy. Refining deep brain stimulation to emulate optogenetic treatment of synaptic pathology. Science. 2015;347:659–64.
Francis TC, Lobo MK. Emerging role for nucleus accumbens medium spiny neuron subtypes in depression. Biol Psychiatry. 2017;81:645–53.
Francis TC, Chandra R, Gaynor A, Konkalmatt P, Metzbower SR, Evans B, et al. Molecular basis of dendritic atrophy and activity in stress susceptibility. Mol Psychiatry. 2017;22:1512–19.
Fox ME, Chandra R, Menken MS, Larkin EJ, Nam H, Engeln M, et al. Dendritic remodeling of D1 neurons by RhoA/Rho-kinase mediates depression-like behavior. Mol Psychiatry. 2020;25:1022–34.
Lim BK, Huang KW, Grueter BA, Rothwell PE, Malenka RC. Anhedonia requires MC4R-mediated synaptic adaptations in nucleus accumbens. Nature 2012;487:183–9.
Christoffel DJ, Walsh JJ, Heifets BD, Hoerbelt P, Neuner S, Sun G, et al. Input-specific modulation of murine nucleus accumbens differentially regulates hedonic feeding. Nat Commun. 2021;12:2135.
Chakroborty S, Geisbush TR, Dale E, Pehrson AL, Sánchez C, West AR. Impact of vortioxetine on synaptic integration in prefrontal-subcortical circuits: comparisons with escitalopram. Front Pharm. 2017;8:764.
Minami S, Satoyoshi H, Ide S, Inoue T, Yoshioka M, Minami M. Suppression of reward-induced dopamine release in the nucleus accumbens in animal models of depression: Differential responses to drug treatment. Neurosci Lett. 2017;650:72–6.
Belujon P, Grace AA. Restoring mood balance in depression: ketamine reverses deficit in dopamine-dependent synaptic plasticity. Biol Psychiatry. 2014;76:927–36.
Witkin JM, Monn JA, Schoepp DD, Li X, Overshiner C, Mitchell SN, et al. The rapidly acting antidepressant ketamine and the mGlu2/3 receptor antagonist LY341495 rapidly engage dopaminergic mood circuits. J Pharm Exp Ther. 2016;358:71–82.
Hunt MJ, Olszewski M, Piasecka J, Whittington MA, Kasicki S. Effects of NMDA receptor antagonists and antipsychotics on high frequency oscillations recorded in the nucleus accumbens of freely moving mice. Psychopharmacol. 2015;232:4525–35.
Koo JW, Chaudhury D, Han MH, Nestler EJ. Role of Mesolimbic Brain-Derived Neurotrophic Factor in Depression. Biol Psychiatry. 2019;86:738–48.
Asim M, Hao B, Waris A, Liang YM, Wang XG. Ketamine attenuates the PTSD-like effect via regulation of glutamatergic signaling in the nucleus accumbens of mice. Mol Cell Neurosci. 2022;120:103723.
Pool ER, Munoz Tord D, Delplanque S, Stussi Y, Cereghetti D, Vuilleumier P, et al. Differential contributions of ventral striatum subregions to the motivational and hedonic components of the affective processing of reward. J Neurosci. 2022;42:2716–28.
Schwarz K, Moessnang C, Schweiger JI, Harneit A, Schneider M, Chen J, et al. Ventral striatal-hippocampus coupling during reward processing as a stratification biomarker for psychotic disorders. Biol Psychiatry. 2022;91:216–25.
Kumar P, Goer F, Murray L, Dillon DG, Beltzer ML, Cohen AL, et al. Impaired reward prediction error encoding and striatal-midbrain connectivity in depression. Neuropsychopharmacol. 2018;43:1581–8.
Reinen JM, Whitton AE, Pizzagalli DA, Slifstein M, Abi-Dargham A, McGrath PJ, et al. Differential reinforcement learning responses to positive and negative information in unmedicated individuals with depression. Eur Neuropsychopharmacol. 2021;53:89–100.
Heller AS, Johnstone T, Light SN, Peterson MJ, Kolden GG, Kalin NH, et al. Davidson RJ. Relationships between changes in sustained fronto-striatal connectivity and positive affect in major depression resulting from antidepressant treatment. Am J Psychiatry. 2013;170:197–206.
Fischer AS, Holt-Gosselin B, Fleming SL, Hack LM, Ball TM, Schatzberg AF, et al. Intrinsic reward circuit connectivity profiles underlying symptom and quality of life outcomes following antidepressant medication: a report from the iSPOT-D trial. Neuropsychopharmacol. 2021;46:809–19.
Sterpenich V, Vidal S, Hofmeister J, Michalopoulos G, Bancila V, Warrot D, et al. Increased reactivity of the mesolimbic reward system after ketamine injection in patients with treatment-resistant major depressive disorder. Anesthesiol. 2019;130:923–35.
Lecca S, Meye FJ, Mameli M. The lateral habenula in addiction and depression: an anatomical, synaptic and behavioral overview. Eur J Neurosci. 2014;39:1170–8.
Poller WC, Madai VI, Bernard R, Laube G, Veh RW. A glutamatergic projection from the lateral hypothalamus targets VTA-projecting neurons in the lateral habenula of the rat. Brain Res. 2013;1507:45–60.
Kim U, Lee T. Topography of descending projections from anterior insular and medial prefrontal regions to the lateral habenula of the epithalamus in the rat. Eur J Neurosci. 2012;35:1253–69.
Li B, Piriz J, Mirrione M, Chung C, Proulx CD, Schulz D, et al. Synaptic potentiation onto habenula neurons in the learned helplessness model of depression. Nature. 2011;470:535–9.
Ji H, Shepard PD. Lateral habenula stimulation inhibits rat midbrain dopamine neurons through a GABA(A) receptor-mediated mechanism. J Neurosci. 2007;27:6923–30.
Matsumoto M, Hikosaka O. Representation of negative motivational value in the primate lateral habenula. Nat Neurosci. 2009;12:77–84.
Lecca S, Meye FJ, Trusel M, Tchenio A, Harris J, Schwarz MK, et al. Aversive stimuli drive hypothalamus-to-habenula excitation to promote escape behavior. Elife. 2017;6:1–16.
Cerniauskas I, Winterer J, de Jong JW, Lukacsovich D, Yang H, Khan F, et al. Chronic Stress Induces Activity, Synaptic, and Transcriptional Remodeling of the Lateral Habenula Associated with Deficits in Motivated Behaviors. Neuron. 2019;104:899–915.
Zheng Z, Guo C, Li M, Yang L, Liu P, Zhang X, et al. Hypothalamus-habenula potentiation encodes chronic stress experience and drives depression onset. Neuron. 2022;110:1400–15.
Jhou TC. The rostromedial tegmental (RMTg) “brake” on dopamine and behavior: A decade of progress but also much unfinished work. Neuropharmacol. 2021;198:108763.
Proulx CD, Aronson S, Milivojevic D, Molina C, Loi A, Monk B, et al. A neural pathway controlling motivation to exert effort. Proc Natl Acad Sci 2018;115:5792–7.
Lecca S, Pelosi A, Tchenio A, Moutkine I, Lujan R, Hervé D, et al. Rescue of GABAB and GIRK function in the lateral habenula by protein phosphatase 2A inhibition ameliorates depression-like phenotypes in mice. Nat Med. 2016;22:254–61.
Yang Y, Cui Y, Sang K, Dong Y, Ni Z, Ma S, et al. Ketamine blocks bursting in the lateral habenula to rapidly relieve depression. Nature. 2018a;554:317–22.
Cui Y, Yang Y, Ni Z, Dong Y, Cai G, Foncelle A, et al. Astroglial Kir4.1 in the lateral habenula drives neuronal bursts in depression. Nature. 2018;554:323–7.
Yang LM, Hu B, Xia YH, Zhang BL, Zhao H. Lateral habenula lesions improve the behavioral response in depressed rats via increasing the serotonin level in dorsal raphe nucleus. Behav Brain Res. 2008;188:84–90.
Li K, Zhou T, Liao L, Yang Z, Wong C, Henn F, et al. βCaMKII in lateral habenula mediates core symptoms of depression. Science. 2013;341:1016–20.
Winter C, Vollmayr B, Djodari-Irani A, Klein J, Sartorius A. Pharmacological inhibition of the lateral habenula improves depressive-like behavior in an animal model of treatment resistant depression. Behav Brain Res. 2011;216:463–5.
Yang Y, Wang H, Hu J, Hu H. Lateral habenula in the pathophysiology of depression. Curr Opin Neurobiol. 2018b;48:90–6.
Mondoloni S, Mameli M, Congiu M. Reward and aversion encoding in the lateral habenula for innate and learned behaviours. Transl Psychiatry. 2022;12:3.
Hofmann SG, Sawyer AT, Fang A, Asnaani A. Emotion dysregulation model of mood and anxiety disorders. Depress Anxiety. 2012;29:409–16.
Harmer CJ, O’sullivan U, Favaron E, Massey-Chase R, Ayres R, Reinecke A, et al. Effect of acute antidepressant administration on negative affective bias in depressed patients. Am J Psychiatry. 2009;166:1178–84.
Fischell J, Van Dyke AM, Kvarta MD, LeGates TA, Thompson SM. Rapid Antidepressant Action and Restoration of Excitatory Synaptic Strength After Chronic Stress by Negative Modulators of Alpha5-Containing GABAA Receptors. Neuropsychopharmacol. 2015;40:2499–509.
Davis AK, Barrett FS, May DG, Cosimano MP, Sepeda ND, Johnson MW, et al. Effects of psilocybin-assisted therapy on major depressive disorder: a randomized clinical trial. JAMA Psychiatry. 2021;78:481–9.
Hesselgrave N, Troppoli TA, Wulff AB, Cole AB, Thompson SM. Harnessing psilocybin: antidepressant-like behavioral and synaptic actions of psilocybin are independent of 5-HT2R activation in mice. Proc Natl Acad Sci. 2021;118:e2022489118.
Shao LX, Liao C, Gregg I, Davoudian PA, Savalia NK, Delagarza K, et al. Psilocybin induces rapid and persistent growth of dendritic spines in frontal cortex in vivo. Neuron 2021;109:2535–.e4.
Barrett FS, Doss MK, Sepeda ND, Pekar JJ, Griffiths RR. Emotions and brain function are altered up to one month after a single high dose of psilocybin. Sci Rep. 2020;10:2214.
Ly C, Greb AC, Cameron LP, Wong JM, Barragan EV, Wilson PC, et al. Psychedelics Promote Structural and Functional Neural Plasticity. Cell Rep. 2018;23:3170–3182.
Acknowledgements
I thank all of the members of the Thompson lab and our collaborators whose ideas and hard work are reflected in this review: Drs. Aileen Bailey, Xiang Cai, Anthony Cole, Jonathan Fischell, Todd Gould, Natalie Hesselgrave, Angy Kallarackal, Mark Kvarta, Tara LeGates, Timothy Troppoli, Andreas Wulff, and Panos Zanos.
Funding
This work is supported by NIH grant R01 MH086828.
Author information
Authors and Affiliations
Contributions
The University of Maryland, Baltimore, has filed multiple patents for novel antidepressant therapeutic approaches on which the author is listed as an inventor. The author is solely responsible for this review.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Thompson, S.M. Plasticity of synapses and reward circuit function in the genesis and treatment of depression. Neuropsychopharmacol. 48, 90–103 (2023). https://doi.org/10.1038/s41386-022-01422-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41386-022-01422-1
This article is cited by
-
Modulators of GABAA receptor-mediated inhibition in the treatment of neuropsychiatric disorders: past, present, and future
Neuropsychopharmacology (2024)
