
Abstract
Cannabis use peaks in adolescence, and adolescents may be more vulnerable to the neural effects of cannabis and cannabis-related harms due to ongoing brain development during this period. In light of ongoing cannabis policy changes, increased availability, reduced perceptions of harm, heightened interest in medicinal applications of cannabis, and drastic increases in cannabis potency, it is essential to establish an understanding of cannabis effects on the developing adolescent brain. This systematic review aims to: (1) synthesize extant literature on functional and structural neural alterations associated with cannabis use during adolescence and emerging adulthood; (2) identify gaps in the literature that critically impede our ability to accurately assess the effect of cannabis on adolescent brain function and development; and (3) provide recommendations for future research to bridge these gaps and elucidate the mechanisms underlying cannabis-related harms in adolescence and emerging adulthood, with the long-term goal of facilitating the development of improved prevention, early intervention, and treatment approaches targeting adolescent cannabis users (CU). Based on a systematic search of Medline and PsycInfo and other non-systematic sources, we identified 90 studies including 9441 adolescents and emerging adults (n = 3924 CU, n = 5517 non-CU), which provide preliminary evidence for functional and structural alterations in frontoparietal, frontolimbic, frontostriatal, and cerebellar regions among adolescent cannabis users. Larger, more rigorous studies are essential to reconcile divergent results, assess potential moderators of cannabis effects on the developing brain, disentangle risk factors for use from consequences of exposure, and elucidate the extent to which cannabis effects are reversible with abstinence. Guidelines for conducting this work are provided.
Similar content being viewed by others
Introduction
Cannabis use is extremely common [1,2,3], particularly among youth [e.g., reported by 38.3% of US 12th graders [4]. While there is substantial evidence that use is associated with harmful outcomes for a subset of cannabis users (CU) [5,6,7,8,9,10,11], many use without negative consequences. Cannabis has recently been decriminalized and/or legalized in many US states [12], and a bill to decriminalize cannabis was recently passed in the House of Representatives. In addition, cannabinoids have been proposed to have therapeutic potential [13]. However, current understanding of the effects of cannabis on brain and behavior—including neural mechanisms underlying cannabis-related harms—remains limited, particularly effects during neural development.
Cannabis use typically begins during adolescence and peaks in adolescence/emerging adulthood [14, 15]. Critically, adolescents may be more vulnerable to neural cannabis effects and cannabis-related harms due to ongoing brain development during this period [5, 16]. However, few studies have directly compared effects of cannabis between adolescents and adults, and results are mixed [17, 18]. Recent reviews and meta-analyses support cognitive deficits among adolescent relative to adult CU, and also suggest that these deficits may be reversible with abstinence [17, 19, 20]. Adolescence is a unique period characterized by the most substantial neural change aside from the perinatal period [21]. In particular, large-scale changes in neural architecture are thought to support the development of higher-order cognitive and emotional processes necessary for adaptive functioning in adulthood [22,23,24,25]. Therefore, effects of cannabis on the brain during adolescence may have important implications for long-term development.
Altered structure and function of brain regions implicated in executive functioning, emotion, reward, and memory have been reported among CU relative to nonusers [19, 26, 27], which may represent potential mechanisms of cannabis-related harms. However, the extant literature on neural correlates of cannabis use has been inconsistent [26, 28]. Many factors likely contribute to mixed results, including methodological differences, small sample sizes, inconsistencies in cannabinoid composition and potency, developmental timing of cannabis use, and various moderators of cannabis effects on the brain, such as sex and different patterns of use [16, 29, 30]. To address these limitations, this review seeks to: (1) review and synthesize extant literature on structural and functional neural correlates of cannabis use during adolescence and emerging adulthood; (2) identify gaps in the literature that critically impede our ability to accurately assess the effects of cannabis on adolescent brain function and development; and (3) provide recommendations for future research to bridge these gaps and elucidate the mechanisms underlying cannabis-related harms in adolescence and emerging adulthood. Strong data on the neural correlates of adolescent cannabis use is essential to facilitate the development of improved prevention, early intervention, and treatment approaches targeting adolescent CU.
Materials and methods
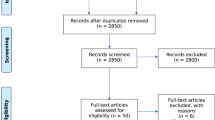
A systematic literature search of Medline and Psychinfo was conducted on June 10th, 2019 with 25 search terms encompassing the following parameters: adolescen* (OR young, youth, pubertal, puberty, minors, emerging adult, development) and cannabis (OR cannabidiol, cannabinoid, cbd, marijuana, thc) and mri (OR diffusion imaging, dti, fmri, fractional anisotropy, functional connectivity, magnetic resonance, microstructure, neuroimaging, resting state, white matter (WM)). This produced 510 studies (see Fig. 1 for PRISMA flowchart). Adolescence refers to the developmental epoch between childhood and adulthood, yet precise definitions vary [31]. While adolescence is traditionally considered to begin at puberty, the appropriate endpoint of adolescence remains debated. While many define adolescence as spanning from age 10–19 [32], it has been argued that an expanded definition of adolescence extending into the mid-20s is more consistent with neurodevelopmental trajectories, as well as the timing of major role transitions associated with adulthood [31]. In line with this expanded view, we define adolescence as extending until age 25, but have used the terms adolescence and emerging adulthood to accommodate the various definitions in the literature. Accordingly, inclusion criteria for the current review were as follows: mean age ≤ 25, minimum sample of 15 participants per group, and participants with personal cannabis use histories. THC administration studies in healthy volunteers and studies of prenatal cannabis exposure were excluded.

A systematic literature search of Medline and Psychinfo was conducted on June 10th, 2019, with 25 search terms, encompassing the following parameters: adolescen* and cannabis and mri. This produced 510 studies. 2 additional studies were identified during peer review. Studies were screened based on inclusion criteria: (1) participants with personal cannabis use histories (THC administration studies in healthy volunteers and studies of prenatal cannabis exposure were removed), (2) mean age ≤ 25, and (3) minimum cell size of 15 participants, and 90 studies were identified for inclusion in the current review. Note that studies that include multiple fMRI tasks or assess multiple structural characteristics are included in multiple sections, as appropriate.
The search strategy was developed by authors SDL, NM, and SWY. Author NM took the lead on screening manuscripts for inclusion, and all eligibility questions were resolved in consultation with SDL and SWY. SDL, NM, and SWY then divided the remaining 88 manuscripts into sections based on their primary methods and assigned sections to co-authors who conducted the literature review and drafted the accompanying table for their section. Two additional studies were identified during peer review, for a final total of 90 studies. To ensure that the information summarized in each table accurately reflects the published literature, each table was cross reviewed for accuracy by a second author, with points of discrepancy resolved by the first author (SDL; see Author Contributions).
Results
Sample characteristics
Ninety studies were selected for inclusion and further categorized into functional MRI studies using neurocognitive, inhibitory control, drug cue reactivity, reward, social/emotion, and resting-state paradigms, and structural studies of brain volumes, morphometry, and WM microstructure, as well as multimodal studies (see Fig. 1 for PRISMA flowchart). Together, these studies are comprised of data from 9441 adolescents and emerging adults (n = 3,924 CU, n = 5517 non-CU). Prevalence estimates for lifetime use are similar for male and female adolescents in the United States [33], yet 12 studies included solely male participants and no study included only females. Across all studies, only 35.9% of participants were women (see Fig. 2). Key findings and areas of convergence/divergence are summarized in the main text below and results of each study are detailed in accompanying tables.

Across all studies, only 35.9% of participants were female. Figure 2 displays the number of male and female participants included in each category reviewed.
Functional MRI literature
Neurocognitive functioning
As summarized in Table 1A, findings from cross-sectional studies generally indicate differences within corticolimbic and frontoparietal regions during memory task performance among youth with CU relative to non-users. However, the direction of these alterations has differed across studies and may be age-dependent, with increased activity reported among younger adolescents [34,35,36] and decreased activity reported among older adolescents and emerging adults [37, 38]. Whereas multiple studies using n-back tasks have not reported significant differences in neural activation during working memory performance between CU and non-users [39, 40], two studies found that working memory-related neural activation prospectively predicts initiation [41] or escalation [39] of use during spatial working memory [41] and n-back [39] tasks, respectively. In particular, increased frontoparietal [39, 41] and decreased visual and precuneus [41] engagement predicted cannabis use 6 months [39] and 3 years [41] later. Together, longitudinal data indicate limited effects of cannabis use on working memory function and raise the possibility that working-memory-related neural differences may precede cannabis use [39,40,41].
Several studies have also used dimensional (i.e., regression) or within-group (e.g., median split) analysis approaches to assess the effects of cannabis use characteristics (e.g., frequency, severity, age of initiation) on patterns of neural function during cognitive tasks. These data suggest that increased activation across several regions (i.e., frontoparietal, cingulate, insular, subcortical, and cerebellar) during neurocognitive processes may be linked to individual differences in cannabis use, including earlier age of initiation [42], and severity [43]/frequency of use [44]. Collectively, these data provide preliminary evidence for altered corticolimbic and frontoparietal activation among adolescent CU during neurocognitive task performance and suggest that these alterations may represent a risk factor for use rather than a consequence of exposure. Nonetheless, differences in cannabis inclusion criteria (e.g., lifetime vs. past-month use, as summarized in Table 1A) and task type (e.g., spatial working memory vs. n-back vs. associative learning tasks, as summarized in Table 1A) may contribute to variation in findings across studies.
Inhibitory control
Findings from studies of inhibitory control processing are summarized in Table 1B. Several studies converge in reporting significantly increased brain activation [45, 46] and/or functional connectivity [47] among CU relative to controls during successful inhibition, primarily within prefrontal, parietal, cingulate, and cerebellar areas. However, several other studies found no significant group differences [47,48,49]. Notably, increased neural activation among CU was coupled with comparable [45, 46, 48,49,50] or worse [47] behavioral task performance, suggesting possible compensatory patterns of neural response. Conversely, one study examining individuals at high familial risk for substance use found that lower right DLPFC activation was characteristic of those with personal cannabis/alcohol use during adolescence [50]. Given that this study combined adolescents with alcohol and cannabis use histories, it is difficult to reconcile these findings with the other studies reviewed. Overall, these data provide mixed evidence for altered frontoparietal, cingulate and cerebellar activation during inhibitory control performance among adolescent CU.
Drug cue reactivity
Findings from studies assessing neural response to cannabis cues are summarized in Table 1C. Among these, three compared cannabis to control cues [51,52,53] or tested a main effect of cannabis cues [53] in CU and reported greater cue reactivity to cannabis cues in brain regions involved in reward, incentive salience, and visual attention. Overlap (i.e., similar brain region reported in ≥2 studies) was found in the superior frontal gyrus, medial frontal gyrus, fusiform gyrus, inferior and middle occipital gyrus, inferior parietal lobule, precuneus, parahippocampal gyrus, hippocampus, amygdala, and thalamus. Additionally, acute cannabis administration reduced striatal cue reactivity for CU compared to non-substance users [54].
Two studies investigated whether cannabis cue-reactivity relates to cannabis dependence and/or addiction severity by comparing dependent and non-dependent CU and found no group differences in activation [55] or reward-related functional connectivity [56]. Nonetheless, cue exposure was associated with heightened functional connectivity between the nucleus accumbens (NAcc) and the anterior cingulate gyrus, caudate, and cerebellum across all CU [56], and cue-reactivity in the left putamen prospectively predicted problem severity at 3-year follow-up [55]. Another study of treatment-seeking adolescent CU [57] found a prospective association between increased striatal and cerebellar activation while participants were concurrently exposed to cannabis cues and their own change talk (statements in favor of reducing their cannabis use) and reduced frequency of use at a 1-month follow-up, suggesting clinical relevance. Two studies examined whether genetic variation may impact cannabis cue-reactivity and found modest effects of risk alleles for the genes that encode the cannabinoid receptor 1 and fatty acid amide hydrolase [58], as well as variants of the serotonin 2A receptor gene [59]. Finally, one study directly compared cue-reactivity between CU and matched controls [60] and found no group differences, though associations with lifetime and future use were noted among CU.
Overall, these studies provide some evidence for heightened frontostriatal, frontoparietal, and frontolimbic activation to cannabis cues among CU, but findings are inconclusive regarding how neural cannabis cue-reactivity relates to cannabis use characteristics. Importantly, the 10 available studies represent only six independent samples (same or overlapping samples: [52, 55,56,57,58,59,60]), and most do not meet current minimum statistical standards, so findings should be considered preliminary.
Reward
Findings from studies assessing non-drug reward responses are summarized in Table 1D. Research using a Monetary Incentive Delay (MID) task reported greater cannabis use was associated with higher concurrent anticipatory reward signals in the striatum [50] and predicted lower striatal anticipatory reward signals two years later [61]. By comparison, a separate study reported greater striatal activity relative to controls during anticipation of neutral monetary outcomes that was inversely related to age of cannabis use onset [62]. Together, findings from MID tasks suggest that striatal anticipatory processing in adolescent CU may be sensitive to use patterns and trajectories.
Findings from reward task studies have been less consistent for cortical brain regions, possibly due to differences in reward paradigms. For example, both increases in orbitofrontal cortex (OFC) activity during Iowa Gambling Task performance [63] and decreases in OFC activity during a Decision-Reward Uncertainty task have been reported among CU relative to controls [64]. A separate study of individuals with escalating use reported negative functional connectivity between striatal and prefrontal regions during a card-guessing game [65], raising the possibility of disrupted coordination of reward responses. Cortical alterations have also been linked to use patterns and trajectories, with greater use linked to higher cortical reward responses [63, 66], and lower OFC responses predicting relapse [64]. Overall, the current literature suggests that adolescents with CU display altered non-drug reward processing in frontostriatal regions.
Emotion regulation and social processing
Findings from studies utilizing emotion regulation and social processing tasks are summarized in Table 1E. Four studies have compared neural response to emotionally valanced stimuli between adolescent CU and non-users [43, 67,68,69], with most finding increased activation among CU in multiple brain regions. One study [67] found CU displayed greater activation in the right medial OFC and increased medial OFC coupling to contralateral dorsal striatum and amygdala to negative emotional stimuli during an emotion processing task. Another study using an affective Stroop task found a positive association between cannabis use disorder (CUD) severity and activation in precuneus, posterior cingulate, and inferior parietal lobule [43]. A third study using an affective face processing task [70] reported greater activation to angry faces in amygdala, middle temporal gyrus, and inferior frontal gyrus among individuals reporting light cannabis experimentation compared to controls [68]. In contrast, a fourth study found that CU displayed less activation to negative words on an emotion-arousal word task in temporal, prefrontal, and occipital cortices, insula, and amygdala [69]. While numerous between-study differences may account for these inconsistent findings, one important factor may be duration of abstinence prior to scanning: the study with the shortest abstinence (48 h) found a decreased neural response [69], whereas studies with more prolonged abstinence (28 days) found increased neural responses [43, 67].
Two studies using social influence tasks found increased neural responses within the caudate and NAcc in response to peer information among CU [71, 72]. There has also been one study in which male CU exhibited higher frontal, cingulate and amygdala activity, as well as reduced connectivity between amygdala and dorsolateral PFC, compared to controls during reappraisal of negative affect, suggesting diminished top-down control over negative affect [73]. Finally, another study examined functional connectivity of the OFC when treatment-seeking CU were exposed to their own change talk statements from a recent motivational interviewing session [74]. Compared to baseline, the change talk condition was associated with both increases and decreases in OFC connectivity with a variety of frontal, parietal, and subcortical regions, and greater connectivity with the anterior cingulate/medial frontal gyrus was associated with more post-treatment cannabis problems. Together, these studies support increased connectivity among CU during active regulation of behavior, providing an important foundation for future work. Overall, these data suggest that adolescent CU display altered frontolimbic activation and connectivity during emotional processing, with most consistent evidence found for the OFC.
Resting state
While task-based fMRI paradigms examine patterns of neural activation during specific cognitive tasks, resting-state fMRI (rsfMRI) aims to characterize intrinsic connectivity patterns independent from any distinct mode of cognitive processing [75]. Findings from these studies are summarized in Table 1F. Overall, resting-state connectivity of frontal [76,77,78,79,80,81,82,83], parietal [47, 78, 79, 82, 83], and cingulate [76,77,78,79] regions has been most consistently implicated among adolescent CU. However, the direction of these effects varies. Nonetheless, frontal, cingulate, and parietal regions are each implicated in multiple canonical resting-state networks [84], so it is unsurprising that the relationship between cannabis exposure and functional connectivity in these areas would be complex, particularly during adolescence when refinement of functional architecture is still ongoing [24, 85, 86]. Future research with larger, independent samples and more consistent methods is urgently needed to replicate and extend extant findings.
Several of the studies reviewed here examined differences in resting-state connectivity following a period of abstinence [78, 79, 83]. These reports converge in reporting that group differences are attenuated [78] or no longer present [79, 83] following one month of abstinence. These findings provide preliminary evidence that alterations in functional connectivity may be reversible following discontinuation of use. Nonetheless, two studies reported on the same sample of individuals [78, 79], and all focused on different regions and/or neural characteristics (functional connectivity [78, 79], cerebral blood flow [83]). Overall, the rsfMRI literature suggests that frontoparietal connectivity is altered among current CU, and that these effects may be attenuated with abstinence.
Structural MRI literature
Brain volume
Findings from cross-sectional studies comparing gray matter volume (GMV) between adolescent CU and non-users are summarized in Table 2A [87,88,89,90,91,92,93,94,95,96,97,98,99]. Aggregate findings indicate smaller thalamic [87] and larger amygdala [98, 99] volume among adolescent CU. Alterations in cerebellar [87,88,89, 94, 99], PFC [93, 99], and hippocampal [95, 99] volume have also been reported, though the direction has not been consistent across studies. In addition, several studies have found no significant volumetric differences between CU and non-CU groups [90,91,92, 96, 97]. These inconsistencies may relate to between-study differences in key demographic features proposed to influence the link between cannabis use and GMV during adolescence, such as sex [93, 98], age of first use [95, 100], frequency of use [100], synthetic versus plant-derived cannabis [87] and genetic variation [92]. It may be necessary to account for these and other key moderators in future work.
Several studies have also prospectively examined alterations in adolescent brain morphology in relation to subsequent cannabis use behaviors [88, 101, 102]. Smaller OFC volumes at age 12 have been associated with cannabis use onset by age 16 [102], and smaller volumes of the hippocampus/amygdala and superior temporal gyrus have been associated with increased quantity of cannabis use across a 3-year follow-up [88]. A separate prospective study reported an association between reduced bilateral amygdala volume and heightened cannabis craving following 28 days of abstinence in heavy CU [101]. These findings support the possibility that morphological variation may influence subsequent cannabis use, emphasizing the need to disentangle premorbid GMV alterations that impact risk for use from GMV changes that result from exposure.
Several studies have also assessed GMV in relation to CU in clinical populations. Findings from studies including adolescents and emerging adults at high-clinical risk for psychosis have been mixed, with decreased thalamus [103], amygdala/hippocampal complex [104] and amygdala volumes [105] reported among CU relative to non-users. In addition, the stability of these effects after co-varying for tobacco and other substance use has also varied [104, 105]. Increased putamen volume has been observed among males with schizophrenia and CUD, relative to those with schizophrenia alone [106], and individuals with schizophrenia and CUD had smaller insula volume relative to controls [106]. Conversely, a second study using a whole-brain approach found no differences between individuals with first episode psychosis with and without a history of cannabis use [107].
Overall, while there is some evidence for volumetric alterations in frontocerebellar [87,88,89, 93, 94, 99] and limbic subcortical regions [87, 95, 98, 99], many studies have failed to observe group differences [90,91,92, 96, 97], and a variety of variables have been identified as potential moderators of this relationship [87, 92, 93, 95, 98, 100]. Furthermore, several reports suggest that GMV alterations may precede cannabis use onset [88, 101, 102].
Brain morphometry
Findings from studies assessing cortical thickness (CT) among adolescent CU and non-users are shown in Table 2B [95, 108,109,110,111,112]. Among these, three studies examined populations without major comorbid psychopathology [108,109,110] and report largely divergent results. However, studies converge in reporting increased CT among CU in right superior and inferior parietal regions and the left paracentral lobule [108, 109], as well as negative correlations between age of onset and CT in the right superior frontal [109] and entorhinal cortices [108].
As with GMV, several studies have also assessed CT in CU with comorbid clinical diagnoses. Among males at high genetic risk for schizophrenia, higher levels of use were associated with reduced overall CT across three large population-based cohorts [112]. Similarly, among individuals with and without CUD and early-onset schizophrenia, those with CUD were characterized by an attenuated decrease in CT across 1.5 years in frontal, parietal, and temporal regions [111]. Finally, among individuals with attention-deficit/hyperactivity disorder, CU were characterized by reduced frontal and cingulate CT [95].
Four studies examined differences in gray matter density (GMD) [91, 97, 113, 114], with one study reporting increased GMD among CU in the NAcc and amygdala [97], and a second study finding no differences between groups [91]. Conversely, among samples with comorbid psychosis, lifetime history of CU was associated with decreased GMD in the right posterior cingulate cortex [113], and frontal, temporal, parietal, and occipital cortical areas, as well as the striatum and cerebellum [114]. Three studies also examined shape differences [91, 97, 114], with only one finding significant shape differences in the amygdala and NAcc [97]. Finally, one study examined local gyrification index and surface area of prefrontal and parietal ROIs and found less gyrification in bilateral medial PFC, frontal poles, and ventral medial PFC in the CU group, but no significant surface area findings [115].
Overall, these data provide initial evidence for alterations in frontoparietal CT [108, 109] and changes in NAcc and amygdala morphology [97] among adolescent CU. However, given the heterogeneity of morphological metrics and patient populations examined, it is difficult to draw any firm conclusions.
White matter
Findings from studies examining WM microstructure among adolescent CU using diffusion-weighted imaging (DWI) are shown in Table 2C. The most commonly reported metric across studies is fractional anisotropy (FA), a scalar index of diffusivity thought to reflect axon fiber density, axon diameter, and myelination (though interpretation of FA in areas of complex fiber architecture is more ambiguous) [116]. Among studies in adolescents without significant comorbid psychopathology [90, 117, 118], one found significantly reduced FA in the bilateral genu of the corpus callosum and the left internal capsule [117]. A second longitudinal study found FA decreased among CU over a two-year period in several tracts, including bilateral superior longitudinal fasciculi, right corticospinal tract, right anterior thalamic radiation, and superior fronto-occipital fasciculus [118]. Conversely, a third study did not find any association between adolescent CU and WM indices [90], but this discrepancy may be attributable to lower levels of use in the latter study. Consistent with this, earlier age of onset (prior to 16 years) has been related to lower genu FA [117], suggesting that patterns of use may be an important consideration.
Several studies have also examined WM microstructure among adolescent CU with significant psychopathology. Smaller WM volume [119] and poorer WM integrity of the anterior thalamic radiations and uncinate fasciculus [120] have been linked to heightened depressive symptoms among adolescent CU. WM alterations have also been examined among adolescent CU with comorbid psychotic disorders [107, 114, 121, 122]. Two of these studies found reduced FA of the inferior longitudinal fasciculus [121], superior longitudinal fasciculus, corpus callosum, internal capsule, and superior cerebellar decussation [114] among CU. A third study reported no differences between CU and non-users in emerging adults with a first episode of psychosis [107]. Conversely, a fourth study found increased FA in the uncinate fasciculus and anterior internal capsule in adolescents with recent-onset schizophrenia and comorbid cannabis and hard drug use [122]. However, the high degree of comorbidity between cannabis and other drug use makes it difficult to determine causality for the WM alterations observed [122].
The majority of the work reviewed here demonstrates that adolescent CU display disruptions in WM integrity, reported as lower FA in CU groups [114, 117, 118, 120, 121], although a subset of studies found no differences in WM microstructure [90, 107, 119], or increases in FA [122]. Negative associations between cannabis use and WM coherence appear to be widespread, affecting callosal [114, 117], association [114, 118, 120, 121], projection [114, 118], and brainstem [114] fibers. However, most studies are cross-sectional [114, 117, 119, 120, 122], and given the ambiguity of measures such as FA within many regions of the brain (i.e., those of complex architecture), additional work using other metrics and more sophisticated analysis approaches is warranted [116, 123,124,125].
Multimodal
One study used a machine learning technique to identify predictors of future cannabis initiation, including both structural and functional neural features [126] (Table 2D). A large sample of adolescents from the IMAGEN study were stratified into groups based on whether they reported any cannabis use by age 16, and a variety of psychosocial, brain, and genetic features at age 14 (when all participants were cannabis naive) were assessed as potential predictors of subsequent initiation. Models reliably predicted future cannabis use and identified both structural and function neural predictors, including frontal, cingulate, temporal, occipital, cerebellar, midbrain, and thalamic regions. Critically, distinct non-overlapping neural features were identified in male and female participants, further supporting the idea that sex may moderate associations between cannabis use and adolescent brain structure and function. Furthermore, the current study underscores the potential utility of predictive modeling approaches that look simultaneously at an array of neural characteristics to provide a deeper understanding of the complex relationships between cannabis use and adolescent neurodevelopment.
Discussion
Overall, extant literature on functional and structural neural alterations among adolescent and emerging adult CU most consistently implicates frontoparietal, frontolimbic, and frontostriatal regions, as well as the cerebellum. This is unsurprising, given that these regions are known to undergo substantial neurodevelopmental change during adolescence [127], and the psychoactive effects of cannabis are primarily mediated by CB1 receptors [128], which are most densely expressed in the neocortex, hippocampus, amygdala, basal ganglia, cerebellum, and brainstem [128, 129]. However, several studies have identified neural alterations that precede cannabis use onset and/or escalation [39,40,41, 88, 101, 102]. Therefore, it remains to be determined whether findings reflect premorbid risk factors or consequences of cannabis exposure. Furthermore, extant studies have also highlighted key factors that may moderate the effects of cannabis on the adolescent brain, including sex [93, 98, 112], genetic variation [58, 59, 92, 120], and differences in individual patterns of cannabis use, including age of onset [42, 77, 95, 100, 108, 109, 117], quantity/frequency of use [44, 100, 112], and duration of abstinence [43, 67, 69, 78, 79, 83]. Additionally, our review included studies with both younger and older adolescents/emerging adults (mean age ≤ 25) and thus includes data from individuals across different stages of puberty, which could also influence the effects of cannabis on the brain. Overall, the relationship between cannabis use and adolescent brain development is likely complex, and future studies should account for potential moderators in order to elucidate the potential impact of cannabis exposure.
Notably, the literature overall is characterized by severely insufficient sample sizes and methodological inconsistencies that limit the conclusions that can be drawn: 81.1% of studies reviewed here included cell sizes <50, and many report uncorrected results or inadequately correct for multiple comparisons. The past decade has seen a revolution in how neuroimaging studies are powered and in how multiple comparisons are corrected for, thus it is essential that future studies recruit larger samples of adolescent CU and apply rigorous statistical thresholds. For example, recent work suggests that cross-sectional studies of N~25 are unlikely to yield highly stable results [130]. To guide future research in this area, we have outlined several key limitations to the existing literature and provided recommendations for future studies, as summarized in Table 3.
Levels of use in both cannabis and comparison “non-cannabis” groups is strikingly inconsistent across studies and is often insufficiently described. Further, many studies describe their sample as “heavy” or “chronic” CU, yet these terms are used inconsistently. While these inconsistencies likely reflect the heterogeneity of CU, it is essential that future studies thoroughly characterize and report cannabis use characteristics for both study groups in a consistent manner to facilitate meaningful comparisons across studies.
Very few studies include measures of cannabis potency and cannabinoid composition. Cannabis potency has increased dramatically over the last several decades [131, 132], and evidence suggests that the effects of cannabis may vary based on the cannabinoid composition of products consumed [133] (i.e., THC:CBD ratio [134]). Therefore, inconsistencies in findings may be attributable to variation in cannabis potency or cannabinoid composition. Although these characteristics are difficult to measure [135], future research should seek to quantify and account for these factors wherever possible.
Length of abstinence prior to scanning may represent an important source of variance that should be considered and accurately reported. Several studies reviewed here suggest that neurobiological differences between current CU and non-users are substantially reduced, or even eliminated, following 1 month of abstinence [78, 79, 83]. These data highlight the need for future research to clearly quantify the duration of abstinence prior to scanning, and to investigate the circumstances under which the effects of cannabis may be reversible with abstinence.
Although several of the studies reviewed here suggest that age of onset [42, 77, 95, 100, 108, 109, 117] and the quantity and/or frequency of cannabis use [44, 100, 112] may moderate the effects of cannabis on the adolescent brain, few studies have examined how different patterns of cannabis use relate to structural and functional neural characteristics. Further, definitions of early- versus late-onset use have varied among studies that have examined this factor. It is important that future studies investigate how different patterns of use impact adolescent neurodevelopment in addition to reporting group differences.
While a handful of studies have incorporated longitudinal neuroimaging [61, 76, 78, 79, 83, 88, 103, 104, 111, 112, 118, 121], the vast majority of the literature remains cross-sectional. Future work incorporating longitudinal neuroimaging [e.g., [136] is essential to adequately disentangle neural mechanisms of risk for cannabis use from sequelae of cannabis exposure, as well as to assess the extent to which cannabis effects on the adolescent brain are reversible with abstinence. Furthermore, research collecting neuroimaging data with greater temporal resolution (i.e., densely sampled data [137, 138]) is essential to adequately characterize dynamic network development within the context of different patterns of adolescent cannabis use.
Finally, despite substantial preclinical evidence [139] and preliminary human findings [93, 98, 112, 140] indicative of sex differences in the effects of cannabis on the adolescent brain, the extent to which cannabis effects differ between female and male adolescents remains very poorly understood. Female adolescents have been underrepresented in the literature reviewed here, and only a small minority of studies have directly examined sex differences [93, 98, 112]. Although cannabis use is more prevalent among men than women at the global level [15], adequate representation of both female and male participants is essential to enable analyses investigating sex differences in cannabis effects.
Overall, the literature reviewed here provides preliminary evidence for functional and structural alterations in frontoparietal, frontolimbic, frontostriatal, and cerebellar regions among adolescent and emerging adult CU. In light of ongoing changes to cannabis policy [12], increased availability [141], reduced perceptions of harm [142], heightened interest in medicinal applications of cannabis [143], as well as drastic increases in cannabis potency [131, 132], it is essential that larger, more rigorous studies build upon this preliminary literature to elucidate the effects of cannabis on the developing adolescent brain. Furthermore, in order to establish how cannabis effects on the brain differ between adolescents/emerging adults and adults, additional research directly comparing these groups is urgently needed. While ongoing open-science, population-level neuroimaging initiatives, such as the Adolescent Brain and Cognitive Development (ABCD) study, will be critical in making adequately powered datasets widely available, there will still be a need for additional systematic work assessing relationships in clinical contexts and at finer temporal resolutions.
References
Substance Abuse and Mental Health Services Administration. Key substance use and mental health indicators in the United States: Results from the 2017 National Survey on Drug Use and Health (HHS Publication No. SMA 18-5068, NSDUH Series H-53). Rockville, MD: Center for Behavioral Health Statistics and Quality, Substance Abuse and Mental Health Services Administration. (2018). Retrieved from https://www.samhsa.gov/data/.
Degenhardt L, Hall W. Extent of illicit drug use and dependence, and their contribution to the global burden of disease. Lancet. 2012;379:55–70.
Organization WH. Substance abuse facts & figures: Cannabis. 2019. https://www.who.int/substance_abuse/facts/cannabis/en/. Accessed 18 July 2019.
Johnston LD, Miech RA, O’Malley PM, Bachman JG, Schulenberg JE, Patrick ME. Monitoring the Future national survey results on drug use 1975-2020: Overview, key findings on adolescent drug use. Ann Arbor: Institute for Social Research, University of Michigan; 2021.
Volkow ND, Baler RD, Compton WM, Weiss SR. Adverse health effects of marijuana use. N Engl J Med. 2014;370:2219–27.
Fergusson DM, Boden JM, Horwood LJ. Psychosocial sequelae of cannabis use and implications for policy: findings from the Christchurch Health and Development Study. Soc Psychiatry Psychiatr Epidemiol. 2015;50:1317–26.
Marconi A, Di Forti M, Lewis CM, Murray RM, Vassos E. Meta-analysis of the association between the level of Cannabis use and risk of psychosis. Schizophr Bull. 2016;42:1262–9.
Gobbi G, Atkin T, Zytynski T, Wang S, Askari S, Boruff J, et al. Association of cannabis use in adolescence and risk of depression, anxiety, and suicidality in young adulthood: a systematic review and meta-analysis. JAMA Psychiatry. 2019;76:426–34.
Lopez-Quintero C, Perez de los Cobos J, Hasin DS, Okuda M, Wang S, Grant BF, et al. Probability and predictors of transition from first use to dependence on nicotine, alcohol, cannabis, and cocaine: results of the National Epidemiologic Survey on Alcohol and Related Conditions (NESARC). Drug Alcohol Depend. 2011;115:120–30.
Blanco C, Hasin DS, Wall MM, Florez-Salamanca L, Hoertel N, Wang S, et al. Cannabis use and risk of psychiatric disorders: prospective evidence from a US National Longitudinal Study. JAMA Psychiatry. 2016;73:388–95.
Agrawal A, Neale MC, Prescott CA, Kendler KS. A twin study of early cannabis use and subsequent use and abuse/dependence of other illicit drugs. Psychol Med. 2004;34:1227–37.
Carliner H, Brown QL, Sarvet AL, Hasin DS. Cannabis use, attitudes, and legal status in the U.S.: a review. Prev Med. 2017;104:13–23.
Whiting PF, Wolff RF, Deshpande S, Di Nisio M, Duffy S, Hernandez AV, et al. Cannabinoids for medical use: a systematic review and meta-analysis. JAMA. 2015;313:2456–73.
Johnston LD, Miech RA, O’Malley PM, Bachman JG, Schulenberg JE, Patrick ME. Monitoring the Future national survey results on drug use 1975-2018: Overview, key findings on adolescent drug use. Ann Arbor: Institute for Social Research, University of Michigan; 2019.
World Drug Report 2021 (United Nations publication, Sales No. E.21.XI.8).
Blest-Hopley G, Colizzi M, Giampietro V, Bhattacharyya S. Is the adolescent brain at greater vulnerability to the effects of cannabis? A narrative review of the evidence. Front Psychiatry. 2020;11:859.
Gorey C, Kuhns L, Smaragdi E, Kroon E, Cousijn J. Age-related differences in the impact of cannabis use on the brain and cognition: a systematic review. Eur Arch Psychiatry Clin Neurosci. 2019;269:37–58.
Levine A, Clemenza K, Rynn M, Lieberman J. Evidence for the risks and consequences of adolescent cannabis exposure. J Am Acad Child Adolesc Psychiatry. 2017;56:214–25.
Lorenzetti V, Hoch E, Hall W. Adolescent cannabis use, cognition, brain health and educational outcomes: a review of the evidence. Eur Neuropsychopharmacol. 2020;36:169–80.
Scott JC, Slomiak ST, Jones JD, Rosen AFG, Moore TM, Gur RC. Association of cannabis with cognitive functioning in adolescents and young adults: a systematic review and meta-analysis. JAMA Psychiatry. 2018;75:585–95.
National Academies of Sciences E, Medicine. The promise of adolescence: realizing opportunity for all youth. Washington, DC: The National Academies Press; 2019.
Casey BJ, Heller AS, Gee DG, Cohen AO. Development of the emotional brain. Neurosci Lett. 2019;693:29–34.
Casey BJ, Galvan A, Somerville LH. Beyond simple models of adolescence to an integrated circuit-based account: a commentary. Dev Cogn Neurosci. 2016;17:128–30.
Power JD, Fair DA, Schlaggar BL, Petersen SE. The development of human functional brain networks. Neuron. 2010;67:735–48.
Kaufmann T, Alnaes D, Doan NT, Brandt CL, Andreassen OA, Westlye LT. Delayed stabilization and individualization in connectome development are related to psychiatric disorders. Nat Neurosci. 2017;20:513–15.
Bloomfield MAP, Hindocha C, Green SF, Wall MB, Lees R, Petrilli K, et al. The neuropsychopharmacology of cannabis: a review of human imaging studies. Pharmacol Ther. 2019;195:132–61.
Blest-Hopley G, Giampietro V, Bhattacharyya S. Residual effects of cannabis use in adolescent and adult brains - a meta-analysis of fMRI studies. Neurosci Biobehav Rev. 2018;88:26–41.
Chye Y, Kirkham R, Lorenzetti V, McTavish E, Solowij N, Yücel M. Cannabis, cannabinoids, and brain morphology: a review of the evidence. Biol Psychiatry Cogn Neurosci Neuroimaging. 2021;6:627–35.
Rossetti MG, Mackey S, Patalay P, Allen NB, Batalla A, Bellani M, et al. Sex and dependence related neuroanatomical differences in regular cannabis users: findings from the ENIGMA Addiction Working Group. Transl Psychiatry. 2021;11:272.
Lorenzetti V, Solowij N, Yucel M. The role of cannabinoids in neuroanatomic alterations in cannabis users. Biol Psychiatry. 2016;79:e17–31.
Sawyer SM, Azzopardi PS, Wickremarathne D, Patton GC. The age of adolescence. Lancet Child Adolesc Health. 2018;2:223–28.
Organization WH. Recognizing adolescence. https://apps.who.int/adolescent/second-decade/section2/page1/recognizing-adolescence.html. Accessed 14 July 2021.
Center for Behavioral Health Statistics and Quality. Results from the 2019 National Survey on Drug Use and Health: Detailed tables. Rockville, MD: Substance Abuse and Mental Health Services Administration. (2020). Retrieved from https://www.samhsa.gov/data/.
Padula CB, Schweinsburg AD, Tapert SF. Spatial working memory performance and fMRI activation interaction in abstinent adolescent marijuana users. Psychol Addict Behav. 2007;21:478–87.
Schweinsburg AD, Nagel BJ, Schweinsburg BC, Park A, Theilmann RJ, Tapert SF. Abstinent adolescent marijuana users show altered fMRI response during spatial working memory. Psychiatry Res. 2008;163:40–51.
Jager G, Block RI, Luijten M, Ramsey NF. Cannabis use and memory brain function in adolescent boys: a cross-sectional multicenter functional magnetic resonance imaging study. J Am Acad Child Adolesc Psychiatry 2010;49:561–72, 72.e1-3.
Carey SE, Nestor L, Jones J, Garavan H, Hester R. Impaired learning from errors in cannabis users: Dorsal anterior cingulate cortex and hippocampus hypoactivity. Drug Alcohol Depend. 2015;155:175–82.
Dager AD, Tice MR, Book GA, Tennen H, Raskin SA, Austad CS, et al. Relationship between fMRI response during a nonverbal memory task and marijuana use in college students. Drug Alcohol Depend. 2018;188:71–78.
Cousijn J, Wiers RW, Ridderinkhof KR, van den Brink W, Veltman DJ, Goudriaan AE. Effect of baseline cannabis use and working-memory network function on changes in cannabis use in heavy cannabis users: a prospective fMRI study. Hum Brain Mapp. 2014;35:2470–82.
Cousijn J, Vingerhoets WA, Koenders L, de Haan L, van den Brink W, Wiers RW, et al. Relationship between working-memory network function and substance use: a 3-year longitudinal fMRI study in heavy cannabis users and controls. Addict Biol. 2014;19:282–93.
Tervo-Clemmens B, Simmonds D, Calabro FJ, Montez DF, Lekht JA, Day NL, et al. Early cannabis use and neurocognitive risk: a prospective functional neuroimaging study. Biol Psychiatry Cogn Neurosci Neuroimaging. 2018;3:713–25.
Becker B, Wagner D, Gouzoulis-Mayfrank E, Spuentrup E, Daumann J. The impact of early-onset cannabis use on functional brain correlates of working memory. Prog Neuropsychopharmacol Biol Psychiatry. 2010;34:837–45.
Aloi J, Blair KS, Crum KI, Meffert H, White SF, Tyler PM, et al. Adolescents show differential dysfunctions related to Alcohol and Cannabis Use Disorder severity in emotion and executive attention neuro-circuitries. Neuroimage Clin. 2018;19:782–92.
Becker B, Wagner D, Gouzoulis-Mayfrank E, Spuentrup E, Daumann J. Altered parahippocampal functioning in cannabis users is related to the frequency of use. Psychopharmacology. 2010;209:361–74.
Gruber SA, Dahlgren MK, Sagar KA, Gonenc A, Killgore WD. Age of onset of marijuana use impacts inhibitory processing. Neurosci Lett. 2012;511:89–94.
Tapert SF, Schweinsburg AD, Drummond SP, Paulus MP, Brown SA, Yang TT, et al. Functional MRI of inhibitory processing in abstinent adolescent marijuana users. Psychopharmacology. 2007;194:173–83.
Behan B, Connolly CG, Datwani S, Doucet M, Ivanovic J, Morioka R, et al. Response inhibition and elevated parietal-cerebellar correlations in chronic adolescent cannabis users. Neuropharmacology 2014;84:131–7.
Claus ED, Feldstein Ewing SW, Magnan RE, Montanaro E, Hutchison KE, Bryan AD. Neural mechanisms of risky decision making in adolescents reporting frequent alcohol and/or marijuana use. Brain Imaging Behav. 2018;12:564–76.
Filbey F, Yezhuvath U. Functional connectivity in inhibitory control networks and severity of cannabis use disorder. Am J Drug Alcohol Abus 2013;39:382–91.
Martz ME, Zucker RA, Schulenberg JE, Heitzeg MM. Psychosocial and neural indicators of resilience among youth with a family history of substance use disorder. Drug Alcohol Depend. 2018;185:198–206.
Karoly HC, Schacht JP, Meredith LR, Jacobus J, Tapert SF, Gray KM, et al. Investigating a novel fMRI cannabis cue reactivity task in youth. Addict Behav. 2019;89:20–28.
Filbey FM, Schacht JP, Myers US, Chavez RS, Hutchison KE. Marijuana craving in the brain. Proc Natl Acad Sci USA. 2009;106:13016–21.
Charboneau EJ, Dietrich MS, Park S, Cao A, Watkins TJ, Blackford JU, et al. Cannabis cue-induced brain activation correlates with drug craving in limbic and visual salience regions: preliminary results. Psychiatry Res. 2013;214:122–31.
de Sousa Fernandes Perna EB, Theunissen EL, Kuypers KP, Evers EA, Stiers P, Toennes SW, et al. Brain reactivity to alcohol and cannabis marketing during sobriety and intoxication. Addict Biol. 2017;22:823–32.
Vingerhoets WA, Koenders L, van den Brink W, Wiers RW, Goudriaan AE, van Amelsvoort T, et al. Cue-induced striatal activity in frequent cannabis users independently predicts cannabis problem severity three years later. J Psychopharmacol. 2016;30:152–8.
Filbey FM, Dunlop J. Differential reward network functional connectivity in cannabis dependent and non-dependent users. Drug Alcohol Depend. 2014;140:101–11.
Feldstein Ewing SW, McEachern AD, Yezhuvath U, Bryan AD, Hutchison KE, Filbey FM. Integrating brain and behavior: evaluating adolescents’ response to a cannabis intervention. Psychol Addict Behav. 2013;27:510–25.
Filbey FM, Schacht JP, Myers US, Chavez RS, Hutchison KE. Individual and additive effects of the CNR1 and FAAH genes on brain response to marijuana cues. Neuropsychopharmacology. 2010;35:967–75.
Feldstein Ewing SW, Mead HK, Yezhuvath U, Dewitt S, Hutchison KE, Filbey FM. A preliminary examination of how serotonergic polymorphisms influence brain response following an adolescent cannabis intervention. Psychiatry Res. 2012;204:112–6.
Cousijn J, Goudriaan AE, Ridderinkhof KR, van den Brink W, Veltman DJ, Wiers RW. Approach-bias predicts development of cannabis problem severity in heavy cannabis users: results from a prospective FMRI study. PLoS ONE. 2012;7:e42394.
Martz ME, Trucco EM, Cope LM, Hardee JE, Jester JM, Zucker RA, et al. Association of marijuana use with blunted nucleus accumbens response to reward anticipation. JAMA Psychiatry. 2016;73:838–44.
Jager G, Block RI, Luijten M, Ramsey NF. Tentative evidence for striatal hyperactivity in adolescent cannabis-using boys: a cross-sectional multicenter fMRI study. J Psychoact Drugs. 2013;45:156–67.
Cousijn J, Wiers RW, Ridderinkhof KR, van den Brink W, Veltman DJ, Porrino LJ, et al. Individual differences in decision making and reward processing predict changes in cannabis use: a prospective functional magnetic resonance imaging study. Addict Biol. 2013;18:1013–23.
De Bellis MD, Wang L, Bergman SR, Yaxley RH, Hooper SR, Huettel SA. Neural mechanisms of risky decision-making and reward response in adolescent onset cannabis use disorder. Drug Alcohol Depend. 2013;133:134–45.
Lichenstein SD, Musselman S, Shaw DS, Sitnick S, Forbes EE. Nucleus accumbens functional connectivity at age 20 is associated with trajectory of adolescent cannabis use and predicts psychosocial functioning in young adulthood. Addiction. 2017;112:1961–70.
Ford KA, Wammes M, Neufeld RW, Mitchell D, Theberge J, Williamson P, et al. Unique functional abnormalities in youth with combined marijuana use and depression: an FMRI study. Front Psychiatry. 2014;5:130.
Zimmermann K, Yao S, Heinz M, Zhou F, Dau W, Banger M, et al. Altered orbitofrontal activity and dorsal striatal connectivity during emotion processing in dependent marijuana users after 28 days of abstinence. Psychopharmacology. 2018;235:849–59.
Spechler PA, Orr CA, Chaarani B, Kan KJ, Mackey S, Morton A, et al. Cannabis use in early adolescence: Evidence of amygdala hypersensitivity to signals of threat. Dev Cogn Neurosci. 2015;16:63–70.
Heitzeg MM, Cope LM, Martz ME, Hardee JE, Zucker RA. Brain activation to negative stimuli mediates a relationship between adolescent marijuana use and later emotional functioning. Dev Cogn Neurosci. 2015;16:71–83.
Grosbras M-H, Paus T. Brain networks involved in viewing angry hands or faces. Cereb Cortex. 2006;16:1087–96.
Gilman JM, Schuster RM, Curran MT, Calderon V, van der Kouwe A, Evins AE. Neural mechanisms of sensitivity to peer information in young adult cannabis users. Cogn Affect Behav Neurosci. 2016;16:646–61.
Gilman JM, Lee S, Kuster JK, Lee MJ, Kim BW, van der Kouwe A, et al. Variable activation in striatal subregions across components of a social influence task in young adult cannabis users. Brain Behav. 2016;6:e00459.
Zimmermann K, Walz C, Derckx RT, Kendrick KM, Weber B, Dore B, et al. Emotion regulation deficits in regular marijuana users. Hum Brain Mapp. 2017;38:4270–79.
Feldstein Ewing SW, Chung T, Caouette JD, Ketcherside A, Hudson KA, Filbey FM. Orbitofrontal cortex connectivity as a mechanism of adolescent behavior change. Neuroimage. 2017;151:14–23.
Power JD, Cohen AL, Nelson SM, Wig GS, Barnes KA, Church JA, et al. Functional network organization of the human brain. Neuron. 2011;72:665–78.
Camchong J, Lim KO, Kumra S. Adverse effects of cannabis on adolescent brain development: a longitudinal study. Cereb Cortex. 2017;27:1922–30.
Lopez-Larson MP, Rogowska J, Yurgelun-Todd D. Aberrant orbitofrontal connectivity in marijuana smoking adolescents. Dev Cogn Neurosci. 2015;16:54–62.
Pujol J, Blanco-Hinojo L, Batalla A, Lopez-Sola M, Harrison BJ, Soriano-Mas C, et al. Functional connectivity alterations in brain networks relevant to self-awareness in chronic cannabis users. J Psychiatr Res. 2014;51:68–78.
Blanco-Hinojo L, Pujol J, Harrison BJ, Macia D, Batalla A, Nogue S, et al. Attenuated frontal and sensory inputs to the basal ganglia in cannabis users. Addict Biol. 2017;22:1036–47.
Houck JM, Bryan AD, Feldstein, Ewing SW. Functional connectivity and cannabis use in high-risk adolescents. Am J Drug Alcohol Abus. 2013;39:414–23.
Osuch EA, Manning K, Hegele RA, Theberge J, Neufeld R, Mitchell D, et al. Depression, marijuana use and early-onset marijuana use conferred unique effects on neural connectivity and cognition. Acta Psychiatr Scand. 2016;134:399–409.
Orr C, Morioka R, Behan B, Datwani S, Doucet M, Ivanovic J, et al. Altered resting-state connectivity in adolescent cannabis users. Am J Drug Alcohol Abus. 2013;39:372–81.
Jacobus J, Goldenberg D, Wierenga CE, Tolentino NJ, Liu TT, Tapert SF. Altered cerebral blood flow and neurocognitive correlates in adolescent cannabis users. Psychopharmacology. 2012;222:675–84.
Finn ES, Shen X, Scheinost D, Rosenberg MD, Huang J, Chun MM, et al. Functional connectome fingerprinting: identifying individuals using patterns of brain connectivity. Nat Neurosci. 2015;18:1664–71.
Vogel AC, Power JD, Petersen SE, Schlaggar BL. Development of the brain’s functional network architecture. Neuropsychol Rev. 2010;20:362–75.
Kelly AM, Di Martino A, Uddin LQ, Shehzad Z, Gee DG, Reiss PT, et al. Development of anterior cingulate functional connectivity from late childhood to early adulthood. Cereb Cortex. 2009;19:640–57.
Nurmedov S, Metin B, Ekmen S, Noyan O, Yilmaz O, Darcin A, et al. Thalamic and cerebellar gray matter volume reduction in synthetic cannabinoids users. Eur Addict Res. 2015;21:315–20.
Koenders L, Cousijn J, Vingerhoets WA, van den Brink W, Wiers RW, Meijer CJ, et al. Grey Matter changes associated with heavy cannabis use: a longitudinal sMRI study. PLoS ONE. 2016;11:e0152482.
Cousijn J, Wiers RW, Ridderinkhof KR, van den Brink W, Veltman DJ, Goudriaan AE. Grey matter alterations associated with cannabis use: results of a VBM study in heavy cannabis users and healthy controls. Neuroimage. 2012;59:3845–51.
Thayer RE, YorkWilliams S, Karoly HC, Sabbineni A, Ewing SF, Bryan AD, et al. Structural neuroimaging correlates of alcohol and cannabis use in adolescents and adults. Addiction. 2017;112:2144–54.
Weiland BJ, Thayer RE, Depue BE, Sabbineni A, Bryan AD, Hutchison KE. Daily marijuana use is not associated with brain morphometric measures in adolescents or adults. J Neurosci. 2015;35:1505–12.
Batalla A, Soriano-Mas C, Lopez-Sola M, Torrens M, Crippa JA, Bhattacharyya S, et al. Modulation of brain structure by catechol-O-methyltransferase Val(158) Met polymorphism in chronic cannabis users. Addict Biol. 2014;19:722–32.
Medina KL, McQueeny T, Nagel BJ, Hanson KL, Yang TT, Tapert SF. Prefrontal cortex morphometry in abstinent adolescent marijuana users: subtle gender effects. Addict Biol. 2009;14:457–68.
Medina KL, Nagel BJ, Tapert SF. Abnormal cerebellar morphometry in abstinent adolescent marijuana users. Psychiatry Res. 2010;182:152–9.
Lisdahl KM, Tamm L, Epstein JN, Jernigan T, Molina BS, Hinshaw SP, et al. The impact of ADHD persistence, recent cannabis use, and age of regular cannabis use onset on subcortical volume and cortical thickness in young adults. Drug Alcohol Depend. 2016;161:135–46.
Koenders L, Lorenzetti V, de Haan L, Suo C, Vingerhoets W, van den Brink W, et al. Longitudinal study of hippocampal volumes in heavy cannabis users. J Psychopharmacol. 2017;31:1027–34.
Gilman JM, Kuster JK, Lee S, Lee MJ, Kim BW, Makris N, et al. Cannabis use is quantitatively associated with nucleus accumbens and amygdala abnormalities in young adult recreational users. J Neurosci. 2014;34:5529–38.
McQueeny T, Padula CB, Price J, Medina KL, Logan P, Tapert SF. Gender effects on amygdala morphometry in adolescent marijuana users. Behav Brain Res. 2011;224:128–34.
Orr C, Spechler P, Cao Z, Albaugh M, Chaarani B, Mackey S, et al. Grey matter volume differences associated with extremely low levels of cannabis use in adolescence. J Neurosci. 2019;39:1817–27.
Battistella G, Fornari E, Annoni JM, Chtioui H, Dao K, Fabritius M, et al. Long-term effects of cannabis on brain structure. Neuropsychopharmacology. 2014;39:2041–8.
Padula CB, McQueeny T, Lisdahl KM, Price JS, Tapert SF. Craving is associated with amygdala volumes in adolescent marijuana users during abstinence. Am J Drug Alcohol Abus. 2015;41:127–32.
Cheetham A, Allen NB, Whittle S, Simmons JG, Yucel M, Lubman DI. Orbitofrontal volumes in early adolescence predict initiation of cannabis use: a 4-year longitudinal and prospective study. Biol Psychiatry. 2012;71:684–92.
Welch KA, Stanfield AC, McIntosh AM, Whalley HC, Job DE, Moorhead TW, et al. Impact of cannabis use on thalamic volume in people at familial high risk of schizophrenia. Br J Psychiatry. 2011;199:386–90.
Welch KA, Moorhead TW, McIntosh AM, Owens DG, Johnstone EC, Lawrie SM. Tensor-based morphometry of cannabis use on brain structure in individuals at elevated genetic risk of schizophrenia. Psychol Med. 2013;43:2087–96.
Buchy L, Mathalon DH, Cannon TD, Cadenhead KS, Cornblatt BA, McGlashan TH, et al. Relation between cannabis use and subcortical volumes in people at clinical high risk of psychosis. Psychiatry Res Neuroimaging. 2016;254:3–9.
Koenders L, Machielsen MW, van der Meer FJ, van Gasselt AC, Meijer CJ, van den Brink W, et al. Brain volume in male patients with recent onset schizophrenia with and without cannabis use disorders. J Psychiatry Neurosci. 2015;40:197–206.
Haller S, Curtis L, Badan M, Bessero S, Albom M, Chantraine F, et al. Combined grey matter VBM and white matter TBSS analysis in young first episode psychosis patients with and without cannabis consumption. Brain Topogr. 2013;26:641–7.
Jacobus J, Squeglia LM, Meruelo AD, Castro N, Brumback T, Giedd JN, et al. Cortical thickness in adolescent marijuana and alcohol users: a three-year prospective study from adolescence to young adulthood. Dev Cogn Neurosci. 2015;16:101–09.
Lopez-Larson MP, Bogorodzki P, Rogowska J, McGlade E, King JB, Terry J, et al. Altered prefrontal and insular cortical thickness in adolescent marijuana users. Behav Brain Res. 2011;220:164–72.
Mashhoon Y, Sava S, Sneider JT, Nickerson LD, Silveri MM. Cortical thinness and volume differences associated with marijuana abuse in emerging adults. Drug Alcohol Depend. 2015;155:275–83.
Epstein KA, Kumra S. Altered cortical maturation in adolescent cannabis users with and without schizophrenia. Schizophr Res. 2015;162:143–52.
French L, Gray C, Leonard G, Perron M, Pike GB, Richer L, et al. Early cannabis use, polygenic risk score for schizophrenia and brain maturation in adolescence. JAMA Psychiatry. 2015;72:1002–11.
Bangalore SS, Prasad KM, Montrose DM, Goradia DD, Diwadkar VA, Keshavan MS. Cannabis use and brain structural alterations in first episode schizophrenia-a region of interest, voxel based morphometric study. Schizophr Res. 2008;99:1–6.
James A, Hough M, James S, Winmill L, Burge L, Nijhawan S, et al. Greater white and grey matter changes associated with early cannabis use in adolescent-onset schizophrenia (AOS). Schizophr Res. 2011;128:91–7.
Shollenbarger SG, Price J, Wieser J, Lisdahl K. Impact of cannabis use on prefrontal and parietal cortex gyrification and surface area in adolescents and emerging adults. Dev Cogn Neurosci. 2015;16:46–53.
Jbabdi S, Behrens TE, Smith SM. Crossing fibres in tract-based spatial statistics. Neuroimage. 2010;49:249–56.
Gruber SA, Dahlgren MK, Sagar KA, Gonenc A, Lukas SE. Worth the wait: effects of age of onset of marijuana use on white matter and impulsivity. Psychopharmacology. 2014;231:1455–65.
Becker MP, Collins PF, Lim KO, Muetzel RL, Luciana M. Longitudinal changes in white matter microstructure after heavy cannabis use. Dev Cogn Neurosci. 2015;16:23–35.
Medina KL, Nagel BJ, Park A, McQueeny T, Tapert SF. Depressive symptoms in adolescents: associations with white matter volume and marijuana use. J Child Psychol Psychiatry. 2007;48:592–600.
Shollenbarger SG, Price J, Wieser J, Lisdahl K. Poorer frontolimbic white matter integrity is associated with chronic cannabis use, FAAH genotype, and increased depressive and apathy symptoms in adolescents and young adults. Neuroimage Clin. 2015;8:117–25.
Epstein KA, Kumra S. White matter fractional anisotropy over two time points in early onset schizophrenia and adolescent cannabis use disorder: a naturalistic diffusion tensor imaging study. Psychiatry Res. 2015;232:34–41.
Peters BD, de Haan L, Vlieger EJ, Majoie CB, den Heeten GJ, Linszen DH. Recent-onset schizophrenia and adolescent cannabis use: MRI evidence for structural hyperconnectivity? Psychopharmacol Bull. 2009;42:75–88.
Jeurissen B, Leemans A, Tournier JD, Jones DK, Sijbers J. Investigating the prevalence of complex fiber configurations in white matter tissue with diffusion magnetic resonance imaging. Hum Brain Mapp. 2013;34:2747–66.
Reveley C, Seth AK, Pierpaoli C, Silva AC, Yu D, Saunders RC, et al. Superficial white matter fiber systems impede detection of long-range cortical connections in diffusion MR tractography. Proc Natl Acad Sci USA. 2015;112:E2820–8.
Jones DK, Knosche TR, Turner R. White matter integrity, fiber count, and other fallacies: the do’s and don’ts of diffusion MRI. Neuroimage. 2013;73:239–54.
Spechler PA, Allgaier N, Chaarani B, Whelan R, Watts R, Orr C, et al. The initiation of cannabis use in adolescence is predicted by sex-specific psychosocial and neurobiological features. Eur J Neurosci. 2019;50:2346–56.
Rubia K. Functional brain imaging across development. Eur Child Adolesc Psychiatry. 2013;22:719–31.
Mackie K. Distribution of cannabinoid receptors in the central and peripheral nervous system. Handb Exp Pharmacol. 2005;168:299–325.
Kendall DA, Yudowski GA. Cannabinoid receptors in the central nervous system: their signaling and roles in disease. Front Cell Neurosci. 2016;10:294.
Marek S, Tervo-Clemmens B, Calabro FJ, Montez DF, Kay BP, Hatoum AS, et al. Towards reproducible brain-wide association studies. bioRxiv. 2020:2020.08.21.257758.
ElSohly MA, Mehmedic Z, Foster S, Gon C, Chandra S, Church JC. Changes in cannabis potency over the last 2 decades (1995-2014): analysis of current data in the United States. Biol Psychiatry. 2016;79:613–9.
Stuyt E. The problem with the current high potency THC marijuana from the perspective of an addiction psychiatrist. Mo Med. 2018;115:482–86.
Solymosi K, Kofalvi A. Cannabis: a treasure trove or Pandora’s box? Mini Rev Med Chem. 2017;17:1223–91.
Freeman AM, Petrilli K, Lees R, Hindocha C, Mokrysz C, Curran HV, et al. How does cannabidiol (CBD) influence the acute effects of delta-9-tetrahydrocannabinol (THC) in humans? A systematic review. Neurosci Biobehav Rev. 2019;107:696–712.
Jikomes N, Zoorob M. The cannabinoid content of legal cannabis in Washington State varies systematically across testing facilities and popular consumer products. Sci Rep. 2018;8:4519.
Albaugh MD, Ottino-Gonzalez J, Sidwell A, Lepage C, Juliano A, Owens MM, et al. Association of cannabis use during adolescence with neurodevelopment. JAMA Psychiatry. 2021;78:1031–40. https://doi.org/10.1001/jamapsychiatry.2021.1258.
Poldrack RA. Precision neuroscience: dense sampling of individual brains. Neuron. 2017;95:727–29.
Gordon EM, Laumann TO, Gilmore AW, Newbold DJ, Greene DJ, Berg JJ, et al. Precision functional mapping of individual human brains. Neuron. 2017;95:791–807 e7.
Cooper ZD, Craft RM. Sex-dependent effects of cannabis and cannabinoids: a translational perspective. Neuropsychopharmacology. 2018;43:34–51.
Matheson J, Sproule B, Di Ciano P, Fares A, Le Foll B, Mann RE, et al. Sex differences in the acute effects of smoked cannabis: evidence from a human laboratory study of young adults. Psychopharmacology. 2020;237:305–16.
Yu B, Chen X, Chen X, Yan H. Marijuana legalization and historical trends in marijuana use among US residents aged 12-25: results from the 1979-2016 National Survey on drug use and health. BMC Public Health. 2020;20:156.
Sarvet AL, Wall MM, Keyes KM, Cerda M, Schulenberg JE, O’Malley PM, et al. Recent rapid decrease in adolescents’ perception that marijuana is harmful, but no concurrent increase in use. Drug Alcohol Depend. 2018;186:68–74.
Ebbert JO, Scharf EL, Hurt RT. Medical cannabis. Mayo Clin Proc. 2018;93:1842–47.
Sagar KA, Dahlgren MK, Gonenc A, Racine MT, Dreman MW, Gruber SA. The impact of initiation: early onset marijuana smokers demonstrate altered Stroop performance and brain activation. Dev Cogn Neurosci. 2015;16:84–92.
Subramaniam P, Rogowska J, DiMuzio J, Lopez-Larson M, McGlade E, Yurgelun-Todd D. Orbitofrontal connectivity is associated with depression and anxiety in marijuana-using adolescents. J Affect Disord 2018;239:234–41.
Buchy L, Cannon TD, Anticevic A, Lyngberg K, Cadenhead KS, Cornblatt BA, et al. Evaluating the impact of cannabis use on thalamic connectivity in youth at clinical high risk of psychosis. BMC Psychiatry. 2015;15:276.
Thijssen S, Rashid B, Gopal S, Nyalakanti P, Calhoun VD, Kiehl KA. Regular cannabis and alcohol use is associated with resting-state time course power spectra in incarcerated adolescents. Drug Alcohol Depend. 2017;178:492–500.
Woo CW, Krishnan A, Wager TD. Cluster-extent based thresholding in fMRI analyses: pitfalls and recommendations. Neuroimage. 2014;91:412–9.
Eklund A, Nichols TE, Knutsson H. Cluster failure: Why fMRI inferences for spatial extent have inflated false-positive rates. Proc Natl Acad Sci USA. 2016;113:7900–5.
Office of Research Translation and Communications, NIAAA, NIH, DHHS. NIAAA Council Approves Definition of Binge Drinking. NIAAA Newsletter, Winter 2004, Issue 3, NIH Publication No. 04–5346. (2004). Retrieved from https://pubs.niaaa.nih.gov/publications/Newsletter/winter2004/Newsletter_Number3.pdf.
CDC. Adult tobacco use glossary. 2017. https://www.cdc.gov/nchs/nhis/tobacco/tobacco_glossary.htm.
Acknowledgements
This work was supported by NIDA T32DA022975 (PIs: Mason & Sinha) and NIDA K08DA051667 to SDL; NIDA K01DA044270 and NCATS UL1TR002240 (PIs: Mashour & Lumeng) to LMC; NIDA 1R01DA046334-01A1 to KAG; NIAAA K01AA024804 to JH; NIAAA K01AA024788, NIDA R03DA045788, and Women’s Health Research at Yale to ATH; and K01DA039299 to SWY. Drs. Lichenstein, Cope, Garrison, Hardee, Hillmer, Worhunsky, Yip, and Nick Manco, Leslie Egbo, Kristen Reeder, and Elisa Stern report no biomedical financial interests or potential conflicts of interest.
Author information
Authors and Affiliations
Contributions
SDL, NM, and SWY conceived of the topic for the current review and conducted the systematic literature search, which yielded 510 manuscripts for screening (see PRISMA flowchart). NM took the lead on screening manuscripts for inclusion, and all eligibility questions were resolved in consultation with SDL and SWY. Once screening was complete, SDL, NM, and SWY divided the remaining 88 manuscripts into sections based on the primary methods used, and assigned sections to additional co-authors. SDL drafted the introduction, discussion, and SDL the resting-state fMRI section of the text, as well as the corresponding table (Table 1F). SDL also conducted the accuracy check for Table 1B. NM drafted the inhibitory control section and corresponding table (Table 1B), as well as conducting the accuracy check for Table 1D. KAG drafted the drug cue-reactivity section and corresponding table (Table 1C), as well as conducting the accuracy check for Table 1A. LMC drafted the morphometry section and corresponding table (Table 2B) and conducted the accuracy check for Table 1F. JH drafted the white matter section and corresponding table (Table 2C) and conducted the accuracy check for Table 1E. PW and KR drafted the reward section and corresponding table (Table 1D), as well as conducting the accuracy check for Table 2C. ATH drafted the emotion regulation and processing section and corresponding table (Table 1E), as well as conducting the accuracy check for Table 2A. LE and SWY drafted the neurocognitive section and corresponding table (Table 1A) and conducted the accuracy check for Table 2B. EFS drafted the gray matter volumes section and corresponding table (Table 2A) and conducted the accuracy check for Table 1C. SWY edited and revised working drafts of the manuscript. SDL, NM, KAG, LMC, JH, PW, KR, LE, EFS, and SWY all reviewed the and approved the final version to be published and agree to be accountable for all aspects of the work.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Lichenstein, S.D., Manco, N., Cope, L.M. et al. Systematic review of structural and functional neuroimaging studies of cannabis use in adolescence and emerging adulthood: evidence from 90 studies and 9441 participants. Neuropsychopharmacol. 47, 1000–1028 (2022). https://doi.org/10.1038/s41386-021-01226-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41386-021-01226-9
This article is cited by
-
Cannabis Use and Cognitive Functioning Across the Lifespan
Current Addiction Reports (2024)
-
Wirksamkeit eines Cannabispräventionsprogramms in der Schule: Ergebnisse einer randomisiert-prospektiven Studie
Bundesgesundheitsblatt - Gesundheitsforschung - Gesundheitsschutz (2024)
-
Distinct neural networks predict cocaine versus cannabis treatment outcomes
Molecular Psychiatry (2023)
-
Consequences of adolescent drug use
Translational Psychiatry (2023)
-
Altered functional connectivity and oscillatory dynamics in polysubstance and cannabis only users during visuospatial processing
Psychopharmacology (2023)
