
Abstract
The ultimate solution to the global HIV-1 epidemic will probably require the development of a safe and effective vaccine. Multiple vaccine platforms have been evaluated in preclinical and clinical trials, but given the disappointing results of clinical efficacy studies so far, novel vaccine approaches are needed. In this Opinion article, we discuss the scientific basis and clinical potential of novel adenovirus and cytomegalovirus vaccine vectors for HIV-1 as two contrasting but potentially complementary vector approaches. Both of these vector platforms have demonstrated partial protection against stringent simian immunodeficiency virus challenges in rhesus monkeys using different immunological mechanisms.
Similar content being viewed by others


Main
Despite major advances in reducing HIV-1 mortality worldwide — mostly as a result of increased global access to antiretroviral therapy — the ultimate control of the global HIV-1 epidemic will almost certainly require the development of a safe and effective HIV-1 vaccine, given the limitations of antiretroviral therapy and other HIV-1 prevention approaches1,2. The goal of an HIV-1 vaccine is to block acquisition of HIV-1 infection, or alternatively, to lead to clearance of a transient infection. Various HIV-1 vaccine strategies have been evaluated in preclinical and clinical trials, but only four concepts have advanced to clinical efficacy testing so far3,4,5,6,7,8, as shown in Table 1. Additional promising and novel vaccine concepts must therefore be evaluated in humans to accelerate HIV-1 vaccine development.
Central to many current HIV-1 vaccine strategies are vaccine vectors, which are typically attenuated or harmless viruses or bacteria that transport HIV-1 antigens into host cells. Recent data have suggested that vectors are not simply inert vehicles for the passive delivery of antigens but rather are active inducers of innate immunity that generate the cytokine milieu in which adaptive immune responses are elicited. Moreover, vector biology seems to differ substantially even among similar vectors within a particular class.
In this Opinion article, we describe recent data regarding the clinical development of novel serotype adenovirus (Ad) and cytomegalovirus (CMV) vaccine vectors for HIV-1 vaccines. Novel Ad and CMV vectors have emerged during the past several years as promising vectors for HIV-1 and other pathogens, owing to their immunogenicity and protective efficacy in stringent non-human primate challenge studies. These stringent challenge models typically involve infection of rhesus monkeys with the neutralization-resistant simian immunodeficiency viruses SIVmac239 or SIVmac251. Moreover, vaccine immunogenicity and viral challenge studies using novel Ad and CMV vectors have shown the unique characteristics and advantages of each of these vector systems, which are discussed in detail below. Novel serotype Ad vectors are currently in early phases of clinical testing, and CMV vectors are being prepared for clinical studies. There are also other promising HIV-1 vaccine platforms, including poxvirus vectors, alphavirus vectors, herpesvirus vectors, DNA vaccines and purified proteins. These topics are beyond the scope of this Opinion article, in which we focus on recent non-human primate studies using novel Ad and CMV vectors.
Novel serotype Ad vectors
Ads are double-stranded DNA viruses that have a characteristic genomic and physical structure. Replication-incompetent Ad vectors are typically stable and immunogenic and can be produced in large quantities, which makes them attractive as vaccine platforms. Ad vectors have long functioned as key model systems for molecular biology and gene therapy. They have also been explored as candidate vaccines in recent years. Interestingly, the potent immunogenicity of Ad vectors has proven to be a major limitation for gene therapy applications by reducing the duration of transgene expression, but this property has been exploited by the vaccine field and has led to the development of Ad vaccine vectors. Ad vectors from multiple species and serotypes are currently being explored as candidate vaccines for a broad range of infectious pathogens as well as for cancers. Most Ad vector development programmes for HIV-1 have used non-replicating Ad vectors in which the early region 1 (E1) gene, which is essential for virus replication, is deleted; however, replicating Ad vectors are also currently being explored.
Experience with Ad5 vector-based HIV-1 vaccines. In the field of HIV-1 vaccine research, the use of non-replicating Ad serotype 5 (Ad5) vectors from Ad subgroup C was first evaluated in detail by Merck Research Laboratories9. In particular, Ad5 vectors that expressed SIV Gag led to effective virological control following challenge of rhesus monkeys with SHIV-89.6P10 but failed to block acquisition of infection or to provide sustained virological control of the more stringent, neutralization-resistant virus SIVmac239 (Refs 11, 12). As shown in Table 1, two Phase IIb clinical efficacy studies — Step (also known as HVTN 502) and Phambili (also known as HVTN 503) — were conducted in America and South Africa, respectively. These studies evaluated the Merck Ad5–Gag–Pol–Nef vaccine that aimed to elicit virus-specific cellular immune responses against internal viral proteins. Vaccinations in the Step study were discontinued in 2007, following an interim analysis that showed a lack of efficacy3. Moreover, there were more HIV-1 infections in vaccinees than in placebo recipients, particularly in the subgroup of men who were baseline Ad5 seropositive and uncircumcized3. The mechanism for this possible increase in the risk of HIV-1 acquisition remains unclear, but it has been hypothesized that activation of vector-specific CD4+ T cells at mucosal surfaces following Ad5 vaccination may have resulted in an increase in the number of target cells for HIV-1 infection13,14,15,16, although this hypothesis has not been confirmed experimentally. Vaccinations in the Phambili study were also terminated5, and unblinded follow-up of these individuals also suggested that there was an increase in HIV-1 infections in vaccinees.
The US National Institutes of Health (NIH) Vaccine Research Center developed another HIV-1 vaccine candidate that involved priming with a DNA vaccine that encoded modified HIV-1 Env immunogens as well as Gag–Pol–Nef and boosting with an Ad5 vector expressing these antigens. This Ad5 vector differed from the Merck Ad5 vector in several aspects, including the E4 region. This vaccine candidate was shown to provide partial protection against acquisition of the low-stringency challenge virus SIVsmE660 but failed to protect against the higher stringency challenge virus SIVmac251, which is more difficult to neutralize17. A Phase IIb efficacy study, HVTN 505, was then carried out, which demonstrated that this vaccine had no efficacy against HIV-1 infection in humans6. These findings indicate that the SIVsmE660 challenge model is insufficiently stringent to predict clinical efficacy in humans, which is consistent with earlier predictions18, and emphasize the importance of using stringent challenge models in the preclinical evaluation of HIV-1 vaccine candidates. The failure of Ad5 vectors in two large HIV-1 vaccine development programmes has also led to a detailed re-evaluation of other ongoing Ad-based HIV-1 vaccine efforts.
Development of alternative serotype Ad vectors. As potential alternatives to Ad5 vectors, a series of vectors from other human Ad serotypes, as well as Ad serotypes from other species, have been constructed, in part to circumvent the high titres of baseline Ad5-specific neutralizing antibodies in the developing world19,20. For example, Ad26 vectors from Ad subgroup D and Ad35 vectors from Ad subgroup B have recently been developed and tested in Phase I clinical trials21,22,23,24,25,26. Similarly, various promising chimpanzee Ad vectors have recently been produced and tested in Phase I clinical studies27,28,29,30.
As a specific case study, non-replicating Ad26 vectors are currently under consideration for advanced HIV-1 vaccine clinical development, and replicating Ad26 vectors are planned for early phase clinical evaluation. In light of the disappointing results with Ad5 vector-based vaccines so far, the rationale to proceed with Ad26 vectors is based on data showing that Ad26 is substantially biologically different from Ad5. Ad26-based vaccines have superior protective efficacy compared with Ad5-based vaccines against stringent SIVmac251 challenges in rhesus monkeys, and Ad26 does not seem to increase the number or activation status of total or vector-specific CD4+ T cells at mucosal surfaces in humans following vaccination. Moreover, next-generation Ad vectors can be engineered to express different and potentially improved HIV-1 antigens from those used in previous Ad5 programmes and are also being explored in the context of more potent heterologous prime–boost vector regimens.
Biological differences between Ad5 and Ad26. Several studies have shown that Ad5 and Ad26 differ markedly from both virological and immunological perspectives, as summarized in Table 2. Ad5 seroprevalence is nearly universal in humans with high neutralizing antibody titres throughout the developing world, whereas the seroprevalence of Ad26 is moderate, with substantially lower neutralizing antibody titres19,20. In terms of primary cellular receptors, it is well established that Ad5 uses the coxsackievirus and adenovirus receptor (CAR), but recent data show that Ad26, like Ad35, uses CD46 instead of CAR as its receptor21,31. Moreover, Ad5 primarily exhibits liver tropism in vivo, as a result of coagulation factor binding to the hypervariable regions of the Ad5 hexon major capsid protein, whereas liver tropism is not exhibited by Ad26 (Ref. 32). In addition, immune complexes that contain Ad5 are substantially more stimulatory to human dendritic cells than immune complexes that contain Ad26 (Ref. 33).
Ad5 and Ad26 also stimulate profoundly different innate cytokine responses, both in mice and in rhesus monkeys34,35, as well as distinct innate immune profiles following vaccination in humans, as shown by differential gene expression profiles on day 1, day 3 and day 7 after immunization (D.H.B., unpublished observations). Certain groups of genes are upregulated or downregulated by Ad5 but not by Ad26, whereas others are upregulated or downregulated by Ad26 but not by Ad5. These data suggest that Ad5 and Ad26 trigger phenotypically distinct innate inflammatory profiles.
Ad5 and Ad26 vectors also have different adaptive immune phenotypes. In mice, Ad5 vectors elicit high-magnitude but partially dysfunctional and exhausted T cell responses, which are characterized by low levels of the cytokine receptor CD127, the homing receptor CD62L and the effector cytokine interferon-γ (IFNγ), as well as high levels of the inhibitory receptor PD-1, whereas Ad26 vectors elicit more polyfunctional T cells, which are characterized by high levels of CD127, CD62L and IFNγ and low levels of PD-1 (Refs 36, 37). Similar differences have been observed in rhesus monkeys38. Moreover, Ad26-induced T cells were shown to expand more robustly than Ad5-induced T cells following re-exposure to a boosting antigen, and they afforded higher protective efficacy in mouse viral challenge models36. Taken together, these data show that Ad5 and Ad26 differ substantially in terms of key aspects of seroepidemiology, basic virology, innate immune profiles and adaptive immune phenotypes.
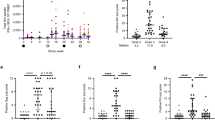
Protective efficacy of Ad26. Consistent with the negative results of the DNA–Ad5 efficacy trial in humans6, DNA–Ad5 vaccines similarly conferred no protective efficacy against repetitive, intrarectal challenges in the stringent SIVmac251 challenge model in rhesus monkeys17. Ad5-only vaccines also did not provide protection in this model11. By contrast, as shown in Fig. 1, heterologous prime–boost regimens that involve Ad26 vectors, together with either modified vaccinia virus Ankara (MVA) or Ad35 vectors, demonstrated partial protective efficacy against repetitive, intrarectal challenges with SIVmac251 in rhesus monkeys. Although most vaccinated animals became infected at the end of the challenge protocol, the risk of infection was reduced by 76–83% per exposure39. These data show that Ad26-based vaccine regimens provided partial protection in the stringent SIV challenge model in which Ad5 and DNA–Ad5 vaccines have failed.

Rhesus monkeys were immunized with Ad26–modified vaccinia virus Ankara (MVA) or Ad26–Ad35 vaccine regimens expressing Env–Gag–Pol antigens or with sham control vaccines and challenged repetitively with heterologous intrarectal inoculations with either SIVmac251 (N = 48; left-hand panel) or SHIV–SF162P3 (N = 36; right-hand panel). The number of challenges required to achieve infection is shown. Red lines indicate means. Data from Refs 39, 43.
We have also explored the use of bioinformatically optimized HIV-1 mosaic antigens that aim at providing improved immunological coverage of global virus diversity40,41,42, and we have shown that Ad26–MVA vaccines that express mosaic antigens provide partial protection against acquisition in repetitive, intrarectal SHIV–SF162P3 challenges43. In this study, correlates of protective efficacy included binding antibodies, functional non-neutralizing antibodies (including antibodies that mediate phagocytosis and complement deposition) and neutralizing antibodies, which suggests that the coordinated activity of multiple antibody functions may mediate protection against difficult-to-neutralize viruses43. We are currently exploring whether boosting with trimeric Env proteins with adjuvant can augment Ad26-primed antibody responses and thereby improve protection in both the SIVmac251 and SHIV–SF162P3 challenge models.
Ad26 and activation of mucosal CD4+ T cells. The mechanism for the possible increase in the risk of HIV-1 acquisition in individuals in the Step study remains unclear, but one hypothesis involves the potential activation of total or vector-specific CD4+ T cells at mucosal surfaces following Ad5 vaccination, which could theoretically result in an increase in the number of target cells for HIV-1 infection13,14,15,16. This hypothesis has never been directly evaluated in humans with Ad5 vectors. Nevertheless, to assess the extent of mucosal CD4+ T cell activation with Ad26 vectors, a randomized, double-blinded, placebo-controlled clinical trial (known as IPCAVD 003) was carried out to determine whether vaccination of healthy human subjects with an Ad26 vector that expressed HIV-1 Env would result in increased numbers or altered activation status of total or vector-specific CD4+ T cells in the colorectal mucosa44. Histopathological evaluation of colorectal biopsies showed no increases in CD4+ T cells, CD8+ T cells, CD3+ T cells, CD25+ T cells, or in cells that express the activation markers Ki67 or human leukocyte antigen-DR (HLA-DR) following Ad26 vaccination. Similarly, flow cytometric analysis of cells extracted from colorectal biopsies showed no increase in Ki67 activation or CC-chemokine receptor 5 (CCR5) expression on total or vector-specific CD4+ T cells following Ad26 vaccination. Although these data cannot guarantee the clinical safety of Ad26 vectors, it is reassuring that this vector did not detectably increase the number or activation status of total or vector-specific CD4+ T cells in colorectal mucosa in humans. However, these data do not address the hypothesis that was generated by the Step study, which used biologically very different Ad5 vectors.
Summary. Taken together, there are compelling scientific arguments for continuing to pursue alternative serotype Ads as candidate HIV-1 vaccine vectors. In particular, Ad26 is substantially different from Ad5 from both a virological and an immunological perspective. Moreover, Ad26–MVA vaccine regimens provided partial protection in the stringent SIVmac251 challenge model in rhesus monkeys in which Ad5 and DNA–Ad5 vaccine regimens failed, and similar protection against acquisition of infection has been observed in the stringent SHIV–SF162P3 challenge model. In addition, Ad26 did not result in increased numbers or activation status of total or vector-specific CD4+ T cells in colorectal mucosa in humans in a randomized, double-blinded, placebo-controlled clinical trial. Ad4, Ad35 and several chimpanzee Ads are also being evaluated in humans as candidate HIV-1 vaccine vectors.
CMV vectors
Conventional T cell-targeted vaccine approaches, including the use of non-replicating and/or non-persistent viral vectors and DNA vaccines, have only a limited ability to manifest effective antiviral effector T cell activity immediately after the onset of HIV or SIV infection before systemic spread when the virus is most vulnerable. This is due to the fact that these vaccines are generally unable to maintain the high-frequency effector-differentiated HIV- or SIV-specific T cells at the portal of HIV or SIV entry and sites of early spread that are thought to be necessary for viral control. By contrast, we have shown that rhesus CMV (RhCMV) vectors, which persist in the vaccinated host, indefinitely maintain systemic high-frequency, circulating and tissue-resident effector memory T cell (TEM cell) responses that have immediate antiviral effector activity and thereby control, and potentially clear, stringent SIV challenges in approximately one-half of vaccinated rhesus monkeys early after exposure45,46,47. In addition, RhCMV vectors can be repeatedly used in CMV-positive rhesus monkeys without any inhibition of immunogenicity by pre-existing immunity. They can be programmed to elicit unusually broad CD8+ T cell responses that recognize conventional and/or unconventional epitopes (including major histocompatibility complex class II (MHC II)-restricted CD8+ T cells), can be modified to express multiple vaccine inserts, totalling 6 kb or more of exogenous sequence, using endogenous promoters to control insert expression, and can be greatly attenuated without loss of immunogenicity or efficacy45,46,48,49. Thus, CMV vectors and the TEM cell responses that they elicit offer a powerful new approach to the development of an HIV/AIDS vaccine.
Efficacy of CMV-elicited effector memory T cells. Conventional vaccine-elicited T cell memory (so-called central memory T cell (TCM cell), or TCM cell-biased, responses) requires that, following initial pathogen encounter, the virus-specific T cells undergo a phase of expansion, effector differentiation and migration to sites of infection before mediating meaningful antiviral activity, and these responses are thus unable to intervene sufficiently early in HIV-1 and SIV infection to provide effective virological control. This limitation supports the idea of an alternative, conceptually distinct, T cell vaccine concept — that of a TEM cell vaccine. TEM cells lack the robust capacity for expansion that is used by TCM cells to mediate a potent anamnestic response but are constitutively localized in effector sites and poised for immediate effector function47,50,51,52,53,54. Indeed, as CD4+ TEM cells are the primary targets of transmitted HIV-1 and SIV55, and as CD4+ TEM and CD8+ TEM cells inhabit the same sites, a vaccine- generated CD8+ TEM cell response would theoretically be ideally positioned to intercept initial or early viral replication in primary infection, which would provide antiviral effector activity during the most vulnerable phase of infection.
Long-term maintenance of TEM cell populations is associated with persistent antigen, and T cell responses to chronic or persistent agents — particularly CMV — are heavily TEM cell-biased47,52,56,57,58. We therefore evaluated RhCMV vectors that express SIV antigens to assess the ability of SIV-specific TEM cells to intervene early in primary SIV infection. RhCMV can be modified to express high levels of SIV proteins, without disruption of other RhCMV genes and with preservation of wild-type replication characteristics45,47. These vectors efficiently re-infect RhCMV seropositive rhesus monkeys and, in the process of re-infection, elicit an indefinitely persistent, polyfunctional, high-frequency, highly TEM cell-biased and extraordinarily broad SIV-specific CD4+ T cell and CD8+ T cell response45,46,47,49. Necropsy analysis confirmed that, in the long-term steady state, these SIV-specific T cells are widely distributed in the body and are present at high frequencies in effector sites, including mucosal sites that function as portals of HIV and/or SIV entry45,46.
These RhCMV/SIV vectors do not elicit major SIV-specific antibody responses, nor do they even seem to prime for such responses45,46,47. So far, their efficacy has been assessed in a total of 64 rhesus monkeys that have been vaccinated with RhCMV/SIV Gag–Pol–Rev–Nef–Tat–Env vectors alone or in combination with Ad5 vectors, compared with 48 controls that were either unvaccinated or vaccinated with RhCMV vectors with irrelevant inserts45,46 (L.J.P., unpublished observations). These monkeys were challenged with highly pathogenic SIV by repeated, limited-dose exposure via either an intrarectal or intravaginal route until SIV infection was confirmed by virological and/or immunological criteria45,46, with efficacy defined by virological outcome after documented acquisition of SIV infection. As shown in FIG. 2 for intrarectal challenge, whereas all controls manifested progressive infection with high viral loads, slightly more than one-half of the RhCMV/SIV vaccinated animals (25 of 48 for intrarectal challenge; 34 of 64, including both intrarectal and intra-vaginal challenge) manifested an unprecedented pattern of control. Infection in these animals was characterized by a variably sized burst, or blip, of viraemia, followed by control of plasma viraemia to undetectable levels. Some of these monkeys manifested additional, low-level viral blips but, except for two animals that demonstrated relapse of infection (one progressor and one elite controller), these blips generally waned over time and were not observed in any animals 70 weeks after infection.

Plasma viral load profiles of 48 RhCMV/SIV vector-vaccinated rhesus monkeys and 30 unvaccinated rhesus monkeys after infection by repeated, limiting dose, intrarectal SIVmac239 challenge, with the day of infection defined as the challenge before the first above-threshold plasma viral load. The figure shows our composite experience to date with intrarectal SIVmac239 challenge of monkeys that have either been vaccinated twice with a strain 68-1 RhCMV/SIV vector set expressing SIVgag, SIVpol, SIVretanef, and SIVenv (week 0 and week 14; n = 36) or once with these vectors (week 0) and were then vaccinated at week 14 with Ad5 vectors expressing SIVgag, SIVpol, SIVnef, and SIVenv (n = 12); all monkeys were challenged at least 1 year following initial vaccination. The 25 initially protected animals (that is, the controllers) had an initial plasma viral blip followed by complete control except for periodic plasma viral blips that waned over time. Of 25 initially protected animals, one relapsed with progressive infection and the remainder showed long-term control and, in many of these monkeys, documented clearance of the SIV infection46. Vaccinated non-controllers were indistinguishable from unvaccinated controls and showed plasma viral load profiles that are consistent with typical progressive SIVmac239 infection. Note that one unvaccinated control monkey had an initially occult infection that spontaneously progressed 16 weeks after infection. Data from Refs 45, 46 and L.J.P., unpublished observations.
Protection was binary — that is, it was either complete or there was none at all — and was not associated with an anamnestic T cell response, which indicates that the SIV-specific T cells that were available at the time of infection were either sufficient or insufficient to control infection45,46,47. Using ultrasensitive nested PCR and co-culture assays, low levels of SIV could be detected at the sites of entry (that is, the rectum and colon) and of early lymphatic spread (in the draining lymph nodes) and haematogenous spread (in the liver, spleen and bone marrow) in 'protected' animals at necropsy 3–6 weeks after infection. Longitudinal analysis of bone marrow biopsies indicated that SIV RNA may persist in this tissue for up to ∼20 weeks. Thereafter, tissue SIV is increasingly difficult to find even with the most sensitive tests and even after CD8+ T cell depletion46. Consistent with this, T cell responses to the SIV Vif antigen, which only develop in response to SIV infection (as Vif was not included in the RhCMV/SIV vaccine), wane over time until they are no longer detectable. Indeed, after 1.5–3 years of RhCMV/SIV vector-mediated control, these animals were indistinguishable from uninfected animals by ultrasensitive nested quantitative PCR and reverse transcription PCR (RT-PCR), exhaustive co-culture analysis of tissue mononuclear cells and adoptive transfer of both blood and lymph node mononuclear cells to naive monkeys46 — findings which are strongly suggestive of immune clearance.
Taken together, these data provide strong support for the hypothesis that a highly TEM cell-biased response elicited by persistent RhCMV vectors can intercept and control stringent SIV challenges early after infection and then clear this infection over time. This protection occurs without anamnestic T cell proliferation, and we think that the observed efficacy reflects the ability of RhCMV vectors to establish and indefinitely maintain robust SIV-specific TEM cells at all potential sites of SIV replication. In addition, the breadth of RhCMV/SIV vector-elicited CD8+ T cell responses (∼threefold more different epitopes are targeted compared with conventional vaccines49) suggests that this vaccine approach will be better able to compensate for the considerable sequence heterogeneity of circulating HIV strains than other T cell vaccines.
The CMV vector-associated 'arrest and clear over time' phenotype of protection is not the same as protection against the acquisition of infection; however, it might seem to be similar in a clinical setting. Vaccinated individuals would presumably never exhibit progressive infection and might have increasingly low — and eventually no — risk of relapse over time. In addition, CMV vector-mediated protection could add to, and potentially synergize with, effective Env-specific antibody responses that are elicited by an antibody-targeted component of a vaccine, which suggests that CMV vectors could be used in combination with other vaccine platforms59. The ability of CMV vector-elicited responses to clear residual SIV infection also suggests the possibility that a CMV vector-based vaccine might be efficacious in therapeutic vaccination of patients with HIV-1 who are on antiretroviral therapy.
Novel CD8+ T cell epitope targeting by CMV vectors. The genomes of human CMV and RhCMV include a three-gene transcriptional complex (known as UL128–UL131 in human CMV and Rh157.5/.4/.6 in RhCMV) that encodes three components of an alternative entry receptor for cells other than fibroblasts (the so-called pentameric complex receptor, which also includes the gH and gL glycoproteins). These three genes are not needed for replication in fibroblasts, and one or more is frequently inactivated or lost by various genetic mechanisms from CMV strains that were multiply passaged in fibroblast culture, including the strain 68–1 RhCMV that was the starting point for our RhCMV/SIV vectors. In contrast to true wild-type (low-passage or colony-circulating) RhCMV, the UL128- and UL130-deleted strain 68–1 RhCMV replicates only poorly in cells other than fibroblasts (particularly epithelial cells), and is much less efficiently shed in vivo than true wild-type RhCMV60.
The presence or absence of these genes has a major influence on RhCMV vector immunogenicity, particularly on the number and nature of the epitopes that are recognized by RhCMV/SIV Gag vector-elicited CD8+ T cells49. It was initially noted that the SIV Gag-specific CD8+ T cells induced by the UL128- and UL130-deleted strain 68–1 RhCMV/SIV vectors failed to include any of the canonical SIV Gag epitopes recognized by CD8+ T cells that are elicited by conventional vaccines or by SIV infection itself; however, the number of different epitopes that are recognized by the RhCMV/SIV Gag vector-elicited CD8+ T cell responses were ∼threefold higher than with conventional vaccination or with vaccination with SIV itself. The breadth of the CD8+ T cell responses in the strain 68–1 RhCMV/SIVgag vector-vaccinated rhesus macaques was so great that >65% of the amino acids within the Gag protein were included in an epitope that was targeted by vector-elicited CD8+ T cells. Analysis of SIV Gag-specific CD8+ T cell responses in monkeys primed with SIV Gag-expressing conventional vectors and then boosted with strain 68–1 RhCMV/SIVgag vectors (or vice versa) showed that there was no epitope overlap between the CD8+ T cell responses that are elicited by the conventional vectors and the strain 68–1 RhCMV vector. Further analysis showed that approximately two-thirds of the SIV Gag epitopes that were recognized by strain 68–1 RhCMV/SIVgag vector-elicited CD8+ T cells were restricted by MHC II molecules, which are antigen-presenting molecules that are not usually involved in CD8+ T cell responses, rather than MHC I molecules, which typically restrict CD8+ T cell responses. Furthermore, many epitopes (that is, supertopes) were recognized in most of the outbred macaques that were studied. Importantly, both the MHC I- and MHC II-restricted CD8+ T cells that were elicited by the strain 68–1 RhCMV/SIVgag vector recognized SIV-infected T cells, which shows that these epitopes are naturally presented on virally infected cells. However, SIV infection and conventional vaccination were unable to elicit such T cell responses. Importantly, normal expression of the UL128–UL131 gene complex completely reverses the unconventional epitope targeting of RhCMV/SIVgag vectors, as strain 68–1.2 RhCMV/SIVgag vectors (with 'repaired' UL128 and UL130 expression) show conventional CD8+ T cell epitope targeting (all MHC I-restricted and no supertopes).
Perspectives
Novel serotype Ad vectors and CMV vectors are two potentially promising vector platforms to expand the current HIV-1 vaccine pipeline. They both provide partial protection in stringent rhesus monkey challenge models but manifest this protection by distinct, and potentially complementary, mechanisms of action. Novel serotype Ad vectors — particularly when boosted by poxvirus vectors or purified Env proteins — provide antibody-mediated partial protection against acquisition of infection as well as T cell-mediated virological control following infection. By contrast, CMV vectors do not protect against acquisition of infection, but they provide robust virological control and potential virological clearance in approximately one-half of vaccinated animals as a result of TEM cell responses. Clinical safety considerations for novel serotype Ad vectors include the possibility that Ad5 vectors may have increased the risk of acquisition in previous HIV-1 vaccine trials. Clinical safety considerations for CMV vectors include their persistence and the potential pathogenicity of wild-type CMV. These issues are being actively evaluated, as advanced clinical development is planned for novel serotype Ad vectors and first-in-human clinical trials are planned for CMV vectors. The possibility of combining these immunologically very different, but potentially complementary, vaccine platforms is also being explored.
Change history
12 October 2017
On page 768 of the article, the text has been changed to clarify that all vaccinated and control animals were challenged until documentation of SIV infection by virological and/or immunological criteria. Furthermore, on page 768, the text has been changed to specify both the number of vaccinated controllers shown in Figure 2 after intrarectal challenge as well as the number of controllers after both intrarectal and intravaginal challenge. Finally, the legend of Figure 2 has been changed to clarify that the graph shows intrarectal SIVmac239 challenge of monkeys that have either been vaccinated twice with a strain 68-1 RhCMV/SIV vector (week 0 and week 14; n = 36) or once with these vectors (week 0) and were then vaccinated at week 14 with Ad5 vectors (n = 12). Changes have been made to both the HTML and PDF versions of the article. The authors apologize to readers for any confusion caused.
References
Barouch, D. H. The quest for an HIV-1 vaccine — moving forward. N. Engl. J. Med. 369, 2073–2076 (2013).
Fauci, A. S. & Marston, H. D. Ending AIDS — is an HIV vaccine necessary? N. Engl. J. Med. 370, 495–498 (2014).
Buchbinder, S. P. et al. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): a double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 372, 1881–1893 (2008).
Flynn, N. M. et al. Placebo-controlled phase 3 trial of a recombinant glycoprotein 120 vaccine to prevent HIV-1 infection. J. Infect. Dis. 191, 654–665 (2005).
Gray, G. E. et al. Safety and efficacy of the HVTN 503/Phambili study of a clade-B-based HIV-1 vaccine in South Africa: a double-blind, randomised, placebo-controlled test-of-concept phase 2b study. Lancet Infect. Dis. 11, 507–515 (2011).
Hammer, S. M. et al. Efficacy trial of a DNA/rAd5 HIV-1 preventive vaccine. N. Engl. J. Med. 369, 2083–2092 (2013).
Pitisuttithum, P. et al. Randomized, double-blind, placebo-controlled efficacy trial of a bivalent recombinant glycoprotein 120 HIV-1 vaccine among injection drug users in Bangkok, Thailand. J. Infect. Dis. 194, 1661–1671 (2006).
Rerks-Ngarm, S. et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N. Engl. J. Med. 361, 2209–2220 (2009).
Shiver, J. W. & Emini, E. A. Recent advances in the development of HIV-1 vaccines using replication-incompetent adenovirus vectors. Annu. Rev. Med. 55, 355–372 (2004).
Shiver, J. W. et al. Replication-incompetent adenoviral vaccine vector elicits effective anti-immunodeficiency-virus immunity. Nature 415, 331–335 (2002).
Casimiro, D. R. et al. Attenuation of simian immunodeficiency virus SIVmac239 infection by prophylactic immunization with DNA and recombinant adenoviral vaccine vectors expressing Gag. J. Virol. 79, 15547–15555 (2005).
Wilson, N. A. et al. Vaccine-induced cellular immune responses reduce plasma viral concentrations after repeated low-dose challenge with pathogenic simian immunodeficiency virus SIVmac239. J. Virol. 80, 5875–5885 (2006).
Benlahrech, A. et al. Adenovirus vector vaccination induces expansion of memory CD4 T cells with a mucosal homing phenotype that are readily susceptible to HIV-1. Proc. Natl Acad. Sci. USA 106, 19940–19945 (2009).
Hutnick, N. A. et al. Baseline Ad5 serostatus does not predict Ad5 HIV vaccine-induced expansion of adenovirus-specific CD4+ T cells. Nature Med. 15, 876–878 (2009).
McElrath, M. J. et al. HIV-1 vaccine-induced immunity in the test-of-concept Step Study: a case-cohort analysis. Lancet 372, 1894–1905 (2008).
O'Brien, K. L. et al. Adenovirus-specific immunity after immunization with an Ad5 HIV-1 vaccine candidate in humans. Nature Med. 15, 873–875 (2009).
Letvin, N. L. et al. Immune and genetic correlates of vaccine protection against mucosal infection by SIV in monkeys. Sci. Transl. Med. 3, ra36 (2011).
Lopker, M. et al. Heterogeneity in neutralization sensitivities of viruses comprising the simian immunodeficiency virus SIVsmE660 isolate and vaccine challenge stock. J. Virol. 87, 5477–5492 (2013).
Barouch, D. H. et al. International seroepidemiology of adenovirus serotypes 5, 26, 35, and 48 in pediatric and adult populations. Vaccine 29, 5203–5209 (2011).
Mast, T. C. et al. International epidemiology of human pre-existing adenovirus (Ad) type-5, type-6, type-26 and type-36 neutralizing antibodies: correlates of high Ad5 titers and implications for potential HIV vaccine trials. Vaccine 28, 950–957 (2010).
Abbink, P. et al. Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J. Virol. 81, 4654–4663 (2007).
Baden, L. R. et al. First-in-human evaluation of the safety and immunogenicity of a recombinant adenovirus serotype 26 HIV-1 Env vaccine (IPCAVD 001). J. Infect. Dis. 207, 240–247 (2013).
Barouch, D. H. et al. Characterization of humoral and cellular immune responses elicited by a recombinant adenovirus serotype 26 HIV-1 Env vaccine in healthy adults (IPCAVD 001). J. Infect. Dis. 207, 248–256 (2013).
Creech, C. B. et al. Randomized, placebo-controlled trial to assess the safety and immunogenicity of an adenovirus type 35-based circumsporozoite malaria vaccine in healthy adults. Hum. vaccin. Immunother. 9, 2548–2557 (2013).
Keefer, M. C. et al. A phase I double blind, placebo-controlled, randomized study of a multigenic HIV-1 adenovirus subtype 35 vector vaccine in healthy uninfected adults. PLoS ONE 7, e41936 (2012).
Vogels, R. et al. Replication-deficient human adenovirus type 35 vectors for gene transfer and vaccination: efficient human cell infection and bypass of preexisting adenovirus immunity. J. Virol. 77, 8263–8271 (2003).
Antrobus, R. D. et al. Clinical assessment of a novel recombinant simian adenovirus ChAdOx1 as a vectored vaccine expressing conserved Influenza A antigens. Mol. Ther. 22, 668–674 (2013).
Barnes, E. et al. Novel adenovirus-based vaccines induce broad and sustained T cell responses to HCV in man. Sci. Transl. Med. 4, ra111 (2012).
Colloca, S. et al. Vaccine vectors derived from a large collection of simian adenoviruses induce potent cellular immunity across multiple species. Sci. Transl. Med. 4, ra112 (2012).
O'Hara, G. A. et al. Clinical assessment of a recombinant simian adenovirus ChAd63: a potent new vaccine vector. J. Infect. Dis. 205, 772–781 (2012).
Li, H. et al. Adenovirus serotype 26 utilizes CD46 as a primary cellular receptor and only transiently activates T lymphocytes following vaccination of rhesus monkeys. J. Virol. 86, 10862–10865 (2012).
Waddington, S. N. et al. Adenovirus serotype 5 hexon mediates liver gene transfer. Cell 132, 397–409 (2008).
Perreau, M. et al. The number of Toll-like receptor 9-agonist motifs in the adenovirus genome correlates with induction of dendritic cell maturation by adenovirus immune complexes. J. Virol. 86, 6279–6285 (2012).
Quinn, K. M. et al. Comparative analysis of the magnitude, quality, phenotype, and protective capacity of simian immunodeficiency virus gag-specific CD8+ T cells following human-, simian-, and chimpanzee-derived recombinant adenoviral vector immunization. J. Immunol. 190, 2720–2735 (2013).
Teigler, J. E., Iampietro, M. J. & Barouch, D. H. Vaccination with adenovirus serotypes 35, 26, and 48 elicits higher levels of innate cytokine responses than adenovirus serotype 5 in rhesus monkeys. J. Virol. 86, 9590–9598 (2012).
Penaloza-MacMaster, P. et al. Alternative serotype adenovirus vaccine vectors elicit memory T cells with enhanced anamnestic capacity compared to Ad5 vectors. J. Virol. 87, 1373–1384 (2013).
Tan, W. G. et al. Comparative analysis of simian immunodeficiency virus gag-specific effector and memory CD8+ T cells induced by different adenovirus vectors. J. Virol. 87, 1359–1372 (2013).
Liu, J. et al. Magnitude and phenotype of cellular immune responses elicited by recombinant adenovirus vectors and heterologous prime–boost regimens in rhesus monkeys. J. Virol. 82, 4844–4852 (2008).
Barouch, D. H. et al. Vaccine protection against acquisition of neutralization-resistant SIV challenges in rhesus monkeys. Nature 482, 89–93 (2012).
Barouch, D. H. et al. Mosaic HIV-1 vaccines expand the breadth and depth of cellular immune responses in rhesus monkeys. Nature Med. 16, 319–323 (2010).
Fischer, W. et al. Polyvalent vaccines for optimal coverage of potential T-cell epitopes in global HIV-1 variants. Nature Med. 13, 100–106 (2007).
Santra, S. et al. Mosaic vaccines elicit CD8+ T lymphocyte responses that confer enhanced immune coverage of diverse HIV strains in monkeys. Nature Med. 16, 324–328 (2010).
Barouch, D. H. et al. Protective efficacy of a global HIV-1 mosaic vaccine against heterologous SHIV challenges in rhesus monkeys. Cell 155, 531–539 (2013).
Baden, L. R. et al. Induction of HIV-1-specific mucosal immune responses following intramuscular recombinant adenovirus serotype 26 HIV-1 vaccination of humans. J. Infect. Dis. http://dx.doi.org/10.1093/infdis/jiu485 (2014).
Hansen, S. G. et al. Profound early control of highly pathogenic SIV by an effector memory T-cell vaccine. Nature 473, 523–527 (2011).
Hansen, S. G. et al. Immune clearance of highly pathogenic SIV infection. Nature 502, 100–104 (2013).
Hansen, S. G. et al. Effector memory T cell responses are associated with protection of rhesus monkeys from mucosal simian immunodeficiency virus challenge. Nature Med. 15, 293–299 (2009).
Hansen, S. G. et al. Evasion of CD8+ T cells is critical for superinfection by cytomegalovirus. Science 328, 102–106 (2010).
Hansen, S. G. et al. Cytomegalovirus vectors violate CD8+ T cell epitope recognition paradigms. Science 340, 1237874 (2013).
Bannard, O., Kraman, M. & Fearon, D. Pathways of memory CD8+ T-cell development. Eur. J. Immunol. 39, 2083–2087 (2009).
Genesca, M., McChesney, M. B. & Miller, C. J. Antiviral CD8+ T cells in the genital tract control viral replication and delay progression to AIDS after vaginal SIV challenge in rhesus macaques immunized with virulence attenuated SHIV 89.6. J. Intern. Med. 265, 67–77 (2009).
Picker, L. J. et al. IL-15 induces CD4 effector memory T cell production and tissue emigration in nonhuman primates. J. Clin. Invest. 116, 1514–1524 (2006).
Pitcher, C. J. et al. Development and homeostasis of T cell memory in rhesus macaque. J. Immunol. 168, 29–43 (2002).
Sallusto, F., Geginat, J. & Lanzavecchia, A. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu. Rev. Immunol. 22, 745–763 (2004).
Grossman, Z., Meier-Schellersheim, M., Paul, W. E. & Picker, L. J. Pathogenesis of HIV infection: what the virus spares is as important as what it destroys. Nature Med. 12, 289–295 (2006).
Champagne, P. et al. Skewed maturation of memory HIV-specific CD8 T lymphocytes. Nature 410, 106–111 (2001).
Gauduin, M. C. et al. Induction of a virus-specific effector-memory CD4+ T cell response by attenuated SIV infection. J. Exp. Med. 203, 2661–2672 (2006).
Kern, F. et al. Distribution of human CMV-specific memory T cells among the CD8pos. subsets defined by CD57, CD27, and CD45 isoforms. Eur. J. Immunol. 29, 2908–2915 (1999).
Picker, L. J., Hansen, S. G. & Lifson, J. D. New paradigms for HIV/AIDS vaccine development. Annu. Rev. Med. 63, 95–111 (2012).
Fruh, K., Malouli, D., Oxford, K. L. & Barry, P. A. in Cytomegalovirus Vol. 2. (ed. Reddehase,M. J.) (Caister Academic Press, 2013).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
L.J.P and Oregon Health and Science University (OHSU) have a significant financial interest in TomegaVax, Inc., which is a company that may have a commercial interest in the results of this research and technology. The potential individual and institutional conflicts of interest have been reviewed and managed by OHSU. D.H.B. is a named co-inventor on various vector and antigen patents.
Glossary
- Anamnestic response
-
The enhanced immune response that occurs against an antigen as a result of previous host exposure to a related antigen.
- CD46
-
A ubiquitously expressed type-1 transmembrane protein that functions to regulate complement. It functions as a receptor for vaccine strains of measles virus and some adenovirus serotypes.
- Central memory T cell
-
(TCM cell). An antigen-experienced resting T cell that expresses cell surface receptors that are required for homing to secondary lymphoid organs. These cells are generally long-lived and can function as the precursors for effector T cells during recall responses.
- Coxsackie and adenovirus receptor
-
(CAR). An immunoglobulin-like transmembrane cell adhesion protein that is used by some coxsackievirus and adenovirus species as a receptor.
- Effector memory T cell
-
(TEM cell). An antigen-experienced T cell that maintains effector differentiation, resides in, or expresses cell surface receptors that are required for homing to, extra-lymphoid effector sites and that has limited expansion potential.
- Elite controller
-
A patient infected with HIV whose immune system can limit viral RNA to below 50 copies per ml for at least 12 months in the absence of highly active antiretroviral therapy.
- Heterologous prime–boost
-
Repeated immunization using different vaccines; it is used to stimulate a better immune response.
- Neutralizing antibodies
-
Antibodies that block infectivity (for example, of a virus), usually by binding to the foreign particle (the virion) and incapacitating it in some way.
Rights and permissions
About this article

Cite this article
Barouch, D., Picker, L. Novel vaccine vectors for HIV-1. Nat Rev Microbiol 12, 765–771 (2014). https://doi.org/10.1038/nrmicro3360
Published:
Issue Date:
DOI: https://doi.org/10.1038/nrmicro3360
This article is cited by
-
Lentiviral vector induces high-quality memory T cells via dendritic cells transduction
Communications Biology (2021)
-
RhCMV serostatus and vaccine adjuvant impact immunogenicity of RhCMV/SIV vaccines
Scientific Reports (2020)
-
Production of HIV-1-based virus-like particles for vaccination: achievements and limits
Applied Microbiology and Biotechnology (2019)
-
Bone marrow stem cells to destroy circulating HIV: a hypothetical therapeutic strategy
Journal of Biological Research-Thessaloniki (2018)
-
First Impressions—the Potential of Altering Initial Host-Virus Interactions for Rational Design of Herpesvirus Vaccine Vectors
Current Clinical Microbiology Reports (2018)
