
Abstract
The widespread reorganization of cellular architecture in mitosis is achieved through extensive protein phosphorylation, driven by the coordinated activation of a mitotic kinase network and repression of counteracting phosphatases. Phosphatase activity must subsequently be restored to promote mitotic exit. Although Cdc14 phosphatase drives this reversal in budding yeast, protein phosphatase 1 (PP1) and protein phosphatase 2A (PP2A) activities have each been independently linked to mitotic exit control in other eukaryotes1,2,3,4,5,6. Here we describe a mitotic phosphatase relay in which PP1 reactivation is required for the reactivation of both PP2A-B55 and PP2A-B56 to coordinate mitotic progression and exit in fission yeast. The staged recruitment of PP1 (the Dis2 isoform) to the regulatory subunits of the PP2A-B55 and PP2A-B56 (B55 also known as Pab1; B56 also known as Par1) holoenzymes sequentially activates each phosphatase. The pathway is blocked in early mitosis because the Cdk1–cyclin B kinase (Cdk1 also known as Cdc2) inhibits PP1 activity, but declining cyclin B levels later in mitosis permit PP1 to auto-reactivate1,7,8,9,10. PP1 first reactivates PP2A-B55; this enables PP2A-B55 in turn to promote the reactivation of PP2A-B56 by dephosphorylating a PP1-docking site in PP2A-B56, thereby promoting the recruitment of PP1. PP1 recruitment to human, mitotic PP2A-B56 holoenzymes and the sequences of these conserved PP1-docking motifs11,12 suggest that PP1 regulates PP2A-B55 and PP2A-B56 activities in a variety of signalling contexts throughout eukaryotes.
Similar content being viewed by others

Main
Cdk1–cyclin B phosphorylation of a conserved site in the carboxy terminus of PP1 depresses PP1 activity at mitotic commitment1,7,8,9,10. Declining Cdk1–cyclin B levels then allow the compromised PP1 to dephosphorylate itself to promote a return to full activity1,7. Of the two fission yeast PP1 enzymes, PP1Sds21 and PP1Dis2, only PP1Dis2 harbours the conserved inhibitory phosphorylation site2,9 (Extended Data Fig. 1a). In vitro PP1Dis2 activity assays recapitulated previous observations that T316 phosphorylation by Cdk1–cyclin B depressed activity (Fig. 1a and Extended Data Fig. 1b, c)1,7,9. Mutating T316 to aspartic acid to mimic phosphorylation reduced the activity to a similar degree to phosphorylation by Cdk1–cyclin B (Fig. 1a and Extended Data Fig. 1c). Replacement of PP1dis2+ with a PP1dis2.T316A allele increased PP1Dis2 levels, whereas they were reduced in PP1dis2.T316D (Fig. 1b), indicating that phosphorylated T316 might act as a phospho-degron. Since this interpretation conflicted with reports of stable PP1Dis2 levels throughout mitosis10,13, we monitored PP1Dis2 levels with both low and high antibody dilutions as size-selected cells synchronously transited the cell cycle. A transient reduction in PP1Dis2 levels as T316 phosphorylation peaked (Fig. 1c) was blocked when proteosome function was inhibited (Extended Data Fig. 1e). Consistently, PP1Dis2 levels were persistently low in PP1dis2.T316D and persistently high in PP1dis2.T316A (Fig. 1c and Extended Data Fig. 1f, g), indicating that phosphorylation of T316 by Cdk1–cyclin B both reduces PP1Dis2 levels and inhibits its phosphatase activity.

a, EnzChek phosphatase assays (Extended Data Fig. 1b) of PP1Dis2 isolated from the indicated strains by antibody precipitation. ‘+Cdk1–cyclin B’ indicates the addition of sepharose beads to which covalently linked p13Suc1 protein had recruited Cdk1–cyclin B from S. pombe cell extracts. NIPP1 is a highly specific PP1 inhibitor. Purvalanol (Pur.) is a Cdk1 inhibitor. b, c, PP1Dis2 and T316 phosphorylation (T316Phos) levels (Rpn12mts3.1 is a temperature-sensitive proteasome mutation). For b, n = 5 biological replicates; error bars show standard deviation (s.d.). c, Size-selected cells transit the cell division cycle. The septation profile is for the wild-type culture. n = 2 biological with two technical replicates each. PP1Dis2.T316D and PP1Dis2.T316A levels from the experiments in Extended Data Fig.1f, g are superimposed on this wild-type data set.
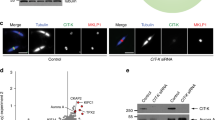
PP2A holoenzymes combine a catalytic and scaffolding subunit with one of four regulatory B subunits14, of which PP2A-B55 and PP2A-B56 have been linked to mitotic control3,4,5,15,16. We noticed that the B55 and B56 regulatory subunits had highly conserved PP1-docking-site consensus motifs (RVxF/RxVxF)12 (Fig. 2a). The Schizosaccharomyces pombe genome encodes one B55 (B55Pab1) and two B56 subunits (B56Par1 and B56Par2)17,18, and we found that both B55Pab1 and B56Par1 associated with PP1Dis2 in immunoprecipitation assays (Fig. 2b and Extended Data Fig. 2a–d). B56Par1 also bound PP1Dis2 in a yeast two-hybrid assay (Extended Data Fig. 3), and the functional replacement of the PP1-docking site of the morphogenesis regulator Wsh3 (also known as Tea4)19 by the SKEVLF motif of B56Par1 confirmed its ability to recruit PP1 (Extended Data Fig. 4a–d). The interaction between PP1Dis2 and B55Pab1 was abolished by mutating the PP1-docking consensus motif (Fig. 2b and Extended Data Fig. 2a). No association was found between PP1Sds21 and any regulatory subunit (Extended Data Fig. 2b, c), nor between B56Par2 and PP1Dis2 (Extended Data Figs 2d and 3). L482 of B56Par2 occupies a position occupied by only valine or isoleucine in validated PP1-docking sites12. Changing this leucine to valine allowed PP1Dis2 to bind B56Par2 (Extended Data Fig. 2e).

a, Clustal W alignments of PP1-docking motifs in B55 and B56 molecules. Consensus residues are in bold. b, f, g, Immunoprecipitation (IP) reactions probed with PP1Dis2(ref. 20) and T316Phos polyclonal and 12CA5 and 366 monoclonal (HA and Pk epitopes, respectively) antibodies. Asynch., asynchronous. Asterisk indicates PP1dis2.T316A. f, Mitotic progression was scored by tubulin staining with telophase defined by post-anaphase arrays of microtubules. b, n = 6; error bars show s.d. c, d, 4′,6-Diamidino-2-phenylindole (DAPI) staining. e, Tubulin immunofluorescence staining for Cdk1cdc2.33 ‘arrest/release’ experiments scored mitotic progression. Biological replicates: for b, n = 6; for e–g, n = 2.
Core K/RxVxF PP1-docking motifs can be accompanied by secondary motifs12. Although the B55Pab1 docking site is an isolated K/RxVxF motif, the GLLR sequence of B56Par1 bears a striking resemblance to the secondary element G/SILK/R11,12 (Fig. 2a, green box). Mutating the GLLR motif (B56par1.G367V) mimicked mutating the R/KxVxF motif (B56par1.K379AV381AF383A) in abolishing interactions between PP1Dis2 and B56Par1 in two-hybrid assays (Extended Data Fig. 3), and in compromising their ability to co-immunoprecipitate (Fig. 2b and Extended Data Fig. 2a). Mutating both motifs had an additive effect, severely compromising the affinity of B56Par1 for PP1Dis2 in co-immunoprecipitation assays (Fig. 2b and Extended Data Fig. 2a). Finally, the GLLR motif efficiently substituted for the characterized PP1-binding GILK motif of Cut12 (Extended Data Fig. 4e–g)20. We conclude that GLLR is a ‘G/SILK/R’ motif.
To confirm that the motifs we identified represent genuine PP1Dis2-docking sites, we generated PP1-docking-site-null mutants (PDSN) of B55pab1 (R52A/V54A/F56A) and B56par1 (G367V/K379A/V381A/F383A), and purified the respective holoenzymes and wild-type controls from yeast cultures. The purified proteins were mixed with purified PP1Dis2 and the PP1Dis2 re-isolated to identify interacting partners. In each case, wild-type but not PDSN holoenzymes were captured by PP1Dis2 (Extended Data Fig. 5a–d). Moreover, PP1Dis2 co-migrated with the largest form of each of the wild-type PP2A holoenzyme complexes in size-exclusion chromatography (Extended Data Fig. 5e). We conclude that both B56Par1 and B55Pab1 contain genuine PP1Dis2-docking motifs.
To assess the mitotic defects of the B55pab1.PDSN and B56par1.PDSN PP1-docking site mutants (Fig. 2c, d), we exploited transient arrest at the G2/M boundary with the temperature-sensitive Cdk1cdc2.33 mutation to synchronize mitotic progression21 (Fig. 2e). Blocking PP1Dis2 recruitment to either B55Pab1 or B56Par1 in synchronized divisions generated major errors in chromosome segregation and delayed the metaphase/anaphase transition (Fig. 2e and Extended Data Fig. 6a). To assay the timing of PP1Dis2 association with each PP2A holoenzyme, we used a Cdk1cdc2.33 strain in which B55Pab1 and B56Par1 were fused to different epitope tags, SV5 P protein (Pk) and haemagglutinin (HA) (Cdk1cdc2.33, B55pab1.Pk and B56par1.HA). PP1Dis2 levels in B55Pab1.Pk precipitates peaked at metaphase (25–35 min) whereas PP1Dis2 levels in B56Par1.HA immunoprecipitates peaked later as spindles disassembled in telophase (60–70 min) (Fig. 2f).
An established assay for PP2A-B56Par1 (ref. 22), in which PP1Dis2 displayed no activity (Extended Data Fig. 6b–d), revealed a mitotic decline in bulk PP2A-B56Par1 activity before recovery at the start of the next cycle (Fig. 3a). PP2A-B56Par1 activity was severely compromised when either the GLLR (GILK) or KEVLF (RxVxF) motifs were individually mutated (Extended Data Fig. 6e, f), and abolished when both were simultaneously mutated to disrupt PP1Dis2 recruitment (Fig. 3b and Extended Data Fig. 6g). This indicated that PP1Dis2 could activate PP2A-B56Par1; we confirmed this by adding immunopurified PP1Dis2 to PP2A-B56Par1 that had been isolated by immunoprecipitation from synchronized cells that lacked PP1Dis2 (PP1dis2.Δ) (Fig. 3c). Moreover, PP1Dis2 activated wild-type PP2A-B56Par1 but not the PP1-docking-site mutant (Fig. 3c, d and Extended Data Fig. 6g, h), and genetically inhibited PP1Dis2.T316D was unable to activate PP2A-B56Par1 (Fig. 3c and Extended Data Fig. 6h). Similar docking-site-dependent activity enhancement of purified PP2A-B56Par1 enzymes by purified PP1Dis2 (Extended Data Fig. 5f) further confirmed that recruitment of active PP1Dis2 to the PP2A-B56Par1 docking site reactivated PP2A-B56Par1.

a, b, g–i, Assays of PP2A-B56Par1 activity after B56Par1.HA immunoprecipitation (with 12CA5 antibodies) of Cdk1cdc2.33 arrest/release synchronized cultures followed for one (b, g–i) or two (a) cycles. Activity changes (loss of GST–Rec8391–561 32P radioactivity22) at each point were normalized to the activity of B56Par1.HA from an asynchronous (Asynch.) culture processed in parallel on the same gel (penultimate lane). sub, phosphorylated substrate. c, d, Aliquots of PP1Dis2 protein immunoprecipitates (isolated under high salt (1.2 M NaCl) extraction conditions to disassociate partners) from Cdk1cdc2.33 arrested (interphase) cultures were added to PP2A-B56Par1 assays from Cdk1cdc2.33 dis2.Δ cultures (Extended Data Fig. 6). e, f, Co-immunoprecipitation assays as for Fig. 2b. Asterisks indicate extracts from asynchronous B56par1.HA cultures. j, Phenotype analysis of Cdk1cdc2.33 synchronized mitoses as for Fig. 2e. Biological replicates: for a, n = 3; for all other panels, n = 2.
Phosphorylation between the G/SILK/R and RxVxF motifs of a bipartite docking site can block PP1 recruitment20. Mass spectrometry analysis of B56Par1 from mitotic cells appeared to indicate phosphorylation on S378 between the GLLR and KEVLF motifs (boxed residue in Fig. 2a; data not shown). Attempts to generate antibodies to recognize B56Par1 when phosphorylated on S378 failed, prompting us to generate a polyclonal antibody that would recognize B56Par1 when phosphorylated on both S377 and S378 (Extended Data Fig. 7a). This antibody (B56-Phos) revealed increasing phosphorylation in mitosis that declined as PP1Dis2 was recruited to PP2A-B56Par1 ( Fig. 4a). Mutating S378 to alanine to block phosphorylation (B56par1.S378A) promoted persistent PP1Dis2 recruitment to PP2A-B56Par1 and persistently high levels of PP2A-B56Par1 activity throughout mitosis that depended upon the presence of PP1Dis2 (Fig. 3e, g, h and Extended Data Fig. 6i, j). Conversely, when S378 was mutated to aspartic acid to mimic phosphorylation (B56par1.S378D), this severely compromised PP1Dis2 recruitment and B56Par1.S378D activity remained low throughout division (Fig. 3f, i and Extended Data Fig. 6k). Anaphase was delayed and frequently abnormal in B56par1.S378D cells (Fig. 3j), and it was striking that the persistent mitotic association of PP1Dis2 with PP2A-B56Par1 in B56par1.S378A cells generated similar mitotic errors (Fig. 3j). Thus, dynamic phosphate turnover on S378 appears to be crucial for orderly mitotic progression. As anticipated from the match to the consensus phosphorylation site for the human polo-like kinase PLK1 (N/D/ExS/T)23, PoloPlo1 was solely responsible for S378 phosphorylation (Fig. 4b, c).

a, e–i, Cdk1cdc2.33 synchronized mitoses. a–c, e, B56-Phos antibodies (Extended Data Fig. 7a) detected phosphorylation on B56Par1 after immunoprecipitation from the indicated strains and the indicated treatments. PP1Dis2 levels were monitored where indicated. λ phosph., λ phosphatase; 3BrB-PP1, 3-[(3-bromophenyl)methyl]-1-tert-butylpyrazolo[3,4-d]pyrimidin-4-amine; PoloPlo1.KDHA, HA-tagged catalytically inactive; inhib., inhibitor. d, B56Par1 precipitates probed for PP1Dis2. f–i, PP2A-B55Pab1 phosphatase assays (Extended Data Fig. 7c–f). j, Bulk activities are illustrated: local activities at any given time will depend on the specific balance of each activity at a particular site. (1) Cdk1–cyclin B activation represses all three phosphatase activities (Extended Data Fig. 9a). Direct phosphorylation of PP1Dis2 TPRR motif inhibits catalytic activity1,7,8,9,10 and promotes destruction. PP1Dis2 binds B55Pab1 but cannot reactivate PP2A-B55Pab1 while TPRR phosphorylation persists. PoloPlo1 phosphorylation of B56Par1 prevents PP1Dis2 recruitment to PP2A-B56Par1. (2) Declining Cdk1–cyclin B activity facilitates PP1 auto-reactivation. (3) Re-activated PP1Dis2 promotes PP2A-B55Pab1 reactivation. (4) PP2A-B55Pab1 dephosphorylates the PP1-docking site in B56Par1 less efficiently than PoloPlo1 phosphorylates it, keeping PP2A-B56Par1 activity low. (5) Declining PoloPlo1 activity enables PP2A-B55Pab1 to dephosphorylate B56Par1 and promote reactivating recruitment of PP1Dis2 (Extended Data Fig. 8). k, Flag-tagged B56γ and B56δ isoforms were stably expressed in HeLa-FRT cell lines, immunoprecipitated from mitotic cells isolated by shake-off from nocodazole-treated cells, and the immunoprecipitates probed with an antibody that recognizes all forms of PP1. Biological replicates: for k, n = 3; for all other panels, n = 2.
PP1Dis2 recruitment to B56Par1 at telophase (Fig. 2f) indicated that S378 should be removed at this time, and the inability of B55pab1.Δ cells to recruit PP1Dis2 to B56Par1 (Fig. 4d) indicated that PP2A-B55Pab1 could be responsible for this dephosphorylation. In support of this, PP2A-B55Pab1 removed B56-Phos reactivity from B56Par1 in an in vitro assay (Extended Data Fig. 7b). This phosphatase activity peaked during mitosis and was abolished when PP1Dis2 recruitment to B55Pab1 was blocked by removal of PP1Dis2, when the PP1-docking site in B55Pab1 was ablated, and when genetically inhibited PP1dis2.T316D was recruited (Fig. 4f–i and Extended Data Fig. 7c–f). Consistently, the failure of PP1Dis2 to associate with PP2A-B56Par1 in B55pab1.Δ cells correlated with persistent phosphorylation on S378 throughout mitosis (B55pab1.Δ; Fig. 4e). Finally, purified PP1Dis2 enhanced the B56-Phos activity of purified PP2A-B55Pab1 in a docking-site-dependent manner (Extended Data Fig. 5f).
In summary, PP1Dis2, PP2A-B55Pab1 and PP2A-B56Par1 are linked in a phosphatase relay (Fig. 4j and Extended Data Fig. 8). At mitotic commitment all three activities are repressed4 (Extended Data Fig. 9a); subsequently PP1Dis2 activity recovers auto-catalytically as cyclin B is degraded. The binding of repressed, T316-phosphorylated PP1Dis2 to PP2A-B55Pab1 (Fig. 2g) promotes rapid restoration of PP2A -B55Pab1 activity upon reduction of Cdk1–cyclin B activity. Reactivated PP2A-B55Pab1 can then begin to dephosphorylate S378 of B56Par1 but is antagonized by PoloPlo1activity towards this site. Consequently, PP1Dis2 can only be recruited to PP2A-B56Par1 at telophase when PoloPlo1 activity declines (Fig. 4j and Extended Data Fig. 8). Thus, Cdk1–cyclin B phosphorylation of PP1 at mitotic commitment1,7,8,9,10 locks all three phosphatases in the ‘off’ state. This lock is bolted through a Cdk1–cyclin B phosphorylation-mediated reduction of PP1Dis2 levels. These controls join a range of other modes of PP1 and PP2A regulation, including Greatwall/ENSA control of PP2A-B55δ, and PP2A-B56 regulation by Bod1, to provide the accurate control of phosphorylation status that is critical for faithful cell division1,6,11,24,25.
The conservation of the PP1-docking-site motifs in PP2A regulatory subunits (Fig. 2a) prompted us to investigate the association between human PP1 and PP2A enzymes. PP1α, β and γ bound B56γ in yeast two-hybrid assays, and PP1 co-immunoprecipitated with Flag epitope-tagged B56γ and B56δ expressed in mitotic human cells. Each of these interactions was abolished by mutation of the PP1-docking-site motif (Fig. 4k and Extended Data Fig. 9b–d). Thus, the principle of a phosphatase relay, whereby PP1 recruitment activates PP2A to control mitosis, is conserved in human cells. It is notable that the NXS Polo kinase consensus sequences in fungal B56 subunits are replaced by SP and SQ motifs in metazoa (Fig. 2a), suggesting that CDK/MAP kinases modulate PP1 docking to B56δ and B56γ, whereas ATM/ATR/DNA-PK may coordinate docking to B56 α, β, ε. Furthermore, these serine/threonine residues also conform to consensus motifs for AGC kinases, including aurora B (KXXS/T), while the threonine highlighted in yellow in Fig. 2a could facilitate control by further kinases. We believe, therefore, that PP1 regulation of PP2A-B56 activity will emerge as a ubiquitous feature in diverse signalling contexts. Although bulk biochemical assays reveal core principles of phosphatase control, local controls sit at the heart of many transitions26. Thus, we propose that competition between kinases and PP1 and PP2A holoenzymes at specific locations will generate locally active, dephosphorylated phosphatases at times when assays show the activity of the bulk population is depressed.
The direct recruitment of PP1 to B55 and B56 subunits that we observed is unanticipated. Although the majority of predicted PP1-docking sites reside within regions that are predicted to be intrinsically disordered11, PP1 recruitment to the docking sites of B55Pab1, B56Par1 and CENP-E11,27 suggest that this requirement is not axiomotic. A second conundrum is that the PP1-docking motifs are inaccessible in structures of the PP2A holoenzyme complexes that have been solved so far28,29,30. PP1-docking site residues, and the equivalent residue to S378, form bonds that are crucial to the integrity of these structures, yet, contrary to predictions arising from these structures, mutation of these residues to alanine has no impact on the integrity of either PP2A-B55 or PP2A-B56 holoenzymes (Extended Data Fig. 10). Moreover, S378 is phosphorylated in purified PP2A-B56Par1 complexes and holoenzymes isolated by one-step immunoprecipitation (Extended Data Figs 5g and 10c). Our data therefore add to established discrepancies between in vivo and in vitro PP2A holoenzyme assembly24 to suggest that alternative structures will emerge from structural studies of active enzymes that have undergone the extensive array of modifications required for activity that impact upon PP2A conformation in vivo24.
Methods
Standard procedures31,32 were used to grow and maintain yeast strains (Supplementary Table 1). Appropriately supplemented EMM2 synthetic medium was used throughout with the exception of TAP tag purification, for which we used 10× YES. The generation of size-selected synchronous cultures was as described previously33. Two-hybrid assays used the matchmaker gold system (Clontech 630489). ‘Phospho-specific’ rabbit polyclonal antibodies were generated by Eurogentec. Commercial antibodies detected CPpa1 and CPpa2 (Cell Signaling, 2038), APaa1 (Abcam, ab28350) and human B56ε (Santa Cruz sc-135223). In house polyclonal antibodies detected PP1Dis2(ref. 20) and green fluorescent protein (GFP). To detect associations between PP1Dis2 and B55Pab1 and PP1Dis2 and B56Par1 the complexes were isolated from 2 × 108 cells in the buffer (50 mM HEPES (pH 7.5), 50 mM NaF, 0.4 mM Na3VO4, 40 mM Na–β-glycerol phosphate, 1 mM EDTA, 50 mM NaCl, 0.1% NP40, 1 mM PMSF). The EnzChek (Molecular Probes E12020) or histone H3 serine 10 dephosphorylation assays34 monitored PP1Dis2 activity (Extended Data Fig. 1) with the use of recombinant rabbit PP1γ (NEB P0754S), full-length NIPP1 (Abcam, ab131705), Phos-Stop (Roche 04906845001) and potato acid phosphatase (Molecular Probes E12020). Manufacturer’s instructions were followed for EnzChek assays (Molecular Probes E12020). For Histone H3 serine 10 assays, histone H3 (Millipore 14-4-11) that had been phosphorylated by established auroraArk1 kinase assays35 was used as a substrate. The dephosphorylation reaction was conducted in 20 mM HEPES, 100 mM NaCl, 1 mM EDTA, 0.1% NP40 at 30 °C for 1 h. An established PP2A-B56Par1 activity assay used GST–Rec8391–561 phosphorylated by recombinant GST–Hhp2 (S. pombe casein kinase I) as a substrate22: PP2A-B56Par1.HA was isolated from 2 × 108 cells with 12CA5 antibody (Roche) under non-denaturing conditions. The dephosphorylation reaction was maintained at 30 °C for 1 h, in 50 mM HEPES (pH 7.5), 100 mM NaCl, 0.1% NP40, 2 mM MnCl2, 2 mM dithiothreitol (DTT). 32P levels in the substrate were determined with a Phosphoimager (BioRad). Phosphatase activity was calculated as the reduction of 32P incorporation per unit B56Par1 normalized to the activity of a standard sample from an asynchronous culture. PP2A-B55Pab1 activity was monitored by detecting phosphorylation on S377 and S378 of B56Par1with the phospho-specific antibody B56-Phos (Extended Data Fig. 7a). B56Par1.HA substrate from 6 × 108 mitotically arrested cells36 was independently isolated for three individual assays (scaling up substrate production failed owing to substrate dephosphorylation and degradation during preparation). To isolate this B56Par1.HA devoid of other subunits for PP2A-B55Pab1 assays (Extended Data Fig. 7b, right), a TCA precipitation of total protein was resuspended in a denaturing IP buffer (50 mM HEPES (pH 7.5), 50 mM NaF, 0.4 mM Na3VO4, 40 mM Na–β-glycerol phosphate, 1 mM EDTA, 200 mM NaCl, 0.2% NP40, 1 mM PMSF). The dephosphorylation assay was performed at 30 °C for 1 h (100 mM Tris (pH 7.5), 150 mM NaCl, 10 mM DTT). PP2A and PP1 enzymes were purified by established sequential TAP and immuno-affinity/peptide elution methods37. For TAP tagging, Econo-Pac Chromatography Columns (Biorad 9704652) were packed with IgG Sepharose 6 FastFlow (GE Healthcare 170969-01) or Calmodulin Affinity Resin (Agilent Technologies 214303-52). The following were used for subsequent affinity isolation: anti-HA Affinity Matrix (Roche 101677100), HA peptide (Sigma, I2149), Dynabeads A (Life Technologies) pre-loaded with PP1Dis2 antibodies20 according to manufacturer’s instructions, and the SQNWHMTPPRKNK peptide for elution of PP1Dis2 from the PP1Dis2 polyclonal antibody. Coomassie (Brilliant Blue, Sigma, B2025-1EA) stained 4–12% SDS–NUPAGE gradient gels (Life Technologies, NP20327). Size-exclusion chromatography was performed on a Superdex 200 PC 3.2/30 column (VWR) using buffer B (50 mM HEPES (pH 7.5), 100 mM NaCl, 0.1% NP-40) collecting 50 µl fractions at a flow rate of 50 µl ml−1(ref. 38). All other S. pombe techniques were performed as described previously20.
For immunoprecipitation of human complexes, tetracycline-inducible HeLa cell lines were generated using a pcDNA5/FRT/TO vector (Invitrogen). Cells were grown in advanced DMEM (Invitrogen) supplemented with 2% fetal bovine serum and penicillin/streptavidin. Cells were blocked with thymidine (Sigma-Aldrich) for 24 h and released into fresh medium with 300 nM nocodazole (Sigma-Aldrich) and 1 µg ml−1 tetracycline (EMD). Prometaphase cells were collected by mitotic shake-off after 12 h. Cells were lysed for 20 min on ice in extraction buffer A (50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 0.2% NP-40, 1 mM EDTA, complete inhibitor cocktail tablet (Roche), 0.2 μM microcystin (LGC-Promotech)) and lysates were cleared by centrifugation for 15 min at 12,000g and 4 °C. Protein complexes were precipitated for 1.5 h at 4 °C with anti-Flag (M2; Sigma-Aldrich) covalently coupled to Dynabeads (Invitrogen), washed in extraction buffer, and bound proteins were eluted with LDS-sample buffer (Invitrogen) before analysis by immunoblotting38. Antibodies against the Flag epitope and PP1 (E-9; Santa Cruz sc-7482) were used at 1;1,000.
References
Wu, J. Q. et al. PP1-mediated dephosphorylation of phosphoproteins at mitotic exit is controlled by inhibitor-1 and PP1 phosphorylation. Nature Cell Biol. 11, 644–651 (2009)
Ohkura, H., Kinoshita, N., Miyatani, S., Toda, T. & Yanagida, M. The fission yeast dis2+ gene required for chromosome disjoining encodes one of 2 putative type-1 protein phosphatases. Cell 57, 997–1007 (1989)
Mochida, S. & Hunt, T. Calcineurin is required to release Xenopus egg extracts from meiotic M phase. Nature 449, 336–340 (2007)
Mochida, S., Ikeo, S., Gannon, J. & Hunt, T. Regulated activity of PP2A–B55δ is crucial for controlling entry into and exit from mitosis in Xenopus egg extracts. EMBO J. 28, 2777–2785 (2009)
Schmitz, M. H. et al. Live-cell imaging RNAi screen identifies PP2A–B55α and importin-β1 as key mitotic exit regulators in human cells. Nature Cell Biol. 12, 886–893. (2010)
Qian, J., Winkler, C. & Bollen, M. 4D-networking by mitotic phosphatases. Curr. Opin. Cell Biol. 25, 697–703 (2013)
Dohadwala, M. et al. Phosphorylation and inactivation of protein phosphatase 1 by cyclin-dependent kinases. Proc. Natl Acad. Sci. USA 91, 6408–6412 (1994)
Kwon, Y. G., Lee, S. Y., Choi, Y., Greengard, P. & Nairn, A. C. Cell cycle-dependent phosphorylation of mammalian protein phosphatase 1 by cdc2 kinase. Proc. Natl Acad. Sci. USA 94, 2168–2173 (1997)
Yamano, H., Ishii, K. & Yanagida, M. Phosphorylation of Dis2 protein phosphatase at the C-terminal Cdc2 consensus and its potential role in cell-cycle regulation. EMBO J. 13, 5310–5318 (1994)
Ishii, K., Kumada, K., Toda, T. & Yanagida, M. Requirement for PP1 phosphatase and 20S cyclosome/APC for the onset of anaphase is lessened by the dosage increase of a novel gene sds23+ . EMBO J. 15, 6629–6640 (1996)
Bollen, M., Peti, W., Ragusa, M. J. & Beullens, M. The extended PP1 toolkit: designed to create specificity. Trends Biochem. Sci. 35, 450–458 (2010)
Heroes, E. et al. The PP1 binding code: a molecular-lego strategy that governs specificity. FEBS J. 280, 584–595 (2013)
Kinoshita, N., Ohkura, H. & Yanagida, M. Distinct, essential roles of type-1 and type-2a protein phosphatases in the control of the fission yeast-cell division cycle. Cell 63, 405–415 (1990)
Sents, W., Ivanova, E., Lambrecht, C., Haesen, D. & Janssens, V. The biogenesis of active protein phosphatase 2A holoenzymes: a tightly regulated process creating phosphatase specificity. FEBS J. 280, 644–661 (2013)
Kitajima, T. S. et al. Shugoshin collaborates with protein phosphatase 2A to protect cohesin. Nature 441, 46–52 (2006)
Suijkerbuijk, S. J., Vleugel, M., Teixeira, A. & Kops, G. J. Integration of kinase and phosphatase activities by BUBR1 ensures formation of stable kinetochore-microtubule attachments. Dev. Cell 23, 745–755 (2012)
Kinoshita, K. et al. The regulatory subunits of fission yeast protein phosphatase 2A (PP2A) affect cell morphogenesis, cell-wall synthesis and cytokinesis. Genes Cells 1, 29–45 (1996)
Jiang, W. & Hallberg, R. L. Isolation and characterization of par1+ and par2+ : two Schizosaccharomyces pombe genes encoding B′ subunits of protein phosphatase 2A. Genetics 154, 1025–1038 (2000)
Alvarez-Tabarés, I., Grallert, A., Ortiz, J. M. & Hagan, I. M. Schizosaccharomyces pombe protein phosphatase 1 in mitosis, endocytosis and a partnership with Wsh3/Tea4 to control polarised growth. J. Cell Sci. 120, 3589–3601 (2007)
Grallert, A. et al. Removal of centrosomal PP1 by NIMA kinase unlocks the MPF feedback loop to promote mitotic commitment in S. pombe . Curr. Biol. 23, 213–222 (2013)
King, S. M. & Hyams, J. S. Synchronization of mitosis in a cell-division cycle mutant of Schizosaccharomyces pombe released from temperature arrest. Can. J. Microbiol. 28, 261–264 (1982)
Ishiguro, T., Tanaka, K., Sakuno, T. & Watanabe, Y. Shugoshin–PP2A counteracts casein-kinase-1-dependent cleavage of Rec8 by separase. Nature Cell Biol. 12, 500–506 (2010)
Johnson, L. N. Substrates of mitotic kinases. Sci. Signal. 4, pe31 (2011)
Janssens, V., Longin, S. & Goris, J. PP2A holoenzyme assembly: in cauda venenum (the sting is in the tail). Trends Biochem. Sci. 33, 113–121 (2008)
Porter, I. M., Schleicher, K., Porter, M. & Swedlow, J. R. Bod1 regulates protein phosphatase 2A at mitotic kinetochores. Nat. Commun. 4, 2677 (2013)
Pines, J. & Hagan, I. The renaissance or the cuckoo clock. Phil. Trans. R. Soc. Lond. B 366, 3625–3634 (2011)
Kim, Y., Holland, A. J., Lan, W. & Cleveland, D. W. Aurora kinases and protein phosphatase 1 mediate chromosome congression through regulation of CENP-E. Cell 142, 444–455 (2010)
Xu, Y. et al. Structure of the protein phosphatase 2A holoenzyme. Cell 127, 1239–1251 (2006)
Cho, U. S. & Xu, W. Crystal structure of a protein phosphatase 2A heterotrimeric holoenzyme. Nature 445, 53–57 (2007)
Xu, Y., Chen, Y., Zhang, P., Jeffrey, P. D. & Shi, Y. Structure of a protein phosphatase 2A holoenzyme: insights into B55-mediated Tau dephosphorylation. Mol. Cell 31, 873–885 (2008)
Moreno, S., Klar, A. & Nurse, P. Molecular genetic analysis of fission yeast Schizosaccharomyces pombe . Methods Enzymol. 194, 795–823 (1991)
Fennessy, D. et al. Extending the Schizosaccharomyces pombe molecular genetic toolbox. PLoS ONE 9, e97683 (2014)
Creanor, J. & Mitchison, J. M. Reduction of perturbations in leucine incorporation in synchronous cultures of Schizosaccharomyces pombe made by elutriation. J. Gen. Microbiol. 112, 385–388 (1979)
Qian, J., Lesage, B., Beullens, M., Van Eynde, A. & Bollen, M. PP1/Repo-man dephosphorylates mitotic histone H3 at T3 and regulates chromosomal aurora B targeting. Curr. Biol. 21, 766–773 (2011)
Petersen, J., Paris, J., Willer, M., Philippe, M. & Hagan, I. M. The S. pombe aurora related kinase Ark1 associates with mitotic structures in a stage dependent manner and is required for chromosome segregation. J. Cell Sci. 114, 4371–4384 (2001)
Hagan, I. & Yanagida, M. Novel potential mitotic motor protein encoded by the fission yeast cut7+ gene. Nature 347, 563–566 (1990)
Gould, K. L., Ren, L., Feoktistova, A. S., Jennings, J. L. & Link, A. J. Tandem affinity purification and identification of protein complex components. Methods 33, 239–244 (2004)
Mansfeld, J., Collin, P., Collins, M. O., Choudhary, J. S. & Pines, J. APC15 drives the turnover of MCC-CDC20 to make the spindle assembly checkpoint responsive to kinetochore attachment. Nature Cell Biol. 13, 1234–1243 (2011)
Beullens, M. et al. Molecular determinants of nuclear protein phosphatase-1 regulation by NIPP-1. J. Biol. Chem. 274, 14053–14061 (1999)
Gordon, C., McGurk, G., Wallace, M. & Hastie, N. D. A. Conditional-lethal mutant in the fission yeast 26-S protease subunit Mts3+ is defective in metaphase to anaphase transition. J. Biol. Chem. 271, 5704–5711 (1996)
Gambus, A. et al. GINS maintains association of Cdc45 with MCM in replisome progression complexes at eukaryotic DNA replication forks. Nature Cell Biol. 8, 358–366 (2006)
Hachet, O. et al. A phosphorylation cycle shapes gradients of the DYRK family kinase Pom1 at the plasma membrane. Cell 145, 1116–1128 (2011)
Nurse, P. Universal control mechanism regulating onset of M-phase. Nature 344, 503–508 (1990)
Hudson, J. D., Feilotter, H. & Young, P. G. stf1: non-wee mutations epistatic to cdc25 in the fission yeast Schizosaccharomyces pombe . Genetics 126, 309–315 (1990)
Ishihama, Y. et al. Exponentially modified protein abundance index (emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein. Mol. Cell Proteomics 4, 1265–1272 (2005)
Ohkura, H. & Yanagida, M. S. pombe gene sds22+ essential for a midmitotic transition encodes a leucine-rich repeat protein that positively modulates protein phosphatase-1. Cell 64, 149–157 (1991)
Stone, E. M., Yamano, H., Kinoshita, N. & Yanagida, M. Mitotic regulation of protein phosphatases by the fission yeast Sds22 protein. Curr. Biol. 3, 13–26 (1993)
Acknowledgements
This work was supported by Cancer Research UK (CRUK) grant numbers C147/A16406 and C29/A13678. We thank I. Alvarez-Tabares, K. Labib, K. Gull, M. Yanagida, V. Simanis, J. Bähler and K. Gould for reagents.
Author information
Authors and Affiliations
Contributions
E.B. carried out the initial identification, validation and timing of PP1Dis2–B55Pab1/B56Par1association, initial two-hybrid and co-immunoprecipitation assays, initial determination of specificity and timing of PP1Dis2.T316 phospho-recognition, and phospho-mapping of B56Par1 (with D.L.S. and Y.C.) and generated some mutant alleles. E.B. generated Fig. 4d; all other data, except for Fig. 4k and Extended Data Figs 5c, 9b–d, were generated and largely devised by A.G. Data for Extended Data Fig. 5c were generated by A.G., Y.C., D.L.S. and J.R.G. Molecular biology and background work for human two-hybrid assays and generation of human vectors was carried out by B.H. I.M.H. devised the work presented in Figs 1c, 2a, c, f and 4d with E.B., and devised the remainder of the work with A.G. A.H. and J.P. designed and executed the human work and gel filtration.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 PP1 phosphorylation by Cdk1–cyclin B on T316 represses activity and reduces abundance in a manner that is mimicked by mutation of the target site to an acidic residue in PP1Dis2.T316D.
a, Clustal W alignments of the C-terminal residues of the indicated PP1 isoforms. The Cdk1–cyclin B phosphorylation site is highlighted by red shading. Red letters within this region of shading highlight the clear deviation from this consensus in the S. pombe PP1Sds21 isoform. PP1Sds21 is unable to bind to either B55Pab1 or B56Par1. The molecular basis for this inability of PP1Sds21 to bind the regulatory subunits of PP2A remains to be established. The initial threonine in the S. pombe PP1Dis2 shaded consensus sequence region is T316. b, c, Phosphatase assays of the indicated samples with activity detection by either EnzChek (b) or a second established PP1 activity assay34 histone H3 serine 10 dephosphorylation (c) . For each assay the graphs show arbitrary units. Error bars show s.d. Identical results were obtained with each assay. The similarity between the recombinant rabbit PP1γ and potato acid phosphatase (PAP) activities indicate that both reactions measure PP1 activity. The inhibition of the activity of PP1Dis2 precipitates by 1 μM, but not 10 nM okadaic acid (OA), suggests that it is the PP1Dis2 activity rather than any co-precipitating PP2A activity that is being monitored in these assays. This view is consolidated by the inhibition of activity in both assays by the PP1-specific inhibitor NIPP1 (2, 5, 10 and 15 nM; c and Fig. 1a). 5 nM is routinely used to inhibit mammalian PP1 (refs 34, 39). Manufacturer’s instructions were followed for EnzChek assays. For histone H3 serine 10 assays, histone H3 that had been phosphorylated by auroraArk1 kinase35 was used as a substrate. The dephosphorylation reaction was conducted in 20 mM HEPES, 100 mM NaCl, 1 mM EDTA, 0.1% NP40 at 30 °C for 1 h. The molarity of NaCl indicates the salt concentration of the buffer used for the immunoprecipitation, not the molarity within the phosphatase reaction. All phosphatase reactions for each substrate were conducted under identical conditions. A 1.2 M NaCl, buffer was used to isolate PP1Dis2 for the experiments in Fig. 3c, d and Extended Data Fig. 6d, g, h. The level of PP1 activity detected in either assay shown in b and c was the same irrespective of the NaCl concentration in the buffer used to isolate PP1Dis2. Phos-Stop is a universal phosphatase inhibitor. The key conclusion from all these assays is that the PP1Dis2.T316D protein has a similar level of activity as the Cdk1–cyclin B phosphorylated enzyme and so behaves as a ‘genetically inhibited’ phosphatase in the subsequent experiments described in the manuscript. d, Validation of the T316Phos antibody. PP1Dis2 immunoprecipitates from the indicated strains were treated as indicated and probed with either PP1Dis2 or T316Phos polyclonal antibodies. e–g, Cultures synchronized with respect to cell cycle progression by size selection through elutrient centrifugation were probed with the PP1Dis2 antibodies at the high dilution that revealed the fluctuations in PP1Dis2 levels in Fig. 1c. Graphs show the septation index alongside the PP1Dis2 levels in each sample normalized to the level of the α-tubulin loading control and a sample from an asynchronous culture that was run on each gel. e, Samples from this Rpn12mts3.1 culture were split into three immediately after elution and one-third was maintained at the permissive temperature of 25 °C for the remainder of the experiment, another third was immediately shifted to 36 °C to inactivate this essential component of the proteasome 26S subunit40, while the inactivating temperature shift of the remaining third was done 200 min later, after cells had completed one round of division at 25 °C. Note that the mitotic decline in PP1Dis2 levels at the permissive temperature as cells progress from mitotic commitment to the metaphase anaphase transition is not seen at 36 °C when the proteasome function is inactivated, despite the fact that the chromosome condensation index indicates that over 60% of cells have arrested cell cycle progression with a metaphase spindle. f, g, PP1Dis2 levels fail to oscillate when the Cdk1–cyclin B phosphorylation site at position T316 is mutated. A switch to the phospho-mimetic glutamic acid results in persistently low protein levels (f) while levels are persistently high upon mutation to alanine to block phosphorylation (g). Because these levels remain constant throughout the cell cycle these normalized levels have been superimposed upon the data from a wild-type culture in Fig. 1c to show each level relative to the oscillating wild-type protein levels. The septation profile in Fig. 1c is the profile for that wild-type culture. The septation profile in Fig. 1c does not contain any data from either of the cultures shown here in Extended Data Fig. 1f, g. The septation profiles for each mutant culture are shown in f and g. Biological replicates: for b and c, n = 3; for d, n = 2; for e–g, n = 1.
Extended Data Figure 2 Association of PP1Dis2 with the PP2A regulatory subunits B56Par1 and B55Pab1.
a–d, Immunoprecipitation reactions in which PP1Dis2 polyclonal antibodies precipitated and detected PP1Dis2, green fluorescent protein (GFP) antibodies precipitated and detected either PP1Dis2 or PP1Sds21 functional fusion proteins19 or 12CA5 monoclonal antibodies precipitated HA-tagged B55Pab1 and B56Par1 regulatory subunits of the PP2A phosphatase, as indicated. These blots establish that the association between PP1Dis2 and both B56Par1 and B55Pab1 was independent of PP1Sds21 function. No association was detected between PP1Sds21 and any PP2A regulatory subunit (including B56Par2 (data not shown)). b, c, Red asterisks indicate a non-specific band that was detected by the anti-GFP antibodies. d, No association was detected between B56Par2 and PP1Dis2, even when cell numbers in the precipitates were increased tenfold (numbers shown in red under the panel are x × 108 cells per ml) to enhance the sensitivity of detection and the B56par1+ gene encoding the B56Par1 subunit was deleted in an attempt to remove any competition from this primary B56 isoform. e, Expression of B56par2.PkC and B56par2. L482VPkC genes from the pINT41PkC integration vector32 was de-repressed by removal of thiamine. B56Par2 proteins (top) or PP1Dis2 (bottom) were precipitated with antibodies against the Pk epitope (top) or PP1Dis2 peptide (bottom), respectively, and these immunoprecipitation reactions were run alongside aliquots of the equivalent whole-cell extracts (WCE), before western blotting detected PP1Dis2 and B56Par2 fusion proteins. In both assays, the PP1-docking-site mutant in which the leucine of B56Par2 was replaced with the valine of B56Par1 (B56Par2.L482V) associated with PP1 whereas the wild-type B56Par2 protein did not. Biological replicates: for a, n = 6; for other panels, n = 2.
Extended Data Figure 3 Yeast two-hybrid assays reveal docking-site-dependent association between B56Par1 and PP1Dis2.
a, Yeast two-hybrid assays in which PP1dis2+ and PP1sds21+ sequences were fused to the activating domain of Gal4 and the indicated version of the core homology domains of the PP2A regulatory subunits B56par1+ (encoding amino acids 112–424) and B56par2+ (encoding amino acids 213–525) were fused to the Gal4 DNA-binding domain according to the procedures of the matchmaker gold yeast two-hybrid system. b, Blotting the cell extracts with 12CA5 and 9E10 monoclonal antibodies that recognized HA and Myc epitopes within the cassettes harbouring the activation (HA) and DNA-binding domains (Myc) indicated that equivalent protein expression levels were achieved for each version of the protein. Probing for the DNA replication factor Ctf4 was used as a loading control41. Thus, the failure of the PP1-docking-site mutants to interact with PP1dis2+ suggests that the change in the PP1-docking site abolished the affinity between the two molecules. Biological replicates: for a, n = 3; for b, n = 1.
Extended Data Figure 4 Validation of the SKEVLF and GLLR motifs of the PP1-docking consensus site of B56Par1 in the polarity protein Wsh3 and the mitotic regulator Cut12.
a–d, The morphology protein Wsh3 is required to recruit PP1Dis2 to cell tips19. Wsh3-recruited PP1 promotes the dephosphorylation of the DYRK family kinase Pom1 (ref. 42). Dephosphorylation of Pom1 promotes its association with the cell cortex. Subsequent auto-phosphorylation diminishes this newfound affinity for the cortex until the kinase loses its affinity for the cortex. Because Wsh3 is only found at cell tips, Pom1 only associates with the cortex at the tips42. We exploited this relationship to test the ability of the SKEVLF sequence in the PP1-docking consensus site of B56Par1 to function as a PP1Dis2-docking site in vivo. a, The indicated wsh3 alleles were cloned into the pINTL41PkN vector32 and integrated at the leu1 locus before introduction into wsh3.Δ background in which the endogenous wsh3+ gene had been deleted. b, Expression of the transgenes was repressed by the addition of thiamine to culture medium (uninduced) before filtration into thiamine-free medium induced expression of each allele 24 h later (induced). c, PP1Dis2.GFP was not enriched at cell tips in the absence of wsh3 induction. Induction of the wild-type or wsh3.I232SR234ET236L allele in which the SKEVLF motif of the B56Par1 PP1Dis2-docking site was substituted for the native IFRVTF motif of Wsh3 led to the recruitment of PP1Dis2.GFP to cell tips whereas expression of the PP1-docking-site mutant wsh3.F237A failed to do so. d, Pom1 recruitment to cell tips upon expression of both the wild-type and SKEVLF alleles, but not the PP1-docking-site alleles, indicated that the PP1Dis2 recruited to Wsh3 by the SKEVLF allele was functionally indistinguishable from that recruited by the wild-type molecule. e–g, Commitment to mitosis is promoted by the activation of Cdk–cyclin B. Cdk1–cyclin B activity is regulated by the level of phosphorylation on threonine 14 and tyrosine 15 within its ATP-binding site. Wee1 kinase phosphorylates these residues while Cdc25 removes the phosphate to trigger mitotic commitment43. The lack of Cdc25 phosphatase activity achieved by incubating cells harbouring the temperature-sensitive cdc25.22 mutation at temperatures above 34 °C kills cells because they are unable to remove the inhibitory phosphate from Cdk1. The cut12.s11 and cut12.s14 mutations enable cdc25.22 cells to divide at 34 °C (refs 20, 44). cut12.s11 is a G71V mutation at the start of a highly conserved bipartite PP1-docking site GILKTPGTLQIKKTVNF (the letters in bold indicate the residues that constitute the PP1-docking site). cut12.s14 is a F87L mutation at the end of the same docking motif20. e, Alignment of the conserved PP1-docking site of Cut12 homologues from the indicated fungi and the mutations generated in each of the indicated cut12 alleles. f, g, Mutation of the PP1-docking site of S. pombe Cut12 to abolish the function of either the GILK or the KxVxF motifs of Cut12 reduces PP1Dis2 association in immunoprecipitation assays (f) and suppresses the lethal mitotic commitment defect of cdc25.22 (g). Simultaneous mutation of both motifs abolishes PP1 association in immunoprecipitation assays and confers a higher level of suppression of cdc25.22 (g) that can enable cells to survive complete removal of cdc25+ coding sequences20. We asked whether the GLLR sequence found upstream of the B56Par1 KxVxF motif (Fig. 2a) would substitute for the GILK sequence of Cut12 in both the co-immunoprecipitation and cdc25.22 suppression assays; in both cases we found that it could. The association between PP1Dis2 and Cut12 in the cut12.I72LK74R mutant that converts the GILK motif to a GLLR was indistinguishable from wild type. Also, the G71V mutation of cut12.s11 suppressed cdc25.22 whereas cut12.I72LK74R did not (g). Thus, the function bestowed on Cut12 by the GILK sequence is fully maintained upon substitution with GLLR, indicating that GLLR is a functional variant of GILK. Error bars show s.d. Biological replicates: for b–d, n = 1; for f, n = 5; for g, n = 3.
Extended Data Figure 5 In vitro association of purified PP1Dis2 with PP2A-B55Pab1 and PP2A-B56Par1 complexes in a docking-site-dependent manner.
a–g, The tandem affinity purification steps used in Extended Data Fig. 10e were followed by immuno-affinity with antibodies directed against the HA epitope on B55Pab1.HA, B56Par1.HA fusion proteins or the unique sequence SQNWHMTPPRKNK in the C terminus of PP1Dis2(ref. 20) using anti-HA Affinity Matrix followed by elution with HA peptide, or Dynabeads A pre-loaded with PP1Dis2 antibodies20 followed by elution with the SQNWHMTPPRKNK peptide, respectively. b, Coomassie-stained 4–12% SDS–NUPAGE gradient gels. In each case the first two sample lanes show the purified wild-type and PDSN PP2A holoenzymes and the fifth lane shows the purified PP1Dis2 enzyme. The quantity of PP1Dis2 in the fifth lane was mixed with the quantity of each PP2A complex shown in lanes 1 and 2 before the PP1Dis2 was re-isolated from the mixture with beads that bore PP1Dis2 antibodies. This re-isolated PP1Dis2 (and any associated partner molecules) was then eluted from these beads with the SQNWHMTPPRKNK peptide and the eluted proteins run in lanes 3 and 4 of each gel. For both the B55Pab1 (left) and B56Par1 (right) PP2A holoenzymes, the wild type but not the PDSN complex bound to PP1Dis2. The numbered arrows indicate the lanes from which bands were excised for protein identification by mass spectrometry in c. c, Protein identification by mass spectrometry of the indicated bands from the lanes on the SDS–PAGE gels highlighted by the numbered arrows in b. emPAI scores45 show that the purified PP1Dis2 preparations used for the in vitro reconstitution reaction and run in lane 5 of the two gels in b also contained the conserved and well-characterized PP1 partner Sds22 (refs 46, 47). emPAI scores for the bands found in the reconstituted PP2A holoenzyme–PP1 complexes show that the predominant molecule identified in each complex corresponds with a known subunit of the PP2A complex, PP1Dis2 or Sds22. d, Western blotting to detect the indicated components in the complexes used for b and c. The epitope-tagged regulatory B55Pab1 and B56Par1 molecules were detected with antibodies against the HA tag, while the catalytic CPpa1 and CPpa2 and scaffolding APaa1 subunits of PP2A were detected with commercial antibodies. e, Western blots of gel filtration chromatography fractions of either the isolated PP2A holoenzyme complexes (top two blots) or the eluted PP2A holoenzyme/PP1 quaternary complexes (bottom two blots) corresponding to lanes 1 and 3 of the two SDS–NUPAGE gels of b. Note the very low level of PP2A-B55Pab1/PP1Dis2 and PP2A-B56Par1/PP1Dis2 quaternary complexes in the isolated PP2A holoenzyme preparations in the top two blots. Migration of the different complexes at the appropriate size suggests that the procedures used isolated correctly folded proteins. f, Phosphatase assays of the isolated enzymes used in b–e show that these molecules are active phosphatases and so the procedures used isolated functional, correctly folded complexes rather than denatured, inappropriately folded, proteins. Error bars show s.d. g, B56-Phos blots of the TAP- and HA-purified holoenzyme complex preparations used in c alongside a similarly purified PP2A-B56Par1.S378A holoenzyme complex. Biological replicates: for b, n = 3; for c and e, n = 1; d and g, n = 2; for f, n = 5.
Extended Data Figure 6 Mitotic enhancement of PP2A-B56 activity relies upon recruitment of active PP1Dis2 and is required for accurate chromosome segregation.
a, DAPI staining of the Cdk1cdc2.33 strains in which mitotic progression has been synchronized by transient arrest at the restrictive temperature of 36 °C. While two evenly sized chromatin masses are generated by the anaphase in wild-type cells, chromosome segregation is uneven in the mutant lines and lagging chromosomes are frequently observed. The frequency of phenotypes is presented in Fig. 2e. b, The PP2A-B56Par1 phosphatase assay was carried out as previously described, in which recombinant GST–Rec8391–561 was phosphorylated by a fusion between glutathione and S. pombe casein kinase I (Hhp2) that had also been produced in Escherichia coli22. PP2A-B56Par1.HA was isolated from 2 × 108 cells with 12CA5 antibody under non-denaturing conditions. Phosphatase activity was calculated from the reduction of 32P incorporation in the GST–Rec8391–561 substrate per unit B56Par1.HA. The level obtained in this assay (and every assay presented in this study) was normalized to the reduction in substrate phosphorylation displayed by a B56Par1 precipitate from 2 × 108 cells of an asynchronous B56par1.HA culture run on the same gel (the second lane in b). OA, okadaic acid. c, PP1Dis2 activity assays conducted in parallel with the same samples used in b. These assays established that both the recombinant rabbit PP1γ and the PP1Dis2 samples that were added to the PP2A-B56Par1 assay in a contained PP1 phosphatase activity. We conclude that we have successfully re-established the phosphatase assays described previously22 and that the addition of PP1 to this assay did not alter the phosphorylation status of the phosphorylated GST–Rec8 substrate, indicating that PP1Dis2 displays no activity towards phosphorylated GST–Rec8 used in this PP2A-B56Par1 enzyme assay. d, This panel presents blots of protein levels (left), PP1Dis2 activity assays (centre) and PP2A-B56Par1 phosphatase assays (right) of the PP1Dis2 samples used in the add back experiments in g (bottom blot) and h (bottom three blots). The blot of PP1Dis2 levels on the left shows that similar levels of the different PP1Dis2 proteins were added in each case, while the central panel shows that these samples possessed PP1Dis2 activity. The panel on the right shows that none of the PP1Dis2 samples exhibited any activity in the PP2A-B56Par1 phosphatase assay. In other words, there was no PP2A-B56Par1 in these pull downs despite the fact that PP1Dis2 is able to bind to PP2A-B56Par1. The absence of PP2A-B56Par1 from these samples is either due to the fact that only a minor fraction of the PP1Dis2 complex formed a complex with PP2A-B56Par1, or, as we anticipate, that the high salt (1.2 M NaCl) conditions we used in the immunopreciptation reactions that isolated these PP1Dis2 molecules for the add back experiments (g, h and Fig. 3c, d) had disassociated any PP2A-B56Par1 molecules that partnered these PP1Dis2 molecules in vivo. e–k, PP2A-B56Par1 assays as for b. A quantitative plot of the data in g (top blot) is shown in Fig. 3b while the data from g (bottom blot) and h are represented in the plots in Fig. 3c, d and data from i–k are shown in Fig. 3g–i. For the experiments in g (middle and bottom blots) and h we exploit the redundancy between PP1Dis2 and PP1Sds21 to use PP1Sds21 to provide essential PP1 function and support the viability of PP1dis2.Δ cells2. The PP2A-B56Par1 complexes isolated from these cells have therefore never been exposed to PP1Dis2 regulation and so will be fully phosphorylated on PP1Dis2-target sites. The addition of PP1Dis2 in vitro enables us to assess the impact of dephosphorylation of these sites. To ensure that there could be no cell cycle dependency to any outcome in these assays we assessed the impact of PP1Dis2 addition on PP2A-B56Par1 samples from all stages of a Cdk1cdc2.33 synchronized mitosis. The data from these experiments that are presented in Fig. 3 and in d, g and h clearly demonstrate that the addition of active PP1Dis2 to naive PP2A-B56Par1 complexes that have never been exposed to PP1Dis2 in vivo was able to reactivate the PP2A activity as long as the PP1Dis2-docking site within B56Par1 was intact. In contrast, PP1Dis2 addition failed to reactivate these naive PP2A-B56Par1 complexes when the PP1Dis2-docking site in B56Par1 had been mutated to block PP1Dis2 recruitment, or the Cdk1–cyclin B inhibition site at T316 of PP1Dis2 had been mutated to aspartic acid to mimic the phosphorylated state. Biological replicates: for b, n = 3; for c and d, n = 1; for all other panels, n = 2.
Extended Data Figure 7 Residue 378 within the PP1-docking site of B56Par1 was dephosphorylated by PP2A-B55Pab1 that had been activated by PP1Dis2 recruitment to its own PP1-docking site.
a, Validation of the B56-Phos antibody that was raised against and affinity purified with a peptide in which residues corresponding to S377 and S378 were each phosphorylated. Immunoprecipitates in which the 12CA5 monoclonal antibody was used to recognize the HA epitope on a B56Par1.HA fusion protein or commercial rabbit antibodies raised against human B56ε were used to precipitate B56Par1 and probed as indicated. b, Validation of the PP2A-B55Pab1 activity assay. This assay exploits the fact that PP2A-B55Pab1 removes B56-Phos reactivity from B56Par1.HA. The phosphorylated B56Par1.HA substrate was precipitated from cells in which mitotic progression had been arrested by incubation of Eg5cut7.24 cells at the restrictive temperature for 3 h. Eg5cut7.24 is a temperature-sensitive mutation in the kinesin 5 motor protein that is required for interdigitation of the two halves of the bipolar spindle36. To ensure that these B56Par1 precipitates were free of any B56Par1 partners, the total protein content of intact cells was precipitated by trichloroacetic acid (TCA) treatment before cells were homogenized and resuspended in 2% SDS. After a centrifugation clearing step, the addition of 1% Triton X-100 to the resultant supernatant sequestered the SDS into micelles thereby generating conditions for immunoprecipitation (IP) of phosphorylated B56Par1.HA substrate. Lane 2 on the right shows that that this procedure disassociated the B56Par1 subunit from the other subunits of the tripartite PP2A-B56 complex whereas non-denaturing immunoprecipitation conditions (lane 1) do not. The CPpa1 and CPpa2 catalytic subunits are detected with a commercial antibody. The B56Par1 samples generated in this way were incubated with the indicated components. ‘Eluted’ indicates reactions in which the B55Pab1 immunoprecipitate was incubated with a peptide corresponding to the PP1Dis2-docking site on B56Par1 (CWPKVNSSKEVLF) to disassociate the PP1Dis2 enzyme from the PP2A-B55Pab1 complex before a re-purification isolated the PP1Dis2 complex once more. It was this re-isolated PP1Dis2 that was used for the assay in lane 9 while the residual PP2A-B55Pab1 was used in the assay in lane 6. The impact of this disassociation can be seen by comparing lanes 8 and 9. The PP1Dis2 immunoprecipitate exhibited a low level of B56-Phos phosphatase activity when the elution step was not performed. Displacement of partner molecules through peptide incubation removed the activity that reduced B56-Phos reactivity in the uneluted PP1Dis2 precipitates. The inhibitor profiles all support the conclusion that this assay specifically detected PP2A-B55Pab1 activity. Phos-Stop is a commercial pan-phosphatase inhibitor. c–f, PP2A-B55Pab1 activity assays of cultures in which mitotic progression had been synchronized by the Cdk1cdc2.33 arrest/release protocol. The quantification of these assays is presented in Fig. 4f–i. Biological replicates: n = 2 for all panels except the right panel of b, where n = 1.
Extended Data Figure 8 Mitotic control of phosphatase activities.
A schematic representation of the control and activity levels of the three phosphatase activities. This view is derived from bulk activity assays. We believe that localized reactions at particular times at discrete locations will differ; however the basic relationships between the three phosphatases outlined here will apply. Left, as previously established in a number of studies1,7,8,9,10, PP1Dis2 activity is repressed by Cdk1–cyclin B phosphorylation upon mitotic commitment through phosphorylation on T316 at a rate that exceeds the rate at which auto-dephosphorylation reactivates inhibited PP1. PP1Dis2T316 phosphorylation also targets PP1Dis2 for destruction to further reduce PP1Dis2 activity, although the majority of the protein persists. It is currently unclear whether this reflects a highly localized destruction of a particular pool of PP1Dis2 in its entirety or a rate of destruction across the population that is simply too slow to outcompete synthesis. Cyclin B destruction curtails the inhibitory phosphorylation on T316, enabling PP1Dis2 to auto-reactivate and persist in an active state for the remainder of mitosis. Middle, PP2A-B55Pab1 activity is inhibited upon mitotic commitment (Extended Data Fig. 9a). The mechanism by which this inhibition is achieved is currently unclear. The inactivated PP2A-B55Pab1 recruits T316-phosphorylated (and therefore catalytically compromised) PP1Dis2 upon mitotic commitment. Cyclin B destruction upon the metaphase/anaphase transition promotes PP1Dis2 reactivation (left). The reactivation of the PP2A-B55Pab1-associated PP1Dis2 enables this PP1Dis2 to reactivate PP2A-B55Pab1. PP1Dis2 then disassociates from B55Pab1. The molecular basis for the staged association between PP1Dis2 and B55Pab1 upon mitotic commitment and dissociation at the metaphase/anaphase transition remains to be established. PP2A-B55Pab1 persists in an active state for the remainder of mitosis. Right, PP2A-B56Par1 activity is inhibited upon mitotic commitment (Extended Data Fig. 9a). The mechanism by which this inhibition is achieved is currently unclear. PoloPlo1 activation upon mitotic commitment enables it to phosphorylate S378 within the PP1-docking site on B56Par1 to block PP1Dis2 recruitment to this B′ regulatory subunit of the PP2A-B56 holoenzyme. This phosphorylation persists to block PP1Dis2 recruitment until telophase when the balance between PoloPlo1 kinase and PP2A-B55Pab1 phosphatase activities tips in favour of PP2A-B55Pab1 to remove phosphate from S378, thereby promoting the recruitment of active, T316-dephosphorylated PP1Dis2. PP1Dis2 recruitment enables this phosphatase to reactivate the PP2A-B56Par1 holoenzyme.
Extended Data Figure 9 PP1Dis2, PP2A-B55Pab1 and PP2A-B56Par1 activities decline as cells arrest cell cycle progression in mitosis, and the association between human PP1 and PP2A-B56 isoforms.
a, The temperature of Eg5cut7.24 B55pab1.HA, Eg5cut7.24 B56par1.HA and Eg5cut7.24 cultures that had been grown overnight to early log phase in supplemented EMM2 medium at 25 °C was increased to 36 °C at t = 0 to inactivate this kinesin 5 and so arrest cell cycle progression in mitosis36. Samples were taken for the phosphatase assays and to monitor the degree of mitotic arrest by scoring the frequency of cells with condensed chromosomes36. Protein phosphatase assays were conducted as described for Extended Data Figs 1b, 6b and 7b. Each activity declined as there was an increase in the frequency of cells in which the inability to form a bipolar spindle has triggered a mitotic arrest due to activation of the spindle assembly checkpoint. b, Two-hybrid assays with the indicated human PP1 isoforms with wild-type and mutant isoforms of the conserved core domain (sequences encoding amino acids 84–400) of human B56γ. c, The mutations introduced in each case. d, Blotting the cell extracts from the two-hybrid clones shown in b with 12CA5 and 9E10 monoclonal antibodies that recognized HA and Myc epitopes within the cassettes harbouring the Gal4 activation and DNA-binding domains, respectively, indicated that equivalent protein expression levels were achieved for each version of the protein. Thus, the failure of the PP1-docking site mutants indicates that the change in the PP1-docking site abolished the affinity between the two molecules. Biological replicates: for a and d, n = 1; for b, n = 3.
Extended Data Figure 10 PP1-docking-site mutations do not alter the stoichiometry of PP2A subunits in immunoprecipitation assays or of holoenzymes isolated by TAP-tag purification.
a–c, Immunoprecipitation reactions from cell extracts of the indicated strains in which the 12CA5 antibodies were used to precipitate epitope-tagged versions of the B55Pab1 or B56Par1 PP2A regulatory subunits or in which polyclonal antibodies against GFP were used to precipitate a GFP-tagged version of the APaa1 PP2A scaffolding subunit. Although mutation of either the PP1-docking site motif sequences or the B56par1.S378D mutation abolished PP1 precipitation with each PP2A holoenzyme, they had no impact upon the stoichiometry of PP2A subunits precipitating with either the regulatory or the scaffolding subunits. We conclude that the PP1-docking site mutations do not alter the integrity of the PP2A-B55 or PP2A-B56 holoenzyme complexes. The ability to detect S377/S378 phosphorylation in APaa1 precipitates consolidates the data in Extended Data Fig. 5g to demonstrate that this site between the two elements of the PP1-docking site of B56Par1 can be phosphorylated in the PP2A-B56Par1 holoenzyme complexes. Catalytic CPpa1 and CPpa2 and the scaffolding APaa1 subunits of PP2A were detected with commercial antibodies. d, The tandem affinity purification scheme used to isolate PP2A phosphatases from yeast cultures via the TAP tag fused to the scaffolding APaa1 subunit. e, Coomassie-stained 4–12% SDS–NUPAGE gradient gels of APaa1.TAP purifications of the indicated strains. The disappearance of bands upon gene deletion combined with mass spectrometric analysis of isolated bands after a further round of purification (Extended Data Fig. 5b, c) confirms that this procedure isolates PP2A enzymes. The persistence of the wild-type pattern in B55pab1.PDSN, B56par1.PDSN and B56par1.S378A strains indicates that these mutations do not alter the composition of either PP2A holoenzyme, that is, they do not alter the structural bonds that maintain the integrity of the PP2A holoenzyme complexes, as predicted from existing current crystal structures of PP2A holoenzyme complexes28,29,30. Biological replicates: for all panels, n = 2.
Supplementary information
Supplementary Table 1
This file contains the strains used in this study. (PDF 167 kb)
Rights and permissions
About this article

Cite this article
Grallert, A., Boke, E., Hagting, A. et al. A PP1–PP2A phosphatase relay controls mitotic progression. Nature 517, 94–98 (2015). https://doi.org/10.1038/nature14019
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14019
This article is cited by
-
Analog-sensitive Cdk1 as a tool to study mitotic exit: protein phosphatase 1 is required downstream from Cdk1 inactivation in budding yeast
Chromosome Research (2023)
-
Core control principles of the eukaryotic cell cycle
Nature (2022)
-
Evolution of opposing regulatory interactions underlies the emergence of eukaryotic cell cycle checkpoints
Scientific Reports (2021)
-
Protein phosphatase 1 regulates atypical mitotic and meiotic division in Plasmodium sexual stages
Communications Biology (2021)
-
Wip1 controls the translocation of the chromosomal passenger complex to the central spindle for faithful mitotic exit
Cellular and Molecular Life Sciences (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
