
Abstract
Replying to R. C. P. Mounce & M. Wills Nature 476, 10.1038/nature10266 (2011); D. A. Legg et al. Nature 476 10.1038/nature10267 (2011)
We welcome the reanalyses by Mounce and Wills1 and Legg et al.2 of our paper3, and although we do not fully concur with their conclusions we are pleased that Diania has reopened the debate about key stages in arthropod evolution. We accept that the position of this fossil remains sensitive to parameters of analysis and in the original publication we conceded that our best-supported tree—Diania as sister-group to (Schinderhannes + Euarthropoda)—could be subject to change, and that the ‘walking cactus’ may have a more basal position within the overall framework of the arthropod stem-group. These alternative treatments of our data would seem to confirm this suspicion, although we find the placement of Diania in an unresolved, and extremely basal, polytomy alongside velvet worms, tardigrades and various other lobopodians similarly problematical. We do not doubt that the authors’ results1,2 are statistically well supported, but what do these cladograms tell us about the evolution of the group? Lobopodians are, by their nature, fairly simple and consequently yield few convincing synapomorphies, either with each other or with arthropods in general. As we discovered, this makes scoring a robust data matrix including both lobopodians and arthropods challenging, and we wonder whether the basal polytomies recovered here are simply due to clustering among taxa with few unequivocal apomorphies and/or much missing data.
Similar content being viewed by others


Main
Our original placement of Diania close to the euarthropods was strongly influenced by the character of jointed trunk appendages. We acknowledge an error in scoring characters 17 and 27 of Tardigrada, which should have been 0 and 1, not 1 and 0, respectively. We also accept the criticism by Mounce and Wills1 that parent–daughter characters are, to some extent, dependent upon one another and can artificially inflate support for particular clades. This situation is hard to avoid when selecting characters across a range of lobopod/arthropod fossils, which include taxa with unusual morphologies (for example, Opabinia) or where there are alternative hypotheses for the homology of a given lobopod feature and its probable arthropod equivalent: should a lobopod/dinocaridid flap be scored the same way as a euarthropod limb exite; could both be subsumed under a ‘biramous limb’, etc.? Furthermore, a disadvantage of PAUP is that ‘gaps’ are automatically treated as missing in the analysis, and the only way to distinguish between them is to treat the gap as a new state; although a gap in our analysis refers to an inapplicable character, which is itself not a new state.
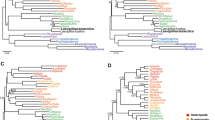
In our initial analysis data were entered into a matrix using MacClade version 3.05 (ref. 4). Analyses were performed with PAUP version 4.0b10 (ref. 5), whereby multi-state characters were treated as ‘unordered’, other characters were treated as ‘ordered’, and a branch and bound search under implied weights (k = 2) produced many most parsimonious trees—including trees similar to those of Legg et al.2 and Mounce and Wills1. For the purpose of character state mapping, we selected one of these most parsimonious trees (see Fig. 4 in ref. 3) which we felt best reflected the complex relationships among lobopodians and arthropods. Also, a constrained analysis was conducted, and three monophyletic groups (lobopodians + Onychophora, Radiodonta, Arthropoda) were enforced. To verify the stability of this tree, we conducted bootstrap analysis and the result (Fig. 1) shows that the contained 26 resolved nodes of this tree were also strongly supported by bootstrap values (100+) and in some nodes bootstrap values showed more resolution. Additionally, a significant value of the partitioning tail permutation (PTP) test (P = 0.01) suggests the presence of a clear phylogenetic signal in the morphological data, also strongly supporting the topology shown. Furthermore, a test of Bremer support still supports our initial analysis (Fig. 2).

Hierarchical structure in the data was assessed using the PTP test6 as implemented in PAUP. Parsimonious trees were found through a heuristic search strategy using tree bisection-reconnection (TBR) branch swapping. In an initial analysis, all characters were treated as unordered. Branch support was based on calculating bootstrap values (10,000 replicates and with rearrangement limit of 10,000,000 rearrangements per additional sequence). A significant value in the PTP test (P = 0.01) does suggest the presence of a phylogenetic signal in the morphological data, supporting the given topology. Thirty-eight characters were parsimony-informative. The heuristic search for a maximum parsimony solution resulted in only one parsimonious tree (length = 121, homoplasy index (HI) = 0.67, consistency index (CI) = 0.33, retention index (RI) = 0.55, excluding 1 uninformative character). This tree contains 26 resolved nodes, of which many were strongly supported by bootstrap values (100+) and in some nodes bootstrap values showed even greater resolution.

Four additional steps would be required to disrupt the Diania–arthropod relationship. As we noted in the original description, Diania is the most arthropod-like of the lobopodians recorded so far—at least in terms of limb morphology—and we feel that it is better placed on the arthropod stem, rather than in an unresolved polytomy.
Legg et al.2 drew attention to the absence of sclerotized trunk limbs in dinocaridids (Anomalocaris, etc.), which remains for us one of the great puzzles of stem-group arthropod evolution. Put simply, dinocaridids have a more arthropod-like head region (cephalisation, eyes, sclerotized mouthparts), whereas Diania lacks such sophistication in the anterior body region, but has jointed trunk appendages. One solution to this puzzle would be to assume that dinocaridids also had trunk limbs, but lost them secondarily. In this scenario Diania could sit comfortably as sister group to a (Dinocaridida + Arthropoda) clade: all three sharing jointed appendages, the latter two sharing cephalisation, etc.
We accept that the tree of Legg et al.2 and Mounce and Wills1 is the strict consensus tree, but we feel that the strict tree is, in this case, too conservative to provide meaningful information about the early evolution of the arthropods. This was undoubtedly a complex process, and may have involved numerous parallel developments introducing homoplasy. Under these circumstances, the comb-like most parsimonious tree is unhelpful and exemplifies a disadvantage of parsimony in this instance when faced with a complex early radiation. Indeed, Legg et al.2 recognized that the strict consensus tree(s) “suggests a polyphyletic origin of arthropodized limbs”. If true, this would be significant. In this context, Mounce and Wills1 seem to have overlooked the potential significance of their reanalysis of our data. Much evidence has been accumulated that Arthropoda is monophyletic, but the logical conclusion of both reanalyses is that jointed legs are homoplastic, at least within the lobopodian–arthropod assemblage. The implications of this are not trivial, bringing us close to the (largely discredited) Manton school of thought in which jointed appendages were proposed to have evolved in parallel in different (here euarthropod) lineages. At a fundamental level we need to know how easy it is to turn a soft, lobopodian limb into a sclerotized arthropod one, and whether this happened once, twice or on multiple occasions. For this reason alone, we believe that Diania is not merely fascinating, but remains invaluable to the evolutionary debate by challenging our notion of what it means to be an arthropod.
References
Mounce, R. C. P. & Wills, M. Phylogeny of Diania challenged. Nature 476 10.1038/nature10266 (2011)
Legg, D. A. et al. Lobopodian phylogeny reanalysed. Nature 476 10.1038/nature10267 (2011)
Liu, J. et al. An armoured Cambrian lobopodian from China with arthropod-like appendages. Nature 470, 526–530 (2011)
Maddison, W. P. & Maddison, D. R. MacClade: Analysis of Phylogeny and Character Evolution Version 3 (Sinauer Associates, 1992)
Swofford, D. L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods) Version 4.0. (Sinauer Associates, 2002)
Faith, D. P. & Cranston, P. S. Probability, parsimony, and Popper. Syst. Biol. 41, 252–257 (1992)
Author information
Authors and Affiliations
Corresponding author
PowerPoint slides
Rights and permissions
About this article
Cite this article
Liu, J., Steiner, M., Dunlop, J. et al. Liu et al. reply. Nature 476, E1 (2011). https://doi.org/10.1038/nature10268
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature10268
This article is cited by
-
Towards intelligent control of influence diffusion in social networks
Social Network Analysis and Mining (2015)
-
Synthesis of rGO–Ag nanoparticles for high-performance SERS and the adsorption geometry of 2-mercaptobenzimidazole on Ag surface
Applied Physics A (2014)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
