
Abstract
Tol1 is an active DNA-based transposable element residing in the genome of the medaka fish Oryzias latipes. This element belongs to the hAT transposable element family, of which complete copies have relatively long sequences. In addition, we found that Tol1 elements as long as 18 and 20 kb occur in the medaka fish genome. These facts suggest that Tol1 is suitable for carrying large DNA fragments as a gene transfer vector. Focusing on this, we conducted two kinds of manipulations of the element. The first was to eliminate internal regions dispensable for transposition. It was revealed that a Tol1 element consisting of 157-bp left- and 106-bp right-terminal regions could be transposed without a loss of transposition efficiency. Next, we prepared long Tol1 elements by incorporating unrelated DNA fragments into this short Tol1 clone and examined their transposition efficiencies. The transposition frequency decreased as the element size increased. The longest Tol1 element we examined measured 22.1 kb, and its transposition frequency was approximately one fifth that of a 2.1-kb element. However, this frequency was still significantly higher than that of a random integration of DNA into the chromosomes. The element size of 22.1 kb is the longest ever reported for DNA-based elements currently used for mammals. Thus, Tol1 is a superior gene-transfer vector with a large cargo capacity.
Similar content being viewed by others

Introduction
DNA-based transposable elements, RNA-mediated elements, retroviruses, and adenoviruses have all been used as tools for genetic manipulation, such as gene transfer, mutagenesis, and gene/promoter/enhancer trapping. A great advantage of DNA-based elements is the simplicity and safety of their usage. Elements of this type currently used for mammals include the naturally occurring elements Tol2 from the medaka fish (Koga et al. 2003; Koga et al. 2006) and piggyBac from a moth (Fraser et al. 1996; Wilson et al. 2007), as well as the resurrected element Sleeping Beauty from salmonid fishes (Ivics et al. 1997; Aronovich et al. 2007). We recently found another element, Tol1 of the medaka fish, which has the potential for development as a genetic tool. The Tol1 element with an embedded neomycin-resistance gene was efficiently transposed into the chromosomes of human and mouse cells, conferring neomycin resistance on the cells (Koga et al. 2007).
It is a general feature of transposable elements that transposition frequency decreases as element size increases. Thus, cargo capacity—the maximum length of a DNA fragment that can be delivered by the element—is an important factor to be considered in choosing an element. The Tol1 element is expected to have an advantage in this regard. First, the element belongs to the hAT (hobo/Activator/Tam3) transposable element family (Calvi et al. 1991). An important feature of this family is that their complete copies are long in comparison with those of other major families, such as the mariner/Tc1 family (cf. Koga et al. 2007). Second, our analysis of the medaka genome revealed the existence of Tol1 copies that are 18 and 20 kb in length, as shown in the initial part of this study. From this observation, we made the inference that Tol1 elements around these sizes can be transposed.
The aim of the present study was to develop the Tol1 element into a gene-transfer vector that has the ability to deliver large DNA fragments into the chromosomes. For this purpose, we first removed internal regions dispensable for the transposition reaction from the original 1.9-kb element (Tol1–tyr, DDBJ accession number D42062), obtaining a basic vector as short as 0.3 kb. We then constructed Tol1 elements of various lengths by inserting extra DNA fragments into the basic vector and measured their transposition frequencies. As previously described (Koga et al. 2007), the Tol1 DNAs were introduced into cells by the lipofection method, and the transposition frequency was determined by counting surviving colonies after selective culturing with G418. Because the efficiency of DNA intake by lipofection is affected by DNA size, we took a measurement to compensate for the size effect: comparisons were made among plasmids of a defined total size that carried Tol1 elements of different sizes. Our results indicated that a Tol1 element of 22.1 kb in length can be efficiently transposed. This is the longest ever reported for DNA-based elements currently used for mammals.
Materials and methods
Genomic library
We prepared a genomic library of an albino medaka fish exhibiting partial melanin pigmentation in the skin and eyes (Koga et al. 2007). This library was used for the selection of Tol1-carrying genomic clones. The vector for the library was pCC1FOS (EPICENTRE Biotechnologies, Madison, WI, USA) and contained 33- to 48-kb mechanically sheared DNA fragments.
Plasmids
Two kinds of plasmids were used: donor plasmids and helper plasmids. In cells, the Tol1 element was excised from the donor plasmid and integrated into the chromosomes by the catalysis of the transposase produced by the helper plasmid. The donor plasmids carried left- and right-terminal regions of the Tol1–tyr element in the plasmid backbone of pUC19. The lengths of these arms varied among experiments. In most cases, the donor plasmids also contained other DNA fragments, such as a marker gene and stuffer fragments. The helper plasmids were the same as those used in our previous study (Koga et al. 2007).
Transposition assays
Human HeLa and mouse NIH/3T3 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (FBS) and antibiotics in an incubator at 37°C with 5.0% CO2. Aliquots of 2 × 105 cells were seeded in 12-well plates (22 mm in diameter) and incubated for 24 h. Mixtures of 100 ng of donor and 900 ng of helper plasmid DNAs per dish were then introduced into the cells using the Lipofectamine LTX reagent (Invitrogen Corp., Carlsbad, NM, USA). After 8 h of incubation, the cells were washed with phosphate-buffered saline (PBS) and incubated for another 24 h in fresh medium without plasmid DNA or transfection reagent. The cells were then trypsinized and suspended in 2.0 ml of medium, and 400-μl aliquots of cell suspension were transferred to dishes of different sizes (35, 60, and 90 mm) containing medium and 500 μg/ml G418. After 12 days under G418 selection, the cells were fixed with 20% formalin and stained with Giemsa’s solution. Cell colonies were counted from dishes containing colony numbers closest to 100, and colony numbers per 105 seeded cells were determined. The transfection assays were carried out in triplicate.
Molecular methods
The molecular methods listed below followed standard procedures (Sambrook et al. 2001) and were also described previously (Koga et al. 2007): preparation of genomic DNA, polymerase chain reaction (PCR), cloning of PCR products, DNA sequencing, and colony hybridization. The PCR enzyme used in this study was LA Taq (Takara Bio Inc., Otsu, Japan). The PCR conditions are described for each case.
Results
Length variation of Tol1 copies
The medaka genome contains 100–200 Tol1 copies, which are not homogenous in length (Koga et al. 1995). To reveal the degree of variation in length, we screened a genomic library for chromosomal fragments containing both terminal regions of Tol1, obtaining 130 such clones. We then conducted PCR to amplify the Tol1 portion in each clone using primers representing the Tol1 terminals. Production of fragments was observed with 114 of the 130 clones. Distribution of the element lengths was roughly unimodal, with a sharp peak at 1–2 kb (S1 of electronic supplementary material). One important finding was that Tol1 elements of about 18 and 20 kb are present in the medaka genome.
Test for transposition activity of short Tol1 clones
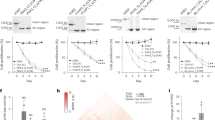
It is possible that the 1.9-kb Tol1–tyr element contains an internal region dispensable for transposition. We thus prepared several shorter versions by PCR and tested their transposition activity by introducing them into mouse culture cells together with either a functional or defective helper plasmid. Importantly, the transposition assays showed that a clone consisting of 157 bp left- and 106 bp right-terminal regions showed a transposition frequency comparable to that of the Tol1–tyr clone (Fig. 1). Further truncation of either the left or right arm to 26 bp led to a significant reduction or loss of transposition activity.

Transposition frequencies of internally deleted Tol1 clones. The tested Tol1 clones are illustrated in the left margin. Only the portions of Tol1 arms are shown, and their lengths are not to scale. Transposition assays were conducted either with the functional helper (shaded bars) or with the defective helper (white bars). The widths of the bars indicate the mean colony numbers from three independent assays, with standard errors of the mean indicated by horizontal lines
Based on the results obtained in the above experiments, we prepared pDon263Mcs containing 157-bp left and 106-bp right Tol1 arms and a multiple cloning site (MCS) for six commonly used restriction endonucleases between the arms (Fig. 2).

Donor plasmids containing Tol1 elements of different sizes. The pDon263Mcs shown at the bottom is the basic vehicle plasmid that carries a multiple cloning site. The pDon263McsNeo shown in the middle was prepared by inserting the Neo R gene into pDon263Mcs. The bars shown at the top are inner and outer stuffer DNA fragments of various lengths prepared by amplifying parts of the DNA of bacteriophage λ (DDBJ J02459) with polymerase chain reaction (PCR). Nucleotide numbers of the amplified regions are shown below the bars. The PCR primers were designed so that they would carry a cutting site for EcoRI or HindIII at their 5′ ends. The stuffer fragments were digested with EcoRI or HindIII and inserted into pDon263McsNeo at their respective sites
Comparison of transposition frequency of Tol1 clones of different lengths
Using PCR, we amplified a DNA fragment of x kb in size (x = 0, 5, 10, 15, 20) and another fragment of y kb (y = 20 − x) as inner- and outer-stuffer fragments, respectively. These fragments were then inserted into the EcoRI (inside Tol1) and HindIII (outside Tol1) cutting sites of the vector pDon263McsNeo (Fig. 2). Each clone was called pDon263McsNeoExHy. The distance between the left and right Tol1 ends on pDon263McsNeoExHy was (x + 2.1 kb), and the size of the entire plasmid was 24.8 kb, irrespective of x.
It is known that plasmid size is a factor that affects the efficiency of DNA intake in the lipofection method. The incorporation of outer-stuffer fragments in addition to inner-stuffer fragments served to eliminate the effects of the plasmid DNA size. This was done to allow an accurate comparison of the transposition frequency among different donor plasmids.
Transposition frequencies were determined for the five donor plasmids, each with the functional and defective helper plasmids (Fig. 3). In human and mouse cells alike, a negative correlation between element size and transposition frequency was observed. In the case of transfection with the functional helper plasmid, the ratio of frequency of the longest element (pDon263McsNeoE20) to that of the shortest element (pDon263McsNeoH20) was 0.21 and 0.28 in human and mouse cells, respectively. Transposition frequencies of the longest element transfected with the functional helper plasmid were eight and ten times higher than when transfected with the defective helper plasmid in human and mouse cells, respectively.

Transposition frequencies of long donor plasmids. Transposition assays were conducted either with the functional helper plasmid (shaded bars) or with the defective helper plasmid (white bars). The donor plasmids used are shown in the form of ExHy in the left margin. Bar width indicates the mean colony numbers from three independent assays, with standard errors of the mean indicated by horizontal lines
Demonstration of transposition
We went on to confirm that the Tol1 element was integrated into the chromosomes by transposition. For this purpose, we first isolated two mouse-cell colonies produced in an assay with the longest element (pDon263McsNeoE20). These cell lines were propagated in different dishes, and their genome DNAs were extracted. Using these DNAs as templates, we amplified the Tol1 terminals and their flanking chromosomal regions by inverse PCR and sequenced the fragments obtained. Alignment of the sequences revealed the creation of 8-bp target-site duplications in both cases (Fig. 4), indicating that the Tol1 element of the donor plasmid was integrated into the recipient DNAs by means of transposition.

Nucleotide sequences around insertion sites. Genomic DNA was extracted from two neomycin-resistant cell clones and digested with HindIII, whose cutting site is not present inside the To1l portion of the donor plasmid. DNA fragments of approximately 10–30 kb in size were isolated from electrophoresis gels and ligated with T4 DNA ligase. Inverse polymerase chain reaction (PCR) was performed on the ligated DNA using primers representing parts of the Tol1 arms (nt 130–101 and nt 1758–1787 of D42062). PCR conditions were as follows: [94°C, 2 min], 36 × [94°C, 20 s; 64°C, 20 s; 72°C, 5 min], and [72°C, 5 min]. The PCR products were cloned into plasmids and sequenced. Nucleotide sequences around the insertion sites of these genomic clones are shown. The corresponding donor plasmid sequence is also shown at the top as reference. The black- and gray-shaded regions are Tol1 arms and target side duplications (TSDs), respectively
The results did not necessarily mean that the entire Tol1 element, including the inner stuffer fragment, was integrated into the chromosomes without internal deletion or aberration. To confirm this, we cloned and analyzed the inserted Tol1 element. Cloning was performed by PCR using primers that straddled the Tol1-terminal regions and their flanking chromosomal regions. The obtained PCR products were of the expected size, and their restriction enzyme digestion patterns were also as expected (S2 of ESM). These results indicate that the entire region of the 22.1 kb Tol1 element was integrated into the chromosomes.
Discussion
We constructed a basic Tol1 vector consisting of 157 bp left- and 106 bp right-terminal regions, which is transposed as efficiently as the original 1,855-bp element. Thus, we were successful in removing an internal 1,592-bp region. This should be helpful in creating more room for cargo and in eliminating possible signals and signal-like sequences that might interact with cargo DNA or the host cell.
Although it might be possible to further shorten the arms of the vector by additional fine analyses, we planned to retain some extra arm regions in the vector because the regions could be used as a tag in DNA analyses after insertion. In many cases, the first step of these analyses is cloning of flanking chromosomal regions, and the principal method for this would be inverse PCR, in which parts of the arms are used for primer regions. Thus, we carefully left nucleotide blocks that can be used for PCR primers. Arm lengths (157 and 106 bp) were decided upon based on this reasoning.
We conducted transposition assays both with a functional helper plasmid and a defective helper plasmid. The numbers of surviving colonies with the defective helper were almost the same among the five donor plasmids. This result indicates that the effects of the plasmid size on the DNA intake efficiency of lipofection could be eliminated sufficiently in our assay system.
Our analysis of transformant cell colonies indicated that the entire Tol1 element had been integrated into the chromosomes by transposition. The integration frequency was significantly higher than that by random integration, even in the case of the longest Tol1 element (pDon263McsNeoE20). The size of the Tol1 portion of this donor plasmid was 22.1 kb, of which 0.3 kb was occupied by the Tol1 arms. Thus, our basic plasmid pDon263Mcs can deliver a DNA fragment of up to 21.8 kb into the chromosomes, at least in the culture cells we used. It is also a significant finding that the DNA fragment delivered did not suffer internal deletion or aberration.
The Sleeping Beauty element is known to lose efficiency of transposition when the element size exceeds 9.1 kb (Karsi et al. 2001). The piggyBac element has been shown to serve as a gene-transfer vector when the entire size is 14.3 kb (Ding et al. 2005). In the case of the Tol2 element, the maximum element size so far reported is 10.2 kb (Balciunas et al. 2006). It is possible that the latter two elements retain transposition activity when they are longer than the reported sizes. At present, the element size of 22.1 kb that we have shown with Tol1 in this study is the largest ever demonstrated with DNA-based elements currently used for mammals. Furthermore, the length of the Tol1 arms in our basic plasmid is as short as 0.3 kb. Thus, Tol1 is a useful gene-transfer vector for delivering large DNA fragments into chromosomes of mammals.
References
Aronovich EL, Bell JB, Belur LR, Gunther R, Koniar B, Erickson DC, Schachern PA, Matise I, McIvor RS, Whitley CB, Hackett PB (2007) Prolonged expression of a lysosomal enzyme in mouse liver after Sleeping Beauty transposon-mediated gene delivery: implications for non-viral gene therapy of mucopolysaccharidoses. J Gene Med 9:403–415
Balciunas D, Wangensteen KJ, Wilber A, Bell J, Geurts A, Sivasubbu S, Wang X, Hackett PB, Largaespada DA, McIvor RS, Ekker SC (2006) Harnessing a high cargo-capacity transposon for genetic applications in vertebrates. PLoS Genet 2:e169
Calvi BR, Hong TJ, Findley SD, Gelbart WM (1991) Evidence for a common evolutionary origin of inverted repeat transposons in Drosophila and plants: hobo, Activator, and Tam3. Cell 66:465–471
Ding S, Wu X, Li G, Han M, Zhuang Y, Xu T (2005) Efficient transposition of the piggyBac (PB) transposon in mammalian cells and mice. Cell 122:473–483
Fraser MJ, Ciszczon T, Elick T, Bauser C (1996) Precise excision of TTAA-specific lepidopteran transposons piggyBac (IFP2) and tagalong (TFP3) from the baculovirus genome in cell lines from two species of Lepidoptera. Insect Mol Biol 5:141–151
Ivics Z, Hackett PB, Plasterk RH, Izsvak Z (1997) Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in human cells. Cell 91:501–510
Karsi A, Moav B, Hackett P, Liu Z (2001) Effects of insert size on transposition efficiency of the Sleeping Beauty transposon in mouse cells. Mar Biotechnol 3:241–245
Koga A, Iida A, Hori H, Shimada A, Shima A (2006) Vertebrate DNA transposon as a natural mutator: the medaka fish Tol2 element contributes to genetic variation without recognizable traces. Mol Biol Evol 23:1414–1419
Koga A, Inagaki H, Bessho Y, Hori H (1995) Insertion of a novel transposable element in the tyrosinase gene is responsible for an albino mutation in the medaka fish, Oryzias latipes. Mol Gen Genet 249:400–405
Koga A, Shimada A, Kuroki T, Hori H, Kusumi J, Kyono-Hamaguchi Y, Hamaguchi S (2007) The Tol1 transposable element of the medaka fish moves in human and mouse cells. J Hum Genet 52:628–635
Koga A, Iida A, Kamiya M, Hayashi R, Hori H, Ishikawa Y, Tachibana A (2003) The medaka fish Tol2 transposable element can undergo excision in human and mouse cells. J Hum Genet 48:231–235
Sambrook J, Russell DW (2001) Molecular cloning: a laboratory manual, 3rd Edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor
Wilson MH, Coates CJ, George AL Jr (2007) PiggyBac transposon-mediated gene transfer in human cells. Mol Ther 15:139–145
Acknowledgments
We are grateful to Akira Tachibana and Samuel S. Chong for their helpful discussion. Human HeLa cells and mouse NIH/3T3 cells were obtained from the HSRRB of the Japan Health Sciences Foundation (Registry Nos. JCRB9004 and JCRB0615, respectively). This work was supported by grant no. 19570003 from the Ministry of Education, Culture, Sports, Science and Technology of Japan to A. K., and a grant from the Yamada Science Foundation to A. K.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Rights and permissions
About this article
Cite this article
Koga, A., Higashide, I., Hori, H. et al. The Tol1 element of medaka fish is transposed with only terminal regions and can deliver large DNA fragments into the chromosomes. J Hum Genet 52, 1026–1030 (2007). https://doi.org/10.1007/s10038-007-0213-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10038-007-0213-7
Keywords
This article is cited by
-
Tandem repeat sequences evolutionarily related to SVA-type retrotransposons are expanded in the centromere region of the western hoolock gibbon, a small ape
Journal of Human Genetics (2012)
-
Delivering the goods: viral and non-viral gene therapy systems and the inherent limits on cargo DNA and internal sequences
Genetica (2010)
-
Transposon-mediated genome manipulation in vertebrates
Nature Methods (2009)
-
The Tol1 element of the medaka fish, a member of the hAT transposable element family, jumps in Caenorhabditis elegans
Heredity (2008)
