
ABSTRACT
In the present study expression of estrogen receptor subtype - α(ER α) and - β (ER β) in the cerebral cortex, cerebellum, and olfactory bulb was investigated and compared between neonatal (1 ∼ 3-days-old) and adult (250 ∼ 350g) rats, using reverse transcription-polymerase chain reaction (RT-PCR). No ERa transcripts were detectable in the adult cerebellum and olfactory bulb, whereas very weak expression of ERa was present in the adult cerebral cortex. No significant difference in ERb transcripts was detectable between the neonatal and adult rats. While transcripts for both ER subtypes were co-expressed in these brain areas of neonatal rats, although ERa expression was significantly weaker than ER β. Even in the cerebral cortex known to contain both ER subtypes in adult rats, ERa transcripts in neonatal rats were much higher than in adult. These observations provide evidence for the existence of different expression patterns of ER α/ER βtranscripts in these three brain areas between the neonatal and adult rats, suggesting that each ER subtype may play a distinct role in the regulation of differentiation, development, and functions of the brain by estrogen.
Similar content being viewed by others

INTRODUCTION A
Estrogen has been known to exert extensive effects via estrogen receptor (ER) on diverse physiological and developmental functions of the brain1, 2. It has been observed that the distribution of the classical ER subtype-α(ERα) and the recently characterized novel ER subtype-β (ERb), and their expression patterns (ER α/ER β) vary greatly among various brain regions1,3. These evidences suggest that each ER subtype may play a different role in estrogen's effects on the brain.
Previous studies on ER in the brain, however, have been done mainly in adult animals. Little information is available concerning ER in neonate, as well as the differences of ER α/ER βexpression patterns between the neonatal and adult brain. A previous work revealed that there existed changes of ER α/ER β expression patterns in hippocampus4 between neonatal (1 ∼ 3days) and adult (250 ∼ 350g) rats. The present study was designed to compare the presence and relative expression level of ER α and ER βmRNA in the cerebral cortex, cerebellum, and olfactory bulb of neonatal (1 ∼ 3days-old) rats with that of adult (250 ∼ 350g) rats, by use of highly sensitive reverse transcription-polymerase chain reaction (RT-PCR) techniques. The brain tissues of cerebellum and olfactory bulb were selected to be used in the present study mainly because previous study3 demonstrated only ER β and no ERa expression in these brain areas of adult rats. We wonder whether it is also the same case for ER expression in these brain regions of neonatal rats. The cerebral cortex known to contain both ER subtypes in adult rats3 was also used for comparison.
MATERIALS AND METHODS
RNA extraction
Brain tissue collection was done in deeply anesthetized (sodium pentobarbital, 100 mg/kg) 12 neonatal (1 ∼ 3-days-old) of either sex, and adult (250-350g) 3 male and 3 female Sprague-Dawley rats. Intact adult males and females were gonadectomized under anesthesia (sodium pentobarbital, 70 ∼ 80mg/kg) and allowed to recover for one week before sacrifice. Animals were transcardially perfused with 150ml (for adult rats) and 30 ml (for neonatal rats) of diethylpyrocarbonate (DEPC)-treated (1%) 0.9% NaCl to eliminate possible ER transcripts that may exist in the blood cells5. Entire brain tissues of cerebral cortex, cerebellum, and olfactory bulb pooled from 2 neonatal rats or one adult rat were respectively collected in one tube. Tissue samples (50 ∼ 100mg mixed tissues for each) were immediately processed for total RNA isolation and purification with Trizol (Gibco BRL). The integrity and quality of total RNA were analyzed by formaldehyde denaturing argarose gel electrophoresis (sharp bands of 18s and 28s rRNA) and by measurement of the A260/A280 nm ratio (> 1.8) 6.
Reverse transcription (RT)
2mg of total RNA (determined by spectrophotometer) together with 1 μM oligo-dT (18mer) and 1ml random hexamer primers (0.2mg/ml) was denatured at 65°C for 5min. The RNA was added to a mixture of RT buffer containing 0.5 μM of each dNTP, 10U of RNasin, and 4U of Omniscript reverse transcriptase (Qiagen, Germany) in a final volume of 20 μl. The reaction was incubated for 1 hr at 37°C, after which the enzyme was inactivated at 93°C for 5min and then at 0°C for 2min. The cDNA samples were stored at −20°C.
Polymerase chain reaction (PCR)
Design and synthesis of specific sense and antisense primers for ERa, ERb , and internal control (glyceraldehyde-3-phosphate dehydrogenase, GAPDH) were based on the published article7 and rat DNA sequence information in GeneBank with the aid of Genamics Expression software (Ver.1.1). ERa sense: 5′-CCTAACTTGCTCTTGGACAGG-3′; ERa antisense: 5′-CAGCAGCAGGTCATAGAGAGG-3′; ERb sense: 5′-GCCAATCATGTGCACCAGTTCCTT-3′; ERb antisense: 5′-AAAGCCAAGAGAAACGGTGGGCAT-3′. GAPDH sense: 5′-TCAACGGCACAGTCAAGGC-3′; GAPDH antisense: 5′-AGGGATGATGTTCTGGGCTG-3′.
The reaction consisted of 0.5 μl of cDNA, 5 μl 10×PCR buffer, 0.1mM dNTPs, 1pM of each primer, and 1U of Taq polymerase (TaKaRa, Japan) in a total volume of 50 μl. The PCR was run for 35 cycles (each cycle: denaturation, 40s at 94°C; annealing, 40s at 59°C; elongation, 40s at 72°C) with an initial denaturation of 5 min at 94°C and a final extension at 72°C for 10 min. The PCR products were collected and cloned in Pucm-T vector (Sangon, Canada), and all of the DNA sequences of the regions amplified for ERa, ERb and GAPDH were analyzed and verified to fit with the published sequences of their counterparts in GeneBank.
Gel electrophoresis and controls
PCR products (20 μl for each) with 2 μl of 10×DNA loading buffer were electrophoresed on 1.7% argarose gels in 1×TAE buffer, stained with ethidium bromide (EB), and photographed by digital camera (GIS-1000, Tanon).
As additional controls for amplification of only cDNA, mock cDNA synthesis (no RT) or PCR lacking template was performed, and no detectable product (equals to background) was generated from any of the negative control reactions.
Values determination and statistical analysis
For each cDNA sample analyzed, a minimum of 3 independent PCR was performed. Such a performance was repeated six times from different animals. Values were determined from each sample normalized to internal control GAPDH following local area background subtraction. Difference in fluorescence resulting from differences in the number of base pairs of ERa (408bp) and ERβ (204bp) products (408 vs 204) were corrected by using a factor of 0.5 for ERa. The value (expressed as mean ± s.e.m) was averaged from data of all individual reactions for each group, and then normalized to the average value (as 1) of ERb (showing the highest level) of neonatal cerebral cortex (NCC).
Statistical analysis was performed using a three-way analysis of variance (ANOVA; P < 0.05; factors =age, ER subtype, and brain area), followed by planned pairwise comparisons using Fishers post hoc analysis (P < 0.05), by using the statistical package, SPSS (V10.0) for windows.
RESULTS AND DISCUSSION
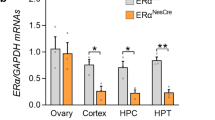
RT-PCR amplification of cDNA, which was reversely transcribed from total RNA isolated from either the neonatal or adult brain tissues, resulted in production of DNA products of the predicted size for ER α (408bp), ER β (204bp), and GAPDH (458bp) (Fig 1). No sex differences of ER mRNA expression were apparent, and therefore, the data from both sexes were pooled and reflect observations of both male and female brain tissues. Comparison of the fluorescent intensity of EB-staining of RT-PCR products indicated that no significant expression differences of ER βtranscripts existed in the cerebral cortex, cerebellum (Fig 1b), and olfactory bulb (Fig 1d) between the neonatal (1 ∼ 3days) and adult (250 ∼ 350g) rats (P > 0.05) (Fig 2). However, it was not the case for ERa. No ERa transcripts were seen in the tissues of adult rat cerebellum (Fig 1a) and olfactory bulb (Fig 1d), although very weak ERa transcripts could be seen in the adult rat cerebral cortex (Fig 1a). Whereas it is worthy of noting that they could be seen clearly in either of these three brain areas of neonatal rats, although significantly weaker than ERβ transcripts (P < 0.01) (Fig 1a,d; Fig 2). Even in the cerebral cortex known to contain both ER subtypes in adult rats3, ERα transcripts in neonatal rats were much higher than in adult (P < 0.01) (Fig 1a; Fig 2).

Typical digital photographs of fractionation of RT-PCR products amplified from cDNA of transcripts for either ERα or ERβ in the cerebral cortex, cerebellum, and olfactory bulb between neonatal and adult rats. Note that no ERα transcripts can be detected in the adult cerebellum (AC) and olfactory bulb (AOB), although very weak expression of ERα transcripts is present in the adult rat cerebral cortex (ACC). Whereas ERα transcripts can be detected clearly, although significantly weaker than ERβ, in the neonatal rat cerebellum (NC), olfactory bulb (NOB), and cerebral cortex (NCC). Even in the cerebral cortex, ERα transcripts in neonatal rats are much higher than in adult. No significant expression differences of ER β transcripts were detectable in these brain areas between the neonatal and adult rats. (a) ERα in the cerebellum and cerebral cortex; (b) ERa in the cerebellum and cerebral cortex; (c) GAPDH in the cerebellum and cerebral cortex; (d) ERα and ERα in the olfactory bulb. (e) GAPDH in the olfactory bulb. M, marker.

Analysis of the relative fluorescent intensity of ERα and ER β products. For determination of values and results of statistical analysis please see text. Abbreviations are the same as used in Fig 1.
This RT-PCR study is in agreement with previous reports that ERβ mRNA3 and its protein immunoreactivity8 were robustly present, and no ERα3,9 was evidently found, in the cerebellum and olfactory bulb of adult rats, as demonstrated by in situ hybridization and immunohistochemistry techniques. To our surprise, the present highly sensitive RT-PCR study demonstrated that ERα transcripts existed, although in significantly lower degree than ERb, in the neonatal (at least 1 ∼ 3-days-old rats) cerebellum and olfactory bulb. Even in the cerebral cortex known to contain both ER subtypes in adult rats3,9, ERα transcripts in neonate were much higher than in adult rats, which is similar to the change of ER expression in the hippocampus as revealed by Western blot analysis4. One possible explanation for this discrepancy is that there may exist obvious changes of ERα expression across the structural and functional development of these brain areas from neonatal to adult. In contrast to ERα, no significant expression change of ERβ transcripts was seen in these three brain areas between neonatal and adult rats. Previous observations reported that ERb also exhibited developmental changes even during the postnatal period in the cerebellum10,11 and olfactory bulb[12] Our experiments only observed the differences of ER expression between neonatal (1 ∼ 3-days-old) and adult (250 ∼ 350g) rats. Hence, our conclusion could not exclude the possibility that ERb also runs a changing profile across the developmental course from neonatal to adult.
Although the detected variations in ER mRNA expression were not confirmed at the protein level, the present results confidently suggest that the relative expression of ERα and ERβ could not only exist among various brain regions, but also be not static developmentally with age. Nothing has yet been known of the regulation of the relative expression of ERα and ERβ in the brain. It seems that both ER subtypes are co-expressed in these brain areas at least in the early postnatal stage, and then ERα expression is lowered to the degree even below the detection levels of some methods available at present such as immunocytochemistry, in situ hybridization, and RT-PCR as seen in the present observations, as animals grow into adult. Although comparison of just two ages gives only a small snapshot of developmental changes of ER expression, the results strongly suggest that each ER subtype may play a distinct role in these brain regions. Most likely mechanism is that either ER subtype is expressed in different brain regions with distinct ontogenic profile and is governed by intrinsic developmental programs13.
Our data both in neonatal and adult rats are consistent with the notion that ERβ is the predominant ER species expressed3,9, 11 and suggest that ERβ isoform other than ERα plays a major role in the regulation of these three brain areas by estrogen. The clear manifestation of ERα mRNA in the neonatal corresponding brain areas, however, merits attention to its role in the regulation of early postnatal differentiation and development of these brain areas by estrogen, since ERα is supposed closely related to cellular differentiation and sexual differentiation of developing brain14,15.
References
Donahue JE, Stopa EG, Chorsky RL et al. Cells containing immunoreactive estrogen receptor-a in the human basal forebrain. Brain Res 2000; 856:142–51.
Shughrue PJ, Merchenthaler I . Estrogen is more than just a “sex hormone”: novel sites for estrogen action in the hippocampus and cerebral cortex. Front-Neuroendocrinol 2000; 21:95–101.
Shughrue PJ, Lane MV, and Merchenthaler I . Comparative distribution of estrogen receptor -α and -β mRNA in the rat central nervous system. J Comp Neurol 1997; 388:507–25.
Su JD, Qiu J, Zhong YP, Li XY, Wang JW, and Chen YZ . Expression of estrogen receptor (ER)-α and β immunoreactivity in hippocampal cell cultures with special attention to GABAergic neurons. J Neurosci Res 2001; 65:396–402.
Tornwall J, Carey AB, Fox RI, Fox HS . Estrogen in autoimmunity: expression of estrogen receptors in thymic and autoimmune T cells. J Gend Specif Med 1999; 2:33–40.
Ma YH, Hu JH, Zhou XG et al. g-Aminobutyric acid transporter (GAT1) overexpression in mouse affects the testicular morphology. Cell Res 2000; 10:59–69.
Sugino N, Zilberstein M, Srivastava RK et al. Establishment and characterization of a simian virus 40-transformed temperature-sensitive rat luteal cell line. Endocrinolgy 1998; 139:1936–42.
Shughrue PJ, Merchenthaler I . Distribution of estrogen receptor-b immunoreactivity in the rat central nervous system. J Comp Neurol 2001; 436:64–81.
Laflamme N, Nappi RE, Drolet G, Labrie C, Revest S . Expression and neuropeptidergic characterization of estrogen receptors (ERa and ERb) throughout the rat brain: anatomical evidence of distinct roles of each subtype. J Neurobiol 1998; 36:357–78.
Belcher SM . Regulated expression of estrogen receptor a and b mRNA in granule cells during development of the rat cerebellum. Brain Res Dev Brain Res 1999; 115:57–69.
Jakab RL, Wong JK, and Belcher SM . Estrogen receptor b imunoreactivity in differentiating cells of the developing rat cerebellum. J Comp Neurol 2001; 430:396–409.
Wong CC, Poon WH, Tsim TY, Wong EYK, Leung MS . Gene expressions during the development and sexual differentiation of the olfactory bulb in rats. Brain Res Dev Brain Res 2000; 119:187–94.
O'Keefe JA, Pedersen EB, Castro AJ, Handa RJ . The ontogeny of estrogen receptors in heterochronic hippocampal and neocortical transplants demonstrates an intrinsic developmental program. Brain Res Dev Brain Res 1993; 75:105–12.
Wagner CK, Pfau JL, De Vries GJ, Merchenthaler IJ . Sex differences in progesterone receptor immunoreactivity in neonatal mouse brain depend on estrogen receptor a expression. J Neurobiol 2001; 47:176–82.
Yokosuka M, Okamura H, Hayashi S . Postnatal development and sex difference in neurons containing estrogen receptor-a immunoreactivity in the preoptic brain, the diencephalon, and the amygdala in the rat. J Comp Neurol 1997; 389:81–93.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (No. 39700053).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
GUO, X., SU, J., SUN, Q. et al. Expression of estrogen receptor (ER) - α and - β transcripts in the neonatal and adult rat cerebral cortex, cerebellum, and olfactory bulb. Cell Res 11, 321–324 (2001). https://doi.org/10.1038/sj.cr.7290103
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/sj.cr.7290103
Keywords
This article is cited by
-
Regulatory Control of Microglial Phagocytosis by Estradiol and Prostaglandin E2 in the Developing Rat Cerebellum
The Cerebellum (2019)
-
Effects of maternal chlorpyrifos diet on social investigation and brain neuroendocrine markers in the offspring – a mouse study
Environmental Health (2015)
-
Sex Steroids Effects on the Content of GAD, TH, GABAA, and Glutamate Receptors in the Olfactory Bulb of the Male Rat
Neurochemical Research (2008)
