
Abstract
Employing a two-lever, food-reinforced, Fixed Ratio 10 drug discrimination procedure, rats were trained to recognize the highly-selective serotonin (5-HT)2A receptor antagonist, MDL100,907 (0.16 mg/kg, i.p.). They attained criterion after a mean ± S.E.M. of 70 ± 11 sessions. MDL100,907 fully generalized with an Effective Dose (ED)50 of 0.005 mg/kg, s.c.. A further selective 5-HT2A antagonist, SR46349, similarly generalized with an ED50 of 0.04 mg/kg, s.c. In distinction, the selective 5-HT2B antagonist, SB204,741 (0.63 and 10.0 mg/kg), the 5-HT2B/2C antagonist, SB206,553 (0.16 and 2.5 mg/kg) and the selective 5-HT2C antagonists, SB242,084 (2.5 and 10.0 mg/kg,) and RS102221 (2.5 and 10.0 mg/kg), did not significantly generalize. In conclusion, selective blockade of 5-HT2A receptors by MDL100,907 elicits a discriminative stimulus in rats which appears to be specifically mediated via 5-HT2A as compared with 5-HT2B and 5-HT2C receptors.
Similar content being viewed by others


Main
Drug discrimination procedures have been extensively used in the characterization of psychoactive agents, including drugs interacting with 5-HT reuptake sites (Millan et al. 1999b) and agonists at 5-HT1A (Schreiber et al. 1995b) and 5-HT3 (Glennon et al. 1992) receptors. Although it has proven difficult to differentiate the roles of closely-related 5-HT2A, 5-HT2B and 5-HT2C receptors (Glennon 1991), it was suggested that discriminative stimulus (DS) properties of several 5-HT2 agonists and hallucinogens, such as mescaline (Appel and Callahan 1989), lysergic acid diethylamide (LSD) (Fiorella et al. 1995) and quipazine (Friedman et al. 1984), are mediated by 5-HT2A receptors. Further, use of the highly selective 5-HT2A receptor antagonist, MDL100,907 (Kehne et al. 1996), demonstrated that 5-HT2A receptors mediate DS properties of 1-[2,5-dimethoxy-4-iodophenyl]-2-aminopropane (DOI), a further hallucinogen (Schreiber et al. 1994). In contrast, DS properties of the 5-HT2 ligand, m-chlorophenylpiperazine (mCPP), appear to be mediated by 5-HT2C receptors (Callahan and Cunningham 1994; see Gommans et al. 1998), and employing the 5-HT2B/2C antagonist, SB206,553, and the selective 5-HT2C antagonist, SB242,084, it was shown that 5-HT2C receptors likewise mediate DS properties of the novel, mixed 5-HT2C/2B agonist, RO60,0175 ((S)-2-(6-chloro-5-fluoroindol-1-yl)-1-methylethylamine; Dekeyne et al. 1999).
Interest in 5-HT2A receptors has been reinforced by evidence that their blockade may contribute to the distinctive functional profile of the “atypical” antipsychotic, clozapine, and, possibly, other novel agents employed for management of psychotic states (Roth and Meltzer 1995). Further, clozapine and several other antipsychotics block DS properties of 5-HT2A agonists such as DOI (Palumbo and Winter 1994; Schreiber et al. 1994), while 5-HT2A receptors are, at least partially, involved in the DS properties of clozapine itself (Millan et al. 1999c).
It would, thus, be of considerable interest to establish whether discrete blockade of 5-HT2A receptors generates a specific DS in rats. In view of its pronounced selectivity for 5-HT2A receptors, the potential antipsychotic agent, MDL100,907, appeared an optimal ligand to address this issue (Kehne et al. 1996; Millan et al. 1999c). In the present study, we evaluated whether MDL100, 907 elicits a reliable DS in rats. We also characterized the role of 5-HT2A as compared with 5-HT2B and 5-HT2C receptors in the mediation of its potential DS properties. Generalization testing was conducted with a further selective and potent antagonist at 5-HT2A receptors, SR46349 (Rinaldi-Carmona et al. 1992), the 5-HT2B/2C antagonist, SB206,553 (Kennett et al. 1996), the selective 5-HT2B antagonist, SB204,741 (Forbes et al. 1995), and the novel selective 5-HT2C antagonists, RS102221 and SB242,084 (Bonhaus et al. 1997; Kennett et al. 1997).
METHODS
All animal use procedures conformed to international European ethical standards (86/609-CEE) and the French National Committee (décret 87/848) for the care and use of laboratory animals. As described previously (Millan et al. 1999b), male Wistar rats (180–200 g upon arrival, Iffa-Credo, L'Arbresle, France) were trained to discriminate MDL100,907 (0.16 mg/kg, i.p.) from saline in operant conditioning chambers equipped with two levers. The training dose was selected in light of the ability of MDL100,907 to abolish actions mediated by 5-HT2A receptors in other paradigms without exerting significant effects at other receptors (Schreiber et al. 1994, 1995a; Millan et al. 1999a; Gobert et al. 2000).
The animals were reinforced with food according to a Fixed Ratio 10 schedule of reinforcement. Each 15-min daily session (five days/week) started 15 min after injection. “MDL100,907” or “saline” sessions alternated randomly. Correct responding was defined as no more than 13 presses on both levers to obtain the first reinforcement. The discrimination criterion was ten consecutive sessions with correct responding. Thereafter, generalization tests were conducted every Wednesday and Friday, whereas training sessions were continued on the other days. Rats were tested only if they showed correct responding on the two preceding training sessions. Test drugs were administered instead of MDL100,907, 15 min before the session. During testing, responding on the selected lever, i.e., the lever for which ten responses were recorded first, was reinforced for the remainder of the 15-min session.
All drug doses are in terms of the base. Test drugs were dissolved in sterile water and administered s.c., except SB204,741 and SB242,084, which were administered i.p. as suspensions in water with a few drops of Tween 80. In order to avoid potential cutaneous toxicity, RS102221 and SB206,553 were also administered i.p. at the highest dose tested. Drug structures, salts and sources were as follows. RS102221 (8-[5-(2,4-dimethoxy -5-(4-trifluoromethylphenyl sulphonamino)phenyl-5-oxopentyl]-1,3,8-triazaspiro[4.5]decane-2,4-dione) HCl was purchased from Tocris Cookson (Bristol, UK), SB206,553 (5 methyl-1-(3-pyridil-carbamoyl)-1,2,3,5-tetrahydropyrrolo[2,3-f] indole) HCL was purchased from Sigma (Chesnes, France) and SR46349B (1(Z)-[2-(dimethylamino)ethoxyimino]-1-(2-fluorophenyl)-3-(4-hydroxyphenyl)-2(E)-propene) hemifumarate was a generous gift of Sanofi Winthrop (Montpellier, France). MDL100, 907 (R(+)-α-(2,3-dimethoxyphenyl)-1-[2-(4-fluorophenylethyl)]-4-piperidine-methanol), SB204,741 (1-(1-methylindol-5-yl)-3-(3-methylisothiazol-5-yl)urea) and SB242, 084 (6-chloro-5-methyl-1-[6-(2-methylpyridin-3-yloxy) pyridin-3-yl carbamoyl) indoline HCl were synthetized by Servier chemists (G. Lavielle and J.-L. Péglion).
RESULTS
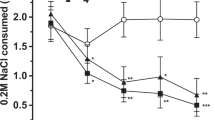
The mean ± S.E.M. number of sessions required to reach the discrimination criterion was 70 ± 11 (n = 7). Administered s.c., MDL100,907 displayed dose-dependent, significant and full generalization (0.16 mg/kg, s.c., 100%, n = 7, p < .05 in Fisher's Exact Probability Test as compared with control training session), an action mimicked by a further selective 5-HT2A antagonist, SR46349 (0.16 mg/kg, s.c., 80%, n = 5, p < .05) (Figure 1 ). In distinction, the selective 5-HT2B antagonist, SB204,741 (0.63 mg/kg, i.p., 0%, n = 5 and 10 mg/kg, i.p., 0%, n = 5), the 5-HT2B/2C antagonist, SB206,553 (0.16 mg/kg, s.c., 40%, n = 5; 2.5 mg/kg, s.c., 20%, n = 5 and 10 mg/kg, i.p., 40%, n = 5), and the selective 5-HT2C antagonists, SB242,084 (2.5 mg/kg, i.p., 20%, n = 5 and 10 mg/kg, i.p., 20%, n = 5) and RS102221 (2.5 mg/kg, s.c., 0%, n = 5 and 10 mg/kg, i.p., 40%, n = 5), did not show significant generalization. None of these antagonists decreased response rates (not shown), with the exception of SB204,741 (10 mg/kg, i.p.) for which a decrease of 5% as compared with the preceding saline training session was observed.

Discriminative stimulus properties of the selective 5-HT2A antagonist, MDL100,907 (0.16 mg/kg, i.p.), in rats. Panel A: Acquisition. Panel B: Generalization of MDL100,907 and SR46349. Data in Panel A are means ± S.E.M. and are shown separately for saline and MDL100,907 sessions. In Panel B, data are percentage of animals generalizing. The Effective Dose50s (95% Confidence Limits) for generalization of MDL100,907 and SR46349 were respectively 0.005 (0.001–0.018) and 0.04 (0.01–0.11) mg/kg, s.c.. Asterisks in Panels B indicate significance of differences to control sessions in Fisher's Exact Probability Test. * p < .05.
DISCUSSION
The present study demonstrates that MDL100,907 elicits a specific and stable DS in rats. Although the 5-HT2 antagonist, pizotifen, has been shown to generate a DS in rats, it does not differentiate 5-HT2A from 5-HT2B and 5-HT2C sites (Minnema et al. 1984). Further, several 5-HT2 antagonists failed to generalize to pizotifen, which interacts with many other sites, including histaminergic receptors, which play a major role in its DS properties (Minnema et al. 1984). Thus, the present study constitutes the first demonstration, to our knowledge, that selective blockade of 5-HT2A receptors can sustain a DS.
Support for a specific role of 5-HT2A receptors in the DS properties of MDL100,907 is provided by several lines of evidence. First, MDL100,907 is >200-fold selective for native, tissue and cloned, human 5-HT2A versus 5-HT2B and 5-HT2C receptors (Table 1). Second, the training dose of MDL100,907, as well as the Effective Dose50 for its “auto-generalization”, were low. These doses correspond well to actions of MDL100,907 in other models of 5-HT2A receptor-mediated responses: notably, inhibition of a DOI-induced DS, blockade of DOI-induced head-twitches and antagonism of hyperlocomotion elicited by phencyclidine (Schreiber et al. 1994, 1995a; Millan et al. 1999a). At these doses, MDL100,907 does not exert significant action in functional models of 5-HT2C receptor-mediated activity (Millan et al. 1997; Dekeyne et al. 1999). Third, a further, highly-selective and potent antagonist at 5-HT2A receptors, SR46349, similarly generalized to MDL100,907 at low doses corresponding to those active in the above-mentioned models (Rinaldi-Carmona et al. 1992; Millan et al. 1999c). Fourth, at doses producing robust increases of extracellular norepinephrine and dopamine levels in the frontal cortex of freely-moving rats, and which abolish 5-HT2C receptor-mediated DS and penile erections, both SB206,553 and SB242,084 (Kennett et al. 1997; Millan et al. 1997; Dekeyne et al. 1999; Gobert et al. 2000), did not generalize to MDL100,907. A further, novel 5-HT2C antagonist, RS102221 (Bonhaus et al. 1997) also did not generalize. Fifth, SB206,553 is a potent antagonist at 5-HT2B as well as 5-HT2C receptors (Kennett et al. 1996), and a further (selective) antagonist at this site, SB204,741 (Forbes et al. 1995; Dekeyne et al. 1999), similarly did not generalize to MDL100,907.
These observations provide compelling evidence that selective blockade of 5-HT2A receptors mediates the DS properties of MDL100,907, although the possibility that other receptor types are (indirectly) involved in their expression would be of interest to evaluate further. It will be of interest to characterize generalization patterns of clozapine and other antipsychotic agents, as well as other classes of psychoactive drug known to interact with 5-HT2A receptors, such as antidepressant agents (Frazer 1997).
In conclusion, the present study shows that MDL100, 907 elicits a stable DS in rats that appears to be mediated by blockade of 5-HT2A receptors. Further characterization of the interoceptive properties of MDL100,907 may provide important insights into the actions of antipsychotic agents and other drug classes which interact with 5-HT2A receptors.
References
Appel JB, Callahan PM . (1989): Involvement of 5-HT receptor subtypes in the discriminative stimulus properties of mescaline. Eur J Pharmacol 159: 41–46
Bonhaus DW, Weinhardt KK, Taylor M, Desouza A, McNeeley PM, Szczepanski K, Fontan DJ, Trinh J, Rocha CL, Dawson MW, Flippin LA, Eglen RM . (1997): RS-102221: A novel high affinity and selective, 5-HT2C receptor antagonist. Neuropharmacology 36: 621–629.
Callahan PM, Cunningham KA . (1994): Involvement of 5-HT2C receptors in mediating the discriminative stimulus properties of m-chlorophenylpiperazine (mCPP). Eur J Pharmacol 257: 27–38
Dekeyne A, Girardon S, Millan MJ . (1999): Discriminative stimulus properties of the novel serotonin (5-HT)2C receptor agonist, RO60–0175: a pharmacological analysis. Neuropharmacology 38: 415–423
Fiorella D, Rabin RA, Winter JC . (1995): The role of 5-HT2A and 5-HT2C receptors in the stimulus effects of hallucinogenic drugs. I: Antagonist correlation analysis. Psychopharmacology 121: 347–356
Forbes IT, Jones GE, Murphy OE, Holland V, Baxter GS . (1995): N-(1-Methyl-5-indolyl)-N′-3-methyl-5-isothiazolyl)urea: a novel, high-affinity 5-HT2B receptor antagonist. J Med Chem 38: 855–857
Frazer A . (1997): Antidepressants. J Clin Psychiatry 58: 9–25
Friedman RL, Barrett RJ, Sanders-Bush E . (1984): Discriminative stimulus properties of quipazine: mediation by serotonin2 binding sites. J Pharmacol Exp Ther 228: 628–635
Glennon RA . (1991): Discriminative stimulus properties of hallucinogens and related designer drugs. In Glennon RA, Järbe TUC, Frankenheim J (eds), Drug Discrimination: Application to Drug Abuse Research. Rockville, Department of Health and Human Services Publication, pp 25–44
Glennon RA, Young R, Dukat M . (1992): 5-HT3 agonist, 2- methylserotonin, as a training drug in drug discrimination studies. Pharmacol Biochem Behav 41: 361–364
Gobert A, Rivet J-M, Lejeune F, Newman-Tancredi A, Adhumeau-Auclair A, Nicolas J-P, Cistarelli L, Melon C, Millan MJ . (2000): Serotonin2C receptors tonically suppress the activity of mesocortical dopaminergic and adrenergic, but not serotonergic, pathways: a combined dialysis and electrophysiological analysis in the rat. Synapse 36: 205–221
Gommans J, Hijzen TH, Maes RAA, Olivier B . (1998): Discriminative stimulus properties of mCPP: evidence for a 5-HT2C receptor mode of action. Psychopharmacology 137: 292–302
Kehne JH, Baron BM, Carr AA, Chaney SF, Elands J, Feldman DJ, Franck RA, Van Giersbergen PLM, McCloskey TC, Johnson MP, McCarty DR, Poirot M, Senyah Y, Siegel BW, Widmaier C . (1996): Preclinical characterization of the potential of the putative atypical antipsychotic MDL 100907 as a potent 5-HT2A antagonist with a favorable CNS safety profile. J Pharmacol Exp Ther 277: 968–981
Kennett GA, Wood MD, Bright F, Cilia J, Piper DC, Gager T, Thomas D, Baxter GS, Forbes IT, Ham P, Blackburn TP . (1996): In vitro and in vivo profile of SB 206553, a potent 5-HT2C/5-HT2B receptor antagonist with anxiolytic-like properties. Br J Pharmacol 117: 427–434
Kennett GA, Wood MD, Bright F, Trail B, Riley G, Holland V, Avenell KY, Stean T, Upton N, Bromidge S, Forbes IT, Brown AM, Midlemiss DN, Blackburn TP . (1997): SB 242084, a selective and brain penetrant 5-HT2C receptor antagonist. Neuropharmacology 36: 609–620
Millan MJ, Brocco M, Gobert A, Joly F, Bervoets K, Rivet J-M, Newman-Tancredi A, Audinot V, Morel S . (1999a): Contrasting mechanisms of action and sensitivity to antipsychotics of phencyclidine versus amphetamine: importance of nucleus accumbens 5-HT2A sites for PCP-induced locomotion in the rat. Eur J Neurosci 11: 4419–4432
Millan MJ, Gobert A, Girardon S, Dekeyne A . (1999b): Citalopram elicits a discriminative stimulus in rats at a dose selectively increasing extracellular levels of serotonin vs. dopamine and noradrenaline. Eur J Pharmacol 364: 147–150
Millan MJ, Peglion J-L, Lavielle G, Perrin-Monneyron S . (1997): 5-HT2C receptors mediate penile erections in rats: actions of novel and selective agonists and antagonists. Eur J Pharmacol 325: 9–12
Millan MJ, Schreiber R, Monneyron S, Denorme B, Melon C, Queriaux S, Dekeyne A . (1999c): S-16924, a novel, potential antipsychotic with marked serotonin1A agonist properties. IV. A drug discrimination comparison with clozapine. J Pharmacol Exp Ther 289: 427–436
Minnema DJ, Hendry JS, Rosecrans JA . (1984): Discriminative stimulus properties of pizotifen maleate (BC105): a putative serotonin antagonist. Psychopharmacology 83: 200–204
Palumbo PA, Winter JC . (1994): Interactions of clozapine with the stimulus effects of DOM and LSD. Pharmacol Biochem Behav 49: 115–120
Rinaldi-Carmona M, Congy C, Santucci V, Simiand J, Gautret B, Nelait G, Labeeuw B, Le Fur G, Soubrié P, Brelière JC . (1992): Biochemical and pharmacological properties of SR46349B, a new potent and selective 5-hydroxytryptamine2 receptor antagonist. J Pharmacol Exp Ther 262: 759–768
Roth BL, Meltzer HY . (1995): The role of serotonin in schizophrenia. In Bloom FE, Kupfer DJ (eds), Psychopharmacology: The Fourth Generation of Progress. New York, Raven Press, pp 1215–1228
Schreiber R, Brocco M, Audinot V, Gobert A, Veiga S, Millan MJ . (1995a): [1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane]-induced head-twitches in the rat are mediated by 5-hydroxytryptamine (5-HT)2A receptors: modulation by novel 5-HT2A/2C antagonists, D1 antagonists and 5-HT1A agonists. J Pharmacol Exp Ther 273: 101–112
Schreiber R, Brocco M, Lefèbvre de Ladonchamps B, Monneyron S, Millan MJ . (1995b): A drug discrimination analysis of the actions of novel serotonin1A receptor ligands in the rat using the 5-HT1A receptor agonist, 8-hydroxy-2-(di-n-propylamino)tetralin. J Pharmacol Exp Ther 275: 822–831
Schreiber R, Brocco M, Millan MJ . (1994): Blockade of the discriminative stimulus effects of DOI by MDL 100,907 and the ‘atypical’ antipsychotics, clozapine and risperidone. Eur J Pharmacol 264: 99–102
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dekeyne, A., Iob, L., Hautefaye, P. et al. The Selective Serotonin2A Receptor Antagonist, MDL100,907, Elicits a Specific Interoceptive Cue in Rats. Neuropsychopharmacol 26, 552–556 (2002). https://doi.org/10.1016/S0893-133X(01)00389-X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1016/S0893-133X(01)00389-X
Keywords
This article is cited by
-
Hallucinogen-like effects of 2-([2-(4-cyano-2,5-dimethoxyphenyl) ethylamino]methyl)phenol (25CN-NBOH), a novel N-benzylphenethylamine with 100-fold selectivity for 5-HT2A receptors, in mice
Psychopharmacology (2015)
-
Discriminative stimulus properties of the “atypical” antidepressant, mirtazapine, in rats: a pharmacological characterization
Psychopharmacology (2009)
-
Hallucinogens as discriminative stimuli in animals: LSD, phenethylamines, and tryptamines
Psychopharmacology (2009)
-
Contribution of serotonin (5-HT) 5-HT2 receptor subtypes to the discriminative stimulus effects of cocaine in rats
Psychopharmacology (2006)
