
Abstract
The neural crest is an embryonic stem cell population unique to vertebrates1 whose expansion and diversification are thought to have promoted vertebrate evolution by enabling emergence of new cell types and structures such as jaws and peripheral ganglia2. Although jawless vertebrates have sensory ganglia, convention has it that trunk sympathetic chain ganglia arose only in jawed vertebrates3,4,5,6,7,8. Here, by contrast, we report the presence of trunk sympathetic neurons in the sea lamprey, Petromyzon marinus, an extant jawless vertebrate. These neurons arise from sympathoblasts near the dorsal aorta that undergo noradrenergic specification through a transcriptional program homologous to that described in gnathostomes. Lamprey sympathoblasts populate the extracardiac space and extend along the length of the trunk in bilateral streams, expressing the catecholamine biosynthetic pathway enzymes tyrosine hydroxylase and dopamine β-hydroxylase. CM-DiI lineage tracing analysis further confirmed that these cells derive from the trunk neural crest. RNA sequencing of isolated ammocoete trunk sympathoblasts revealed gene profiles characteristic of sympathetic neuron function. Our findings challenge the prevailing dogma that posits that sympathetic ganglia are a gnathostome innovation, instead suggesting that a late-developing rudimentary sympathetic nervous system may have been characteristic of the earliest vertebrates.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
All raw RNA-sequencing data generated from this study are publicly available through the National Center for Biotechnology Information’s Gene Expression Omnibus database under the accession number GSE246248. RNA-sequencing data used for comparison to mouse sympathetic neurons were published previously41 and are available under the accession number GSE78845. Raw data can be made available upon request. Source data are provided with this paper.
References
Le Douarin, N. The Neural Crest (Cambridge Univ. Press, 1982).
Martik, M. L. et al. Evolution of the new head by gradual acquisition of neural crest regulatory circuits. Nature 574, 675–678 (2019).
Botar, J. Evolution and general anatomy of the autonomic nervous system. Gegenbaurs Morphol. Jahrb. 120, 271–279 (1974).
Burnstock, G. Evolution of the autonomic innervation of visceral and cardiovascular systems in vertebrates. Pharmacol. Rev. 21, 247–324 (1969).
Haming, D. et al. Expression of sympathetic nervous system genes in lamprey suggests their recruitment for specification of a new vertebrate feature. PLoS ONE 6, e26543 (2011).
Johnels, A. G. On the peripheral autonomic nervous system of the trunk region of Lampetra planeri. Acta Zool. 37, 251–286 (1956).
Nicol, J. C. Autonomic nervous systems in lower chordates. Biol. Rev. 27, 1–48 (1952).
Rovainen, C. M. Neurobiology of lampreys. Physiol. Rev. 59, 1007–1077 (1979).
Shu, D. G. et al. Head and backbone of the Early Cambrian vertebrate Haikouichthys. Nature 421, 526–529 (2003).
Green, S. A., Simoes-Costa, M. & Bronner, M. E. Evolution of vertebrates as viewed from the crest. Nature 520, 474–482 (2015).
Green, S. A. & Bronner, M. E. The lamprey: a jawless vertebrate model system for examining origin of the neural crest and other vertebrate traits. Differentiation 87, 44–51 (2014).
Anderson, D. J. & Axel, R. A bipotential neuroendocrine precursor whose choice of cell fate is determined by NGF and glucocorticoids. Cell 47, 1079–1090 (1986).
Anderson, D. J., Carnahan, J. F., Michelsohn, A. & Patterson, P. H. Antibody markers identify a common progenitor to sympathetic neurons and chromaffin cells in vivo and reveal the timing of commitment to neuronal differentiation in the sympathoadrenal lineage. J. Neurosci. 11, 3507–3519 (1991).
Shtukmaster, S. et al. Sympathetic neurons and chromaffin cells share a common progenitor in the neural crest in vivo. Neural Dev. 8, 12 (2013).
Le Douarin, N. M. & Smith, J. Development of the peripheral nervous system from the neural crest. Annu. Rev. Cell Biol. 4, 375–404 (1988).
Loring, J. F. & Erickson, C. A. Neural crest cell migratory pathways in the trunk of the chick embryo. Dev. Biol. 121, 220–236 (1987).
Reissmann, E. et al. Involvement of bone morphogenetic protein-4 and bone morphogenetic protein-7 in the differentiation of the adrenergic phenotype in developing sympathetic neurons. Development 122, 2079–2088 (1996).
Schneider, C., Wicht, H., Enderich, J., Wegner, M. & Rohrer, H. Bone morphogenetic proteins are required in vivo for the generation of sympathetic neurons. Neuron 24, 861–870 (1999).
Ernsberger, U., Reissmann, E., Mason, I. & Rohrer, H. The expression of dopamine beta-hydroxylase, tyrosine hydroxylase, and Phox2 transcription factors in sympathetic neurons: evidence for common regulation during noradrenergic induction and diverging regulation later in development. Mech. Dev. 92, 169–177 (2000).
Furlan, A. et al. Multipotent peripheral glial cells generate neuroendocrine cells of the adrenal medulla. Science 357, eaal3753 (2017).
Kastriti, M. E., Kameneva, P. & Adameyko, I. Stem cells, evolutionary aspects and pathology of the adrenal medulla: A new developmental paradigm. Mol. Cell. Endocrinol. 518, 110998 (2020).
Augustinsson, K. B., Fange, R., Johnels, A. & Ostlund, E. Histological, physiological and biochemical studies on the heart of two cyclostomes, hagfish (Myxine) and lamprey (Lampetra). J. Physiol. 131, 257–276 (1956).
Paiement, J. M. & McMillan, D. B. The extracardiac chromaffin cells of larval lampreys. Gen. Comp. Endocrinol. 27, 495–508 (1975).
Butler, D. G. Structure and function of the adrenal gland of fishes. Am. Zool. 13, 839–879 (1973).
Reid, S. G., Bernier, N. J. & Perry, S. F. The adrenergic stress response in fish: control of catecholamine storage and release. Comp. Biochem. Physiol. C 120, 1–27 (1998).
Kirby, M. & Gilmore, S. A correlative histofluorescence and light microscopic study of the formation of the sympathetic trunks in chick embryos. Anat. Rec. 186, 437–449 (1976).
Kastriti, M. E. et al. Schwann cell Precursors generate the majority of chromaffin cells in Zuckerkandl organ and some sympathetic neurons in paraganglia. Front. Mol. Neurosci. 12, 6 (2019).
Thomsen, E. R. et al. Fixed single-cell transcriptomic characterization of human radial glial diversity. Nat. Methods 13, 87–93 (2016).
Ernsberger, U., Kramer, M., Tsarovina, K., Deller, T. & Rohrer, H. Coordinate expression of pan-neuronal and functional signature genes in sympathetic neurons. Cell Tissue Res. 370, 227–241 (2017).
Friedman, S. & Kaufman, S. 3,4-Dihydroxyphenylethylamine beta-hydroxylase: a copper protein. J. Biol. Chem. 240, PC552–PC554 (1965).
Beesley, P. W., Herrera-Molina, R., Smalla, K. H. & Seidenbecher, C. The neuroplastin adhesion molecules: key regulators of neuronal plasticity and synaptic function. J. Neurochem. 131, 268–283 (2014).
Poea-Guyon, S. et al. The V-ATPase membrane domain is a sensor of granular pH that controls the exocytotic machinery. J. Cell Biol. 203, 283–298 (2013).
Coughlin, M. D., Boyer, D. M. & Black, I. B. Embryologic development of a mouse sympathetic ganglion in vivo and in vitro. Proc. Natl Acad. Sci. USA 74, 3438–3442 (1977).
Kuntz, A. The development of the sympathetic nervous system in man. J. Comp. Neurol. 32, 173–229 (1920).
An, M., Luo, R. & Henion, P. D. Differentiation and maturation of zebrafish dorsal root and sympathetic ganglion neurons. J. Comp. Neurol. 446, 267–275 (2002).
Young, J. Z. Memoirs: The autonomic nervous system of Selachians. J. Cell Sci. 2, 571–624 (1933).
Nicol, J. A. The autonomic nervous system of the chimaeroid fish Hydrolagus colliei. Q. J. Microsc. Sci. 91, 379–399 (1950).
Colin Nicol, J. A. Autonomic nervous system of the ratfish. Nature 165, 854 (1950).
Romer, A. S. in Evolutionary Biology (eds Dobzhansky, T., Hecht, M.K. & Steere, W.C.) 121–156 (Springer, 1972).
Young, J. Z. Memoirs: On the autonomic nervous system of the Teleostean fish Uranoscopus scaber. J. Cell Sci. 2, 491–536 (1931).
Furlan, A. et al. Visceral motor neuron diversity delineates a cellular basis for nipple- and pilo-erection muscle control. Nat. Neurosci. 19, 1331–1340 (2016).
Han, S. et al. Direct evidence for the role of neuropeptide Y in sympathetic nerve stimulation-induced vasoconstriction. Am. J. Physiol. 274, H290–H294 (1998).
Brazeau, M. D. & Friedman, M. The origin and early phylogenetic history of jawed vertebrates. Nature 520, 490–497 (2015).
Gans, C. & Northcutt, R. G. Neural crest and the origin of vertebrates: a new head. Science 220, 268–273 (1983).
Eklov, P. & Svanback, R. Predation risk influences adaptive morphological variation in fish populations. Am. Nat. 167, 440–452 (2006).
Vinterstare, J. et al. Predation risk and the evolution of a vertebrate stress response: parallel evolution of stress reactivity and sexual dimorphism. J. Evol. Biol. 34, 1554–1567 (2021).
Charmandari, E., Tsigos, C. & Chrousos, G. Endocrinology of the stress response. Annu. Rev. Physiol. 67, 259–284 (2005).
Pierre, J., Mahouche, M., Suderevskaya, E. I., Reperant, J. & Ward, R. Immunocytochemical localization of dopamine and its synthetic enzymes in the central nervous system of the lamprey Lampetra fluviatilis. J. Comp. Neurol. 380, 119–135 (1997).
Chen, E. Y. et al. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 14, 128 (2013).
Kuleshov, M. V. et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 44, W90–W97 (2016).
Xie, Z. et al. Gene set knowledge discovery with Enrichr. Curr. Protoc. 1, e90 (2021).
Nikitina, N., Bronner-Fraser, M. & Sauka-Spengler, T. Culturing lamprey embryos. Cold Spring Harb. Protoc. 2009, prot5122 (2009).
Tahara, Y. Normal stages of development in the lamprey, Lampetra reissued (Dybowski). Zool. Sci. 5, 109–118 (1988).
Choi, H. M. et al. Programmable in situ amplification for multiplexed imaging of mRNA expression. Nat. Biotechnol. 28, 1208–1212 (2010).
Nikitina, N., Bronner-Fraser, M. & Sauka-Spengler, T. DiI cell labeling in lamprey embryos. Cold Spring Harb. Protoc. 2009, prot5124 (2009).
Kim, K. M., Son, K. & Palmore, G. T. Neuron Image Analyzer: automated and accurate extraction of neuronal data from low quality images. Sci. Rep. 5, 17062 (2015).
Hempel, C. M., Sugino, K. & Nelson, S. B. A manual method for the purification of fluorescently labeled neurons from the mammalian brain. Nat. Protoc. 2, 2924–2929 (2007).
Bray, N. L., Pimentel, H., Melsted, P. & Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 34, 525–527 (2016).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Acknowledgements
We thank A. Collazo and G. Spigolon for microscopy training and assistance; I. Antoshechkin for discussion and assistance with library preparation and sequencing; G. Shin and N. Pierce for discussion and assistance with HCR probe design; and J. Stundlova, R. Fraser and D. Mayorga for lamprey husbandry. This work was supported by the grants NIH R35NS111564 to M.E.B., NIH F32HD106627 to B.M.E. and NIH F31DE031154 to H.A.U. J.S. is supported by a Marie Skłodowska-Curie grant, agreement no. 897949.
Author information
Authors and Affiliations
Contributions
The project was conceived of and designed by B.M.E. and M.E.B. Descriptive and transcriptomic analyses were carried out by B.M.E. CM-DiI labelling was carried out by J.S. and H.A.U. Preparation and sectioning of all T21 and T30 CM-DiI-labelled embryos was carried out by J.S. Lamprey husbandry and spawning were led by J.S. Writing and interpretation were carried out by B.M.E. and M.E.B. All authors approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Uwe Ernsberger, Andrew Gillis, Takayuki Onai and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
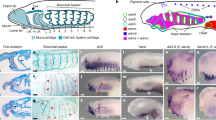
Extended Data Fig. 1 Expression of sympathoadrenal genes at early embryonic stages.
(a-c) HCR detection of Phox2 (teal), Hand (red), and Ascl1 (white) in lamprey at T25. (d) The three transcripts are not co-expressed. Scale bar = 50 μm. n = 6 embryos across three independent replicates.
Extended Data Fig. 2 Co-expression of sympathoadrenal fate-specifying genes with catecholamine biosynthetic enzymes in post-embryonic lamprey.
HCR detection of Th (teal) and Dbh (red) with (a-d) Ascl1, (e-h) Phox2, and (i-l) Hand in white at T30+ in whole mount. The core sympathoadrenal transcription factors are persistently co-expressed with catecholamine synthesis enzymes. Scale bars = 20 μm. n = 9 embryos across three independent replicates.
Extended Data Fig. 3 Early expression of catecholamine biosynthetic enzymes.
(a-d) HCR detection of Th (teal) and Dbh (red) in lamprey at T27. Colocalization of the transcripts is seen in cells surrounding the heart and spanning the initial segment of the trunk in bilateral streams. Scale bars = 100 μm. Red asterisk denotes the heart. n = 6 embryos across three independent replicates.
Extended Data Fig. 4 Quantification of relative morphology of TH+ neuroblasts/neurons from T27 to T30.
Morphology of TH+ cells was compared from T27- and T30- staged embryos by plotting [neurite length/soma diameter]. This measure revealed increased neurite length in T30 as compared to T27, consistent with greater neuronal maturation. **** p < 0.0001 by two-tailed Mann-Whitney U-test. A total of fifty cells from five embryos were measured for each developmental stage. Error bars indicate SEM.
Extended Data Fig. 5 Late neurogenesis of lamprey sympathoblasts.
(a-i) Immunohistochemical detection of TH (teal) and Neurofilament-M (Nf-M, red) at T30 (a-c), 2.5 month ammocete (d-f), and 4 month ammocete (g-i) stages. Co-expression of TH and Nf-M is not observed in late embryonic or earlier ammocete stages. Consistent co-labeling is observed by four months of age. n = 12 embryos, 12 ammocetes of each stage per marker across four independent replicates (j-o) Co-expression of TH with additional neuronal markers is observed in four month old ammocetes. HuC/D (j-l) and SCG10 (m-o) are shown. DAPI is shown in white. Scale bars = 50 μm. n = 9 ammocetes of each stage per marker across three independent replicates.
Extended Data Fig. 6 Lamprey sympathoblasts are neural crest-derived.
Lineage tracing of TH+ sympathoblasts. CM-DiI-labeled neural crest (red) localizes to TH+ cells (teal) at (a-c) T27 and (d, e) T30, indicating a neural crest origin. Arrowheads (b, c, and e) indicate CM-DiI-labeled cells. DAPI is shown in white. Scale bars = 20 μm. DA=dorsal aorta, E=esophagus, I=Intestine. Experimental details are shown in Extended Data Table 1.
Extended Data Fig. 7 Analysis of gene expression in lamprey sympathetic neurons.
(a) Representative image of a wild-caught lamprey ammocete used for selection of trunk sympathetic neurons. Scale bar = 1 cm. (b and c) Immunohistochemical detection of TH (teal) in transverse sections through ammocete trunks. DAPI is shown in white. Yellow box indicates the site of the sympathetic chain neurons. SC=spinal cord, NC=notochord. Scale bars = 250 μm (a) and 10 μm (b). n = 4 ammocetes across three independent replicates. (d and e) GO analyses of Metabolites and Cell Compartment for TH+ neurons. Analyses are performed using the top 500 significantly upregulated transcripts with an enrichment cutoff of 1.5 log2(Fold Change). p-values are determined using Fisher’s exact test. (f) Immunohistochemical detection of DBH (teal) at T28. Punctate expression is consistent with vesicular localization of DBH protein. Scale bar = 20 μm. n = 6 embryos across three independent replicates.
Extended Data Fig. 8 Comparison of gene expression between mouse and lamprey sympathetic neurons.
(a) GO analysis of Enrichr Protein-Protein Interaction (PPI) Hubs49 for mouse sympathetic neurons previously reported41. Analysis is performed using the 500 most abundantly expressed genes across all noradrenergic neuron subtypes. p-values are determined using Fisher’s exact test. (b) Venn diagram indicating PPI Hub terms implicated in both mouse and lamprey sympathetic neurons.
Extended Data Fig. 9 Analysis of noradrenergic subtypes in lamprey.
HCR detection of Th (teal) with (a-c) Npy (red), (d-g) Npy (red) and Ret (white), and (h-k) Gfra2 (red) and Ret (white) in whole mount ammocete trunks. Consistent co-expression of Th and Npy (a-c) suggests the presence of the NA3 noradrenergic subtype in lamprey. A lack of consistent co-expression of Th with Npy and Ret (d-g) and Gfra2 and Ret suggest the absence of noradrenergic subtypes NA2 and NA5, respectively. Scale bars = 20 μm. n = 12 ammocetes across four independent replicates.
Supplementary information
Source data
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Edens, B.M., Stundl, J., Urrutia, H.A. et al. Neural crest origin of sympathetic neurons at the dawn of vertebrates. Nature 629, 121–126 (2024). https://doi.org/10.1038/s41586-024-07297-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-024-07297-0
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
