
Abstract
Circulating microRNAs (miRNAs) are promising disease biomarkers. However, the influence of high-altitude hypoxic environments on plasma miRNA profiles remains unknown. This study included a total of 509 plasma samples from 278 native Tibetans and 80 newly arrived migrant Han Chinese (Tibet Han) residing at 3560 m and 151 Han Chinese residing at 8.9 m (Nanjing Han). The levels of 754 miRNAs were initially determined using a TaqMan Low Density Array (TLDA) in two pooled samples from 50 Tibet Han and 50 Nanjing Han individuals. Some markedly altered miRNAs in Tibet Han were subsequently measured in all 509 plasma samples by individual qRT-PCR. Compared with the Nanjing Han, 172 miRNAs were differentially expressed in the Tibet Han (105 upregulated and 67 downregulated). The correlation coefficient for the two groups was 0.72. Several upregulated miRNAs were randomly selected for analysis by qRT-PCR and the results were consistent with those identified by TLDA. These miRNAs were also significantly increased in the Tibetans compared with the Nanjing Han. Furthermore, these altered miRNAs showed strong positive correlations with red blood cell counts and hemoglobin values. These data are the first to provide clear evidence that a high-altitude hypoxic environment significantly affects human plasma miRNA profiles.
Similar content being viewed by others

Introduction
MicroRNAs (miRNAs) are a class of small non-coding RNAs of ~22 nucleotides in length that regulate gene expression at the posttranscriptional level by either degrading or blocking the translation of messenger RNA targets. The involvement of miRNAs in a variety of physiological and pathological conditions has been demonstrated1,2. Recent studies by our group and others have shown that circulating miRNAs are stably detectable and may serve as potential biomarkers for cancer and non-cancer diseases3,4,5,6,7. Therefore, circulating miRNA expression profiles are being intensively investigated for their involvement in various pathogenic processes. To translate miRNA assays from the bench side to the bedside, the pre-assay conditions of human circulating miRNA profiles must be determined. Circulating miRNA concentrations can be affected by some confounding factors, such as age and sex8. However, the influence of environmental factors, such as living conditions, residence and altitude, have not been determined.
Among the most severe environmental challenges to confront human is the reduced oxygen availability in high-altitude regions, such as in the Tibetan Plateau9. The Tibetan Plateau is the highest plateau in the world with an average altitude of over 4500 m and it is inhospitable to human settlement because of cold temperatures and low atmospheric oxygen pressure (~40% lower than at sea level)10. When lowlanders immigrate to high-altitude locations, they are not well adapted to the high-altitude hypoxic conditions. For most individuals, acclimatization to low oxygen involves a marked increase in hemoglobin (HGB) levels10. Recently, a few reports revealed that hypoxia was an important proximal regulator of miRNA biogenesis and function and that miRNAs could rapidly respond to the stress caused by hypoxia by post-transcriptional translational modulation at the cellular level11,12,13. Therefore, it is reasonable to speculate that high-altitude hypoxic environments are likely to have an impact on circulating miRNA concentrations.
To test our hypothesis, in the present study, we performed high-throughput TaqMan Low Density Array (TLDA) screening followed by confirmation using a quantitative reverse-transcription polymerase chain reaction (qRT-PCR) assay to systematically and comprehensively evaluate and compare plasma miRNA profiles between Han Chinese residing in Nanjing at sea level (Nanjing Han) and Han Chinese recently immigrated from the plains to Tibet (Tibet Han) as well as ethnic Tibetans. We also analysed the correlations between the altered plasma miRNAs and hematological indices.
Results
TLDA chip analysis of plasma miRNA profiles
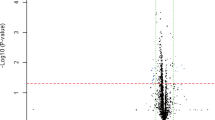
First, we globally analysed the plasma miRNA expression profiles from the Tibet Han and Nanjing Han samples using a TLDA chip. Fifty plasma samples randomly selected from 80 Tibet Han people and 151 Nanjing Han people were separately pooled to form two sample pools for the TLDA analysis. The mean age and sex distribution of the 50 Nanjing Han were exactly matched with those of the 50 Tibet Han (34.49 ± 9.17 years, 28 male/22 female vs. 35.38 ± 11.69 years, 32 male/18 female; P = 0.765 and P = 0.414, respectively). Pearson correlation analysis showed the correlation coefficient (r) for the two groups to be 0.72 (Fig. 1). The miRNAs assessed were considered to be upregulated if their quantification cycle (Cq) values were <35 in the Tibet Han sample and downregulated if their Cq values were <35 in the Nanjing Han sample and if there was a 2-fold difference in the expression level between the two groups. Of the 754 miRNAs tested, 172 miRNAs were differently expressed in the following manner: 105 miRNAs were upregulated and 67 miRNAs downregulated in the Tibet Han group compared with the Nanjing Han group (Supplementary Table S1). Forty-five of the markedly altered miRNAs from the Tibet Han group (change fold >100) are listed in Table 1. The TLDA results demonstrated that the plasma miRNA patterns in the Tibet Han group were markedly different from those in the Nanjing Han group.

Pearson correlation scatter plot of plasma miRNA levels in the Tibet Han and Nanjing Han groups as determined by TLDA.
We used the miRBase database (version 22.0) to analyse whether the markedly upregulated or downregulated miRNAs in Table 1 are part of families residing in the same genomic location. miRNA gene clusters were named based on their close physical distances to genes on the same chromosome (<10 kb). We found that miR-495 and miR-323a-3p, miR-500a-5p and miR-501-5p, miR-518e-3p and miR-520d-3p and miR-516a-3p and miR-522-3p are each a set of two members of four miRNA clusters and each pair is likely to be co-regulated.
Confirmation of the TLDA results using individual qRT-PCR assays
Next, to confirm the TLDA results, we carried out qRT-PCR assays of miRNAs with individual plasma samples. Four plasma miRNAs that were upregulated in the Tibet Han group compared with the Nanjing Han group upon TLDA analysis (Fig. 1 and Supplementary Table S1); specifically, miR-130a-3p, miR-302b-5p, miR-572 and miR-629-5p, were randomly selected for qRT-PCR validation with individual plasma samples from 80 Tibet Han and 151 Nanjing Han persons. In addition, these 4 miRNAs were also examined in the plasma samples from 278 ethnic Tibetan highlanders. Table 2 summarizes the demographic and hematological features of all participants. No significant differences in age or sex were identified among the Tibet Han, Nanjing Han and Tibetan groups. However, the red blood cell counts (RBC), HGB, hematocrit (HCT) and mean corpuscular hemoglobin (MCH) values of both males and females were significantly increased (P < 0.005), whereas blood platelet (PLT) levels were markedly decreased (P < 0.001) in the Tibet Han group compared with the Nanjing Han group. The HGB, HCT and MCH values of both males and females, along with the female RBC values, were also significantly increased in the Tibetan group compared with the Nanjing Han group, whereas PLT levels were markedly decreased between the two groups (P < 0.001); however, no marked difference in male RBC values was observed between the two groups. In addition, the male RBC, HGB and HCT values were significantly increased in the Tibet Han group compared with those of the Tibetan group (P < 0.001).
The results of the individual qRT-PCR assays were in agreement with those of the TLDA analysis of the pooled samples showing that the plasma concentrations of all 4 of the selected miRNAs were significantly increased in the Tibet Han group compared with the Nanjing Han group (fold change >2.0 and P < 0.0001) (Fig. 2 and Supplementary Table S2). Furthermore, the levels of the 4 miRNAs were also markedly higher in the Tibetan group than in the Nanjing Han group (P < 0.0001) (Fig. 2 and Supplementary Table S2). However, the Tibet Han and Tibetan groups were not significantly different with respect to the concentrations of the 4 miRNAs with the exception of miR-302b-5p, which was higher in the Tibet Han group than in the Tibetan group (P = 0.016).

The relative concentrations of miR-130a-3p, miR-302b-5p, miR-572 and miR-629-5p in the plasma samples from the Nanjing Han (n = 151), Tibet Han (n = 80) and Tibetan (n = 278) groups (A–D).
Cq values were converted to relative concentrations normalized to MIR2911 values and were calculated using the comparative Cq method (2−ΔCq). Each point represents the mean of triplicate samples. *P < 0.05; ***P < 0.0001.
Expression levels of the validated upregulated miRNAs in peripheral blood cells
We measured the levels of the 4 validated upregulated miRNAs in peripheral blood cell samples of 30 Nanjing Han, 30 Tibet Han and 30 Tibetan individuals. The results showed that the expression levels of these 4 miRNAs were significantly higher in the Tibet Han and Tibetan groups than in the Nanjing Han group (P < 0.05), whereas no marked difference in these miRNAs was observed between the Tibet Han and Tibetan groups (Supplementary Fig. S3).
The co-correlation of the validated hypoxia miRNAs and their relationships with hematological indices
We used a Spearman rank correlation analysis to test for the co-correlation between the 4 validated increased plasma miRNAs in all studied individuals. As shown in Supplementary Fig. S4, a strong positive correlation was observed between the 4 miRNAs (P < 0.0001). These results indicate that some individuals have higher miRNA responses overall. Furthermore, we evaluated whether these 4 miRNAs were related to hematological parameters. We used Spearman rank correlation analysis to test for correlations between each of these 4 miRNAs and the measured hematological parameters. All 4 miRNAs showed strong positive correlations with RBC, HGB and HCT (P < 0.01) and significantly negative relationships with PLT (P < 0.05) (Table 3 and Supplementary Fig. S5–Fig. S7). Together with the qRT-PCR results, these data suggest that the upregulation of these 4 miRNAs in populations residing in Tibet may be a result of the high-altitude hypoxic environment.
Target analysis of miRNAs altered by high-altitude hypoxia
To explore the possible roles and molecular basis of the high-altitude-altered miRNAs in response to a hypoxic environment, we performed a bioinformatics prediction of the potential target genes associated with hypoxia through the use of several widely utilized target prediction databases, including TargetScan, miRanda and PicTar. Computational analysis revealed that some hypoxia-related genes, such as DDX6, hypoxia-inducible factors-3 (HIF-3), vascular endothelial growth factor-A (VEGFA) and egl nine homologue 1 (EGLN1), were potential target genes of the validated miRNAs (Table 4). In addition, several erythroid-related and megakaryocytic-related genes, such as SMAD5 and ETV6, were also predicted to be potential targets of miR-130a-3p and miR-302b-5p (Table 4).
Subsequently, we measured the plasma concentrations of the growth factors VEGFA and erythropoietin (EPO) in the plasma samples from 64 Nanjing Han, 46 Tibet Han and 64 Tibetan individuals using an enzyme-linked immunosorbent assay (ELISA). The plasma concentrations of both VEGFA and EPO were found to be significantly elevated in the Tibet Han (P < 0.05 and P < 0.001, respectively) and Tibetan groups (P = 0.001 and P < 0.001, respectively) compared with the Nanjing Han group; however, no marked difference in the two growth factors was observed between the Tibet Han and Tibetan groups (P > 0.05) (Supplementary Table S3). Furthermore, we evaluated the associations between VEGFA, EPO and the expression levels of the 4 validated hypoxia miRNAs using Spearman rank correlation analysis. The plasma concentrations of both VEGFA and EPO exhibited strong positive correlations with the expression levels of the 4 miRNAs (Supplementary Fig. S8 and Fig. S9).
Discussion
miRNAs have been increasingly recognized as biomarkers for diverse classes of disease; however, the influences of pre-assay conditions, such as the effects of environmental factors on human circulating miRNA profiles, are largely unknown. In this study, we investigated for the first time the influence of a high-altitude hypoxic environment on human plasma miRNA profiles. By performing a genome-wide plasma miRNA differentiation scan using a TLDA chip followed by qRT-PCR validation, we demonstrated that the plasma miRNA profile of Han Chinese who had recently immigrated to highland Tibet is dramatically different from that of those residing at sea level. More importantly, the upregulated miRNAs showed strong positive correlations with RBC, HGB and HCT values. These results suggest that a high-altitude hypoxic environment has a remarkable influence on human plasma miRNA patterns.
The main physiological challenge in high-altitude plateau environments is hypoxia. When people living in a plain environment migrate to a plateau, they face the threat of hypoxia14. Physiological responses to high-altitude hypoxia are complex and involve a range of mechanisms15. Previous studies have discovered a special group of miRNAs that are differentially expressed in response to hypoxia and altered miRNA concentration profiles, or signatures, from various organisms, cell types and disease states in relation to hypoxia have been determined16,17,18. However, these studies mainly focused on miRNA expression levels in tissues or cells. Considering that miRNAs are key elements in the response to hypoxia, we speculated that high-altitude hypoxic environments are likely to have an impact on human circulating miRNA patterns. To test our hypothesis, we used a high-throughput TLDA assay to comparatively analyse miRNA expression profiles of the Tibet Han and Nanjing Han populations. As expected, the expression profiles of plasma miRNAs from the Tibet Han were dramatically different from those of the Nanjing Han, with 172 differently expressed miRNAs. Next, we used qRT-PCR assays to confirm the TLDA results. We randomly selected 4 miRNAs to measure their concentrations by qRT-PCR in individual plasma samples in a large validation cohort. Consequently, the results of the individual qRT-PCR assays were in accordance with those of the TLDA analysis showing that the plasma concentrations of these selected miRNAs were significantly increased in the Tibet Han group compared with the Nanjing Han group. To confirm that the miRNA dysregulation observed in the Tibet Han was the result of a high-altitude hypoxic environment, we also examined these markedly altered miRNAs in native Tibetan highlanders. We found that the levels of all 4 miRNAs were also markedly higher in the Tibetan highlanders than in the Han lowlanders, whereas most of these miRNAs did not show any significant difference between the native Tibetan and Tibet Han individuals. Furthermore, we observed a similar trend of these miRNAs in the peripheral blood cells of the studied individuals, suggesting that these plasma miRNA levels may reflect cellular changes in the populations investigated. These results demonstrate that a number of miRNAs were differentially expressed in the circulation in response to a high-altitude hypoxic environment. In addition, these hypoxia-associated miRNAs exhibited strong positive correlations, indicating that some individuals have a higher overall miRNA response when exposed to high altitude.
Hypoxia, or low oxygen availability, has a complex and extensive impact on human and animal physiology and elaborate adaptive mechanisms have evolved to respond to hypoxic stress. Of the 4 high-altitude hypoxia-upregulated plasma miRNAs identified in our study, miR-130a-3p and miR-572 have been reported to be upregulated in cells under hypoxic conditions19,20. In addition, a subset of the altered miRNAs (e.g., miR-21, miR-24, miR-30b, miR-224) identified during our TLDA analysis has also been associated with the hypoxic response in mammalian cells by other investigators17,21. These miRNAs may be regarded as general responders to hypoxia. Functional studies of miRNAs that respond during hypoxia may be helpful in uncovering the molecular basis of hypoxic acclimatization and illuminating the complexity of the hypoxia-response pathways in humans. Interestingly, by analysing the resulting data sets based on multiple target prediction algorithms, we found that some target genes of our identified miRNAs are involved in oxygen sensing and metabolism. For instance, DDX6, a DEAD-box helicase, was predicted to be the target gene of miR-130a. Decreased expression of DDX6 by miR-130a has been found to enhance the translation of hypoxia-inducible factor-1alpha (HIF-1α), a key transcription factor in the cellular response to hypoxia, in an internal ribosome entry site19. HIF-3, another oxygen-dependent transcription factor that activates a distinct transcriptional response to hypoxia22, was predicted to be a target gene of miR-629-5p. In addition, some genes, including EGLN1 and peroxisome proliferator-activated receptor alpha (PPARA), which are involved in the HIF pathway, were potential target genes of miR-302b-5p. EGLN1 plays a central role in the activation of hypoxia-inducible genes and the homeostasis of HIF under hypoxia and normoxia23,24. EGLN1 targets two HIFα proteins for degradation under normoxic conditions, decreasing the transcription of HIF-regulated targets, such as EPO, the erythropoietin gene whose product induces RBC production9. PPARA is also associated with HIF activity. PPARA expression is repressed by HIF1 during hypoxia9,25 and genes targeted by HIF are regulated by a HIF-independent mechanism involving PPARG coactivator-1α26.
In our study, we found that the plasma concentrations of the growth factors VEGFA and EPO exhibited strong positive correlations with the expression levels of the validated hypoxia-associated miRNAs. VEGFA was predicted to be a potential target of miR-302b-5p by bioinformatics analysis. The exact reason for the positive relationship between miR-302b-5p and VEGFA is currently unknown. Nevertheless, a recent study has demonstrated that hypoxia increases VEGFA expression in a number of epithelial cells27. Interestingly, our results also revealed that hypoxia could induce miR-302b-5p expression in peripheral blood cells. This evidence indicates that elevated VEGFA and miR-302b-5p levels may originate from different cells under hypoxic conditions and can explain why they show a strong positive correlation even though VEGFA is a potential target of miR-302b-5p. However, further studies are necessary both to clarify this issue and to validate the target genes of these miRNAs and the mechanisms by which circulating miRNAs respond to hypoxia.
In this study, we also tested the association of the validated upregulated miRNAs in the Tibet Han with several hematological indices that had distinctively high levels in the newly arrived migrant individuals originating from low-altitude populations. The specialized function of red blood cells is to transport O2 from pulmonary capillaries to tissue capillaries, where it is exchanged for CO2. When lowlanders immigrate to high altitude locations, they are not well adapted to the high-altitude hypoxic conditions and for most individuals, acclimatization to low oxygen involves rapid increases in blood RBC, HGB and HCT levels9,10,28. In our study, we observed that RBC, HGB and HCT levels were significantly higher in both the Tibet Han and Tibetan groups than in the lowlander Han group and these hematological indices were higher in the Tibet Han group than in the Tibetan group, which was consistent with previous reports29,30. It is worth mentioning that, in addition to these hematological indices, the levels of miR-302b-5p, a high-altitude hypoxia-upregulated miRNA validated in our study, was also higher in the Tibet Han than in the native Tibet population. The reason for this phenomenon may be that the physiological response to low oxygen of individuals of low-altitude origin is different than that of native Tibetans highlanders. Tibetans have lived at very high altitudes for thousands of years and they have evolved a blunted physiological response to high-altitude hypoxia9. Furthermore, we used Spearman rank correlation analysis to test for correlations between the 4 plasma miRNAs and the hematological indices. We found that all 4 of the miRNAs showed strong and significant positive correlations with RBC, HGB and HCT values. These data provide evidence for the expression/activation of an array of circulating miRNAs in a highly coordinated manner with other physiological responses, such as increased hematological responses, to achieve enhanced cell survival under hypoxic conditions. In addition, target prediction revealed that some genes with important roles in erythroid and megakaryocytic differentiation were potential targets of our identified miRNAs. For example, genes related to the erythropoietin, such as SMAD531 and SOX632 and transcription factors with well-known functions in megakaryocytopoiesis, such as MAFB33 and ETV634, are putative targets for miR-130a-3p and miR-302b-5p, respectively. These results indicate that miR-130a-3p and miR-302b-5p could be involved in regulating erythroid and megakaryocytic differentiation and are likely involved in high-altitude acclimatization and/or adaptation. Further study regarding the alteration and the roles of circulating miRNAs in physiological responses may shed light on the connections between these miRNAs and altitude acclamation.
In a previous study, it was observed that with increasing altitude, men had progressively more HGB than women in the Han but not the Tibetan population, indicating that a gender-related difference was greater in Han than in Tibetan individuals. Consequently, the difference in HGB values between the Tibet Han men and Tibetan men was larger than the Tibet Han women and Tibetan women29. A similar phenomenon was found in our study. The reasons underlying the progressively higher HGB in Han men compared with Han women as altitude increases are unknown. One possible reason, we suspect, is that Han men may have a stronger chronic erythropoietic response to altitude than their female counterparts. Further study is needed to clarify this issue.
In conclusion, we have demonstrated for the first time that a high-altitude hypoxic environment has a marked effect on human plasma miRNA expression profiles. In particular, we have identified a set of high-altitude hypoxia-altered plasma miRNAs that show strong positive correlations with hematological phenotypes. These data provide new insight into the physiological traits related to hypoxia and the molecular mechanisms involved in the hypoxic response, which has important implications for the prevention and treatment of some hypoxia-related diseases, such as mountain sickness and high-altitude pulmonary disease.
Materials and Methods
Participants and plasma sample processing
The study population consisted of a total of 509 plasma samples from 80 ethnic Han Chinese who immigrated one to two years ago (mean 17 months) from the plains (Jiangsu Province, Anhui Province and Shanghai City of China) to Nêdong County, a county of Lhoka prefecture in the Tibet Autonomous Region with an altitude of 3560 m (Tibet Han); 151 Han Chinese residing in Nanjing (Nanjing Han), a city of Jiangsu Province, with an altitude of 8.9 m; and 278 native Tibetans residing in Nêdong since birth. The Tibet Han and Tibetan study participants were recruited from individuals seeking routine health check-ups at the Forty-First Hospital of PLA in Nêdong, China and those in the Nanjing Han group were recruited from individuals who had visited the Jinling Hospital, Nanjing, China, for routine check-ups between 2013 and 2014. Jiangsu, Anhui and Shanghai are all in the east of China and close to each other. The genetic backgrounds of the Tibet Han were matched with the Nanjing Han35.
All blood samples were collected in EDTA tubes using a standard operating procedure. All blood donors participating in this study provided written informed consent and the ethics committees of the Forty-First Hospital of PLA and Jinling Hospital approved the study protocol in accordance with the Declaration of Helsinki.
RNA isolation, TLDA and qRT-PCR
We carried out RNA isolation and measurements as previously described5. In brief, blood was centrifuged at 3000 rpm for 10 min and the plasma supernatant was then collected and stored at −80 °C until RNA extraction. For TLDA, RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) from two pooled samples from 50 Tibet Han people or 50 Nanjing Han people (each plasma sample was 250 μL and each pool contained 12.5 mL). The TLDA was carried out on an ABI PRISM 7900HT Sequence Detection System (Applied Biosystems, Foster City, CA, USA) to analyse the miRNA profiles of 754 different human miRNAs. The concentrations of the miRNAs are presented as Cq values and were normalized to an internal reference gene recommended by the manufacturer. The relative miRNA concentrations were calculated by the comparative Cq method (2−ΔCq). For the qRT-PCR assay, the total RNA was extracted from 100 μL individual plasma samples using a 1-step phenol/chloroform purification protocol. A TaqMan probe-based qRT-PCR assay was conducted on a 7300 Sequence Detection System (Applied Biosystems). All reactions were run in triplicate and the average Cq values were calculated. To control for variability in the RNA extraction and purification procedures, an exogenous reference gene, plant miRNA MIR2911 (5′-GGCCGGGGGACGGGCUGGGA), was spiked into each sample at a final concentration of 106 fmol/L during RNA isolation. This gene served as a synthetic external reference for the normalization of plasma miRNAs as it has no mammalian homologue and the measurement of its expression has high repeatability and reproducibility (Supplementary Methods and Supplementary Fig. S1 and Fig. S2). The relative levels of miRNAs were normalized to MIR2911 and were calculated using the comparative Cq method (2−ΔCq). The total RNA of peripheral blood cells was extracted with TRIzol Reagent according to the manufacturer’s instructions. The concentration and quality of the extracted RNA was determined using a spectrophotometer (Eppendorf, Hamburg, Germany) at 260 nm and 280 nm. To calculate the relative expression levels of target miRNAs, U6 was used as an internal reference for the qRT-PCR analysis of miRNAs in peripheral blood cells.
Hematology and growth factor analysis
Hematological indices, including RBC, HGB, HCT, MCH and PLT levels, were measured using XE-2100 analysers (Sysmex, Kobe, Japan) with commercial reagents immediately after blood collection. Growth factors including EPO and VEGFA were measured using commercial ELISA kits (Cusabio, Wuhan, China).
Statistical analysis
All statistical analyses were performed using the Statistical Analysis System software SPSS 17.0. Data are presented as the mean ± SEM for miRNAs or the mean ± s.d. for other variables. The differences in variants among groups were analysed using a one-way ANOVA and the differences between groups were subsequently determined using a nonparametric Mann-Whitney U-test. A two-sided x2 test was used to compare sex distribution between two groups. A P-value < 0.05 was considered statistically significant. Correlations of miRNA profiles between two groups were calculated by using Pearson correlation analysis and the co-correlation of the miRNAs and their relationship with other parameters were calculated using Spearman rank correlation analysis.
Additional Information
How to cite this article: Yan, Y. et al. Influence of a high-altitude hypoxic environment on human plasma microRNA profiles. Sci. Rep. 5, 15156; doi: 10.1038/srep15156 (2015).
References
Calin, G. A. & Croce, C. M. MicroRNA signatures in human cancers. Nat Rev Cancer 6, 857–866 (2006).
Small, E. M. & Olson, E. N. Pervasive roles of microRNAs in cardiovascular biology. Nature 469, 336–342 (2011).
Chen, X. et al. Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res 18, 997–1006 (2008).
Mitchell, P. S. et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci USA 105, 10513–10518 (2008).
Zhang, C. et al. Expression profile of microRNAs in serum: a fingerprint for esophageal squamous cell carcinoma. Clin Chem 56, 1871–1879 (2010).
Luque, A. et al. Usefulness of circulating microRNAs for the prediction of early preeclampsia at first-trimester of pergnancy. Sci Rep 4, 4882 (2014).
Wang. C. et al. A panel of five serum miRNAs as a potential diagnostic tool for early-stage renal cell carcinoma. Sci Rep 5, 7610 (2015).
Meder, B. et al. Influence of the confounding factors age and sex on microRNA profiles from peripheral blood. Clin Chem 60, 1200–1208 (2014).
Simonson, T. S. et al. Genetic evidence for high-altitude adaptation in Tibet. Science 329, 72–75 (2010).
Huerta-Sànchez, E. et al. Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA. Nature 512, 194–197 (2014).
Nallamshetty, S., Chan, S. Y. & Loscalzo, J. Hypoxia: a master regulator of microRNA biogenesis and activity. Free Radic Biol Med 64, 20–30 (2013).
Yan, H. L. et al. Repression of the miR- 17-92 cluster by p53 has an important function in hypoxia-induced apoptosis. EMBO J 28, 2719–2732 (2009).
Rupaimoole, R. et al. Hypoxia-mediated downregulation of miRNA biogenesis promotes tumor progression. Nat Commun 5, 5202 (2014).
Luo, Y., Wang, Y., Lu, H. & Gao, Y. ‘Ome’ on the range: update on high-altitude acclimatization/adaptation and disease. Mol Biosyst 10, 2748–2755 (2014).
Sarkar, S., Banerjee, P. K. & Selvamurthy, W. High altitude hypoxia: an intricate interplay of oxygen responsive macroevents and micromolecules. Mol Cell Biochem 253, 287–305 (2003).
Biggar, K. K., Kornfeld, S. F., Maistrovski, Y. & Storey, K. B. MicroRNA regulation in extreme environments: differential expression of microRNAs in the intertidal snail Littorina littorea during extended periods of freezing and anoxia. Genomics Proteomics Bioinformatics 10, 302–309 (2012).
Kulshreshtha, R. et al. A microRNA signature of hypoxia. Mol Cell Biol 27, 1859–1867 (2007).
Camps, C. et al. Hsa-miR-210 is induced by hypoxia and is an independent prognostic factor in breast cancer. Clin Cancer Res 14, 1340–1348 (2008).
Saito, K., Kondo, E. & Matsushita, M. MicroRNA 130 family regulates the hypoxia response signal through the P-body protein DDX6. Nucleic Acids Res 39, 6086–6099 (2011).
Hebert, C., Norris, K., Scheper, M. A., Nikitakis, N. & Sauk, J. J. High mobility group A2 is a target for miRNA-98 in head and neck squamous cell carcinoma. Mol Cancer 6, 5 (2007).
Hua, Z. et al. MiRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS One 1, e116 (2006).
Zhang, P., Yao, Q., Lu, L., Li, Y., Chen, P. J. & Duan, C. Hypoxia-inducible factor 3 is an oxygen-dependent transcription activator and regulates a distinct transcriptional response to hypoxia. Cell Rep 6, 1110–1121 (2014).
Fong, G. H. & Takeda, K. Role and regulation of prolyl hydroxylase domain proteins. Cell Death Differ 15, 635–641 (2014).
Lofstedt, T. et al. Hypoxia inducible factor-2 alpha in cancer. Cell Cycle 6, 919–926 (2007).
Zundel, W. et al. Loss of PTEN facilitates HIF-1-mediated gene expression. Genes Devs 14, 391–396 (2000).
Arany, Z. et al. HIF-independent regulation of VEGF and angiogenesis by the transcriptional coactivator PGC-1alpha. Nature 451, 1008–1012 (2008).
Eric, D., Miao, Z., Bao-Han, V. & Shafiq, A. K. Vascular endothelial growth factor A, secreted in response to transforming growth factor-beta1 under hypoxic conditions, induces autocrine effects on migration of prostate cancer cells. Asian J Androl 14, 745–751 (2012).
Rasmussen, P., Siebenmann, C., Díaz, V. & Lundby, C. Red cell volume expansion at altitude: a meta-analysis and Monte Carlo simulation. Med Sci Sports Exerc 45, 1767–1772 (2013).
Wu, T. et al. Hemoglobin levels in Qinghai-Tibet: different effects of gender for Tibetans vs. Han. J Appl Physiol 98, 598–604 (2004).
Wu, T. Y., Liu, F. Y., Ouzhou-Loubu, Cui, C. Y., Qi, X. B. & Su, B. A genetic adaptive pattern-low hemoglobin concentration in the Himalayan highlanders. Zhongguo Ying Yong Sheng Li Xue Za Zhi 29, 481–493 (2013).
Fuchs, O. et al. Inhibition of smad5 in human hematopoietic progenitors blocks erythroid differentiation induced by BMP4. Blood Cell Mol Dis 28, 221–233 (2002).
Cantu, C. et al. Sox6 enhances erythroid differentiation in human erythroid progenitors. Blood 117, 3669–3679 (2011).
Garzon, R. et al. MicroRNA fingerprints during human megakaryocytopoiesis. Proc Natl Acad Sci USA 103, 5078–5083 (2006).
Hock, H. et al. Tel/Etv6 is an essential and selective regulator of adult hematopoietic stem cell survival. Genes Dev 18, 2336–2341(2004).
Xu, S. et al. Genomic dissection of population substructure of Han Chinese and its implication in association studies. Am J Hum Genet 6, 762–774 (2009).
Acknowledgements
C.Y.Z. is supported by the grants from national basic research program of China (no. 2014CB 542300) and the research special fund for public welfare industry of health of China (no. 201302018). C.Z. is sponsored by national natural science foundation of China (NSFC 81472021 and 81171661) and the medical scientific research foundation of Nanjing Military Command (no. 12Z28). C.W. is supported by national natural science foundation of China (NSFC 81401257), natural science foundation of Jiangsu province (BK20140730) and the fundamental research funds for the central universities (no. 021414330028). J.W. is supported by national natural science foundation of China (NSFC 81271904).
Author information
Authors and Affiliations
Contributions
C.Z. and C.Y.Z. conceived and designed the study. C.Z. and Y.Y. wrote the manuscript. Y.Y., Y.S. and C.W. performed the experiments. Y.Y. performed the statistical analysis. P.G. and J.W. contributed reagents, materials and analysis tools.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Yan, Y., Shi, Y., Wang, C. et al. Influence of a high-altitude hypoxic environment on human plasma microRNA profiles. Sci Rep 5, 15156 (2015). https://doi.org/10.1038/srep15156
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep15156
This article is cited by
-
Different expression pattern of human cytomegalovirus-encoded microRNAs in circulation from virus latency to reactivation
Journal of Translational Medicine (2020)
-
Role of miRNAs in hypoxia-related disorders
Journal of Biosciences (2018)
-
Genome-wide Long Non-coding RNA Analysis Identified Circulating LncRNAs as Novel Non-invasive Diagnostic Biomarkers for Gynecological Disease
Scientific Reports (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
