
Abstract
Mosquitoes are one of the deadliest and most hazardous animals on Earth, where they transmit several diseases that kill millions of people annually. There is an ongoing search almost everywhere in the world for more effective and contemporary ways to control mosquitoes other than pesticides. Phytochemicals are affordable, biodegradable biological agents that specialize in eliminating pests that represent a risk to public health. The effectiveness of Acacia nilotica methanol and aqueous leaf extracts against 4th instar larvae was evaluated. The results revealed that the methanol extract of A. nilotica had a noticeable influence on the mortality rate of mosquito larvae, especially at high concentrations. Not only did the mortality rate rise significantly, but the hatching of the mosquito eggs was potentially suppressed.Terpenes, fatty acids, esters, glycosides, pyrrolidine alkane, piperazine, and phenols were the most prevalent components in the methanol extract, while the aqueous extract of A. nilotica exclusively showed the presence of fatty acids. The insecticidal susceptibility tests of both aqueous and alcoholic extract of A. nilotica confirmed that the Acacia plant could serves as a secure and efficient substitute for chemical pesticides because of its promising effect on killing larvae and egg hatching delaying addition to their safety as one of the natural pesticides. Molecular docking study was performed using one of the crucial and life-controlling protein targets, fatty acid binding protein (FABP) and the most active ingredients as testing ligands to describe their binding ability. Most of the structurally related compounds to the co-crystallized ligand, OLA, like hexadecanoic acid furnished high binding affinity to the target protein with very strong and stable intermolecular hydrogen bonding and this is quite similar to OLA itself. Some other structural non-related compounds revealed extraordinarily strong binding abilities like Methoxy phenyl piperazine. Most of the binding reactivities of the majortested structures are due to high structure similarity between the positive control, OLA, and tested compounds. Such structure similarity reinforced with the binding abilities of some detected compounds in the A. nilotica extract could present a reasonable interpretation for its insecticidal activity via deactivating the FABP protein. The FABP4 enzyme inhibition activity was assessed for of both methanolic and aqueous of acacia plant extract and the inhibition results of methanol extract depicted noticeable potency if compared to orlistat, with half-maximal inhibitory concentration (IC50) of 0.681, and 0.535 µg/ml, respectively.
Similar content being viewed by others


Introduction
Mosquitoes are a crucial insect because of their significance as a vector in disease transmission1. Diseases including dengue, malaria, filariasis, west nile fever, lumpy skin disease, yellow fever, and Japanese encephalitis can be spread by them2,3,4,5,6. In recent years, dengue viruses, which are spread by infected female Culicidae mosquitoes, specifically Aedes aegypti and Aedes albopictus—have grown to be a serious global public health problem7,8,9. Gibbons identified Aedes aegypti as the primary vector of arboviral dengue virus infections in tropical and subtropical regions10,11. Worldwide, between 50 and 100 million people contract the disease each year, and around 2.5 percent of those cases result in death12.
The annual incidence of malaria is approximately 4–5 million in poor countries, such as India, where about 236 million people live in filariasis-endemic areas. Dengue fever still poses a threat to millions of people in different parts of the globe13. The most important vectors implicated in disease transmission to humans are the genera Culex, Aedes, and Anopheles14.
Several control approaches are routinely employed to control mosquito-borne risks. Insects are compelled to adapt to synthetic insecticides, which have been around for a while. Moreover, continuous use of pesticides poses concerns to the ecosystem since insects are becoming resistant to the chemicals15. Phytochemicals represent a significant substitute for synthetic insecticides. Mosquitoes are affected by phytochemicals in a number of ways, including as oviposition deterrent, developmental toxin, hatching blocker, adulticidal, ovicidal, and emergence blocker16.
The effectiveness of several medicinal plant preparations as larvicidal agents or as mosquito repellents has not yet been determined. Recently, plant-based insecticides and those that don’t hurt the environment have received increased attention while creating new pesticides17,18,19. Therefore, natural substances are the ideal alternative since they are eco-friendly, bio-sourced, and safe to use, causing less harm to the ecosystem and non-target species. Furthermore, a variety of extracts and compounds derived from diverse plant families have been investigated as possible new larvicides20. Studies have revealed that saponin21, steroids22, isoflavonoids23, essential oils24, alkaloids25, and tannins26 are all effective against mosquito larvae.
The plants are known to have an extensive number of bioactive chemicals that are both sustainable and regenerative. Royal Poinciana, Delonix regia is one of most interested plants which are widely distributed in temperate and tropical regions, and its leaves have long been used in folk medicine, notably as an antimalarial, particularly in East Asia. A wide spectrum of bioactivities was reported for the Delonix Regia extract. The plant has demonstrated positive antioxidant and antimicrobial properties27,28,29. Additionally, it has been reported that the flower's acetone and methanol extracts have effective larvicidal properties.
Plant products are one of the finest solutions for mosquito control, and numerous plant treatments were attempted before the discovery of chemical pesticides30,31,32,33. Moreover, the expanding trend and favorable community reaction to phytochemicals and their ecologically beneficial behavior offer an open field for plant-based pesticide research and innovation. Keeping in view the toxic activities of the family Fabaceae and many other plant families, which were demonstrated against other mosquito species34,35.
The Domain of therapeutical targets and drug design has been developed and expanded after the full accomplishment of human genome project, synchronizing with extensive and devolving advanced strategies throughputs related to x-rays diffraction (XRD), nuclear magnetic resonance (NMR) and Fourier transform infra-red (FTIR) and other structure- identifying tools permits to more success in knowing both protein–ligand and protein complex structure easily36. The rapid and effectual in structure identifications lead to a jump in computer-aided drug design and consequently the molecular docking, which presented a theoretical-based simulation between the drug and the host protein to a special part called pocket, such interactions are controlled by some sophisticated calculations. Nowadays, most of the research articles comprise one of the artificial intelligence applications especially that deals with ligand–protein37 and protein–protein interactions37. Synthetic and therapeutical chemistry are one of the most probable branches that heavily depends on the drug design and molecular docking38. The molecular simulation docking facilitates the selection of the most powerful candidate form huge databases and consequently facilitating the Structure Activity Relationship (Q-SAR) which connect between the structure and its biological impact. Such theoretical studies linked the drug candidate physical activity, e.g., biological activity, and their structure, making the biological activity easier to be predicted before the chemical synthesis39. Molecular docking used in various chemical design and synthesis of some EGFR analogues40, PI3k41, carbonic anhydrase42, acetylcholinesterase43, topoisomerase44 and m-TOR inhibitors45.
The effectiveness of plant extracts and its activated ingredients against mosquito larvae varies greatly depending on many factors related to the plant itself environment that the plant grew up, components of the plant used, its age (young, mature, or senescent), and the solvent used during the extraction. So, this study was designed to compare the lethal effects of some Fabaceae plants on filarial vectors Cx. pipiens and Rift Valley fever, Ae. aegypti. Moreover, both aqueous and methanolic acacia extract were prepared and the most activated ingredients were expected using GC/MS analysis and some of the detected compounds at high ratio were taken into consideration. All detected compounds were investigated to the Molecular docking simulation to assess their activity to interact and inhibit FABP protein, where 2flj, fatty acid binding protein, was used as the target protein and the most abundant active ingredients in the plant extract as tested ligands. The structure activity relationship between OLA and most of the tested compounds helps to give a convenient interpretation of the inhibition abilities to the FABP protein and may interpret the potential biochemical change due to the treatment using both types of acacia extracts.
Materials and methods
Ethical statement
All plant experiments in the study were conducted in accordance with relevant guidelines and legislation of ethical committee of Benha University.
Plant materials and analysis
Plant collection and extraction
Leaves of Acacia plants, Acacia nilotica (Fabaceae) were collected from different areas in Qalyubiya Governorate, Egypt, during August–September 2022. Plants were identified in the Flora and Phytotaxonomic Section of the Agricultural Research Centre in Giza, Egypt. The plant components were shade-air-dried at room temperature for seven days until the plant was thoroughly dried and the dry weight contracted. To keep the dried tissues free from moisture, they were pulverized in an electric mixer made of stainless steel and placed within airtight containers. To make the methanol extract, 20 g of dry plant materials were soaked in 100–150 ml of methanol in a conical flask (cap. 250 ml) at room temperature 27 ± 2 °C. Another 20 g of plant components were mixed with warm water (60 °C) at room temperature. After 48 h, the solutions were filtered via a Buchner funnel using Whatman No. 1 filter paper. A rotary evaporator was used to concentrate the extract for 3–5 h at room temperature (28 °C) and solid product then stored at 4 °C until used46.
Mosquito larvicidal assay
Culex pipiens rearing
Larvae of Cx. pipiens were raised in the insectary at the Department of Entomology, Faculty of Science, Benha University, Egypt, for 12 generations, where they were kept at 27 ± 2 °C and 75 ± 5% relative humidity during a light/dark photoperiod of 12:12 h. The larvae were fed a 3:1 ratio of ground bread to fish meal (Tetramin). After being transferred from the enamel pans to a cup of dechlorinated water, the pupae were placed in screened cages of 35 × 35x40 cm, from which the adults finally emerged. The adult mosquito colony received a 10% sugar solution, while the females were periodically fed blood by a hamster rat. Two developmental stages, larvae, and pupae were consistently accessible for the testing and maintained in the same lab.
Larvicidal activity
Under laboratory circumstances, the plant leaf methanol, and aqueous extracts of A. nilotica were tested against the 4th larval instar of Cx. pipiens (No. = 20 larvae). The fourth larval instar was exposed to doses of 62.5, 125, 250, 500, 1000, and 1500 ppm (1 g/1000 ml distilled water). Twenty larvae were introduced to a glass beaker holding 250 ml of distilled water for each concentration. Each concentration received three replicates. Temephos was employed at a rate of 0.25 ppm as a positive control according to WHO47 standard concentration within a similar volume of test beaker48. Mortality rates were obtained 24 h, 48 h, and 72 h after the first exposure and post-treatments (PT)12. No food was provided to the larvae during the experimental period.
Oviposition assay
To analyze the effects of Acacia extract concentration on oviposition, three concentrations of 250, 500, and 1000 ppm in transparent glass beakers (6.5 cm high and 4.5 cm diameter, volume 200 ml) were prepared and exactly, 75 Cx. pipiens females were tested. After taking the blood meal, the number of egg rafts in the three cups was monitored for four to five days. The experiment was repeated three times with untreated control groups (dechlorinated tap water only). The hatching rate of mosquito eggs was also determined according to Zheng et al.49, where all egg rafts were examined under binocular microscope. Hatch rate was calculated after 72 h by dividing the number of eggs observed and considered to be hatched by the total number of eggs present on the paper.
Efficacy of selected plant extracts against non-target predators
Acacia methanol and aqueous extracts were tested against Gambusia affinis and Sphaerodema urinator and, two of the among field predators that live in mosquito’s places. After catching, they were placed in plastic bags partially filled with water and transported to the lab. 24 h, predators were fasted prior to the bioassay. They were then treated with LC50 of Acacia extract, 3 predators per 1 l of dechlorinated water inside an enamel plate (35 × 25 × 15 cm) with 300 larvae. Three replicates were carried with controls.
GC/MS analysis
For the biochemical analyses, Thermo Scientific Trace GC Ultra/ISQ Single Quadrupole MS, TG-5MS fused silica capillary columns were utilized, with thicknesses of 0.1 mm, 0.251 mm, and 30 m. It was done with a 70 eV ionization energy electronic ionizer. Helium gas was employed as a carrier gas (flow rate: 1 ml/min). The MS transmission line and injector were both set to 280 °C. Preheating the oven to 35 °C, it was then raised to 150 °C at a rate of 7 °C per minute, 270 °C at a rate of 5 °C per minute (with a two-minute break in between), and finally 310 °C at a rate of 3.5 °C per minute (continual for 10 min). To investigate the quantification of every component found, a relative peak area was utilized. The substances were at least partially identified by matching their retention periods and mass spectra to NIST and Willy Library data from the GC/MS instrument. The aggregate spectrum of user-generated reference libraries was used for identification. Single-ion chromatographic reconstructions were carried out to assess peak uniformity. When possible, co-chromatographic analysis of reference chemicals was employed to validate GC retention durations50.
Fatty acid binding protein inhibition assay
Th inhibition assay of Fatty acid binding protein (FABP4) was determined according to Zimmerman and Veerkamp51. The solid aqueous and methanolic extracts were dissolved in DMSO to make the required serial dilution which has no detrimental effect on the fatty acid binding protein. FABP assay buffer was prepared via dilution of the (10 × buffer) by taking 2 ml and 18 ml HPLC-grade water. Followed by preparation of FABP assay detection reagent, FABP4 assay protein, arachidonic acid and inhibitor/ligand concentrations according to Cayman chemical kit recommendations. In 96 well plat, 40 µl of FABP assay buffer to each well followed by adding of 25 µl of the detection reagent and 10 µl of the inhibitor/ligand dilution. Then the plates were covered and incubated for 10 min and the fluorescence measured at 475 nm after excitation at 370 nm using a plate reader.
Molecular docking study
Source of the objective protein
The binding affinity of the recognized active ingredients from both aqueous and alcoholic acacia extract against lm-FABP binding site were examined using one of the theoretical approaches in molecular docking, MOE tool, to decide whatever the active ingredients could make successive binding with the target protein and accordingly suggesting the mode of action. There is not a three-dimensional structure of fatty acid binding protein owing to Cx. pipiens in the protein data bank, hence the well-known crystal three-dimensional structure of the Fatty acid binding protein from locust flight muscle in complex with oleate lm-FABP(PDB code:2FLJ)43 was utilized for this purpose and downloaded from protein data bank (https://www.rcsb.org/structure/2FLJ) as PDB format, water molecules were excluded in addition to any heteroatom subtracted and only chain a constrained.
Energy minimization
The most abundant and activated ingredients from both aqueous and alcoholic acacia extracts were automatically identified by the GC/MS. A database set of twenty candidates of the active ingredients were chosen to perform this study, were drawn using CAMBRIDGESOFT CHEMOFFICE 2015 Professional 15.0.0 software after recalling their smiles from PubChem database. The drawn structures were saved as Mol format after fulfillment energy minimization step using the defaults’ function Amber12: EHT forcefield till gradient convergence of 0.01 kcal/mol. Energy minimization step in addition to molecular simulation were done using the software Molecular Operating Environment MOE_2015.10, which installed by 64-bit operating system [Intel (R) Core (TM) i5-2400 CPU @ 2.40 GHz, 8 GB RAM] system.
Docking procedure
After uploading lm-FABP (PDB code: 2FLJ), both twenty candidate ligands and target protein were prepared, the reference drug (oleate) was assorted in green color to be distinguished easily. The binding site was recognized automatically from the surfaces and maps option and accordingly the co-crystallized ligand’s binding site, pocket site, was separated and saved as MDB file. After assorting the twenty candidates in energy-minimized in new database, the docking step was done from “compute” option using the defaults of “Rotate bonds” choice to permits flexible-ligand rigid receptor docking to be fulfilled. Additionally, the scoring energy function was changed to be London G with triangle matcher replacement were set. A “fifteen conformers” was re-adjusted instead of the “thirty” automatic option conformers of the best score energy done by the ligand. One of top five conformers of best energetically favored ligand–protein docking was chosen to represent the ligand docking and showed two-and three-dimensional receptor interactions52. Docking results of all the tested twenty candidates were assorted into one table, considering the preference of number of interactions, scoring energy (kcal/mol), RMSD (Å), and the bond length (Å). Three- and two-dimensional docking interaction were captured, and the green color was used to indicate the ligand under docking investigation to be easily distinguished, intermolecular hydrogen bonding and π-π staking (aromatic) were labelled in magenta and yellow color, respectively, meanwhile the loops, helical, etc. of protein structures were colored automatically by the software and some renderings set up are made for best manifestation show.
Fatty acid binding protein activity test (FABP4)
Fatty acid binding protein (FABPs) are group of intracellular liquid binding protein (iLBP) usually binds to intracellular hydrophobic ligands and shielding them throughout cellular compartments. The activity of FABP4 caused by both aqueous and methanolic acacia extracts treatment were observed to be diminished and the minimal inhibitory concentration caused by methanolic extract 0.681 µg/ml meanwhile aqueous extract 2.311 µg/ml against the positive control, orlistat, 0.535 µg/ml. Studies conducted in FABP4 levels in mice have proven that the down-fluctuations of such proteins are related to many metabolic diseases53,54. With crucial role future clinical treatments. Various effective FABP4 inhibitors were developed but unfortunately, none of them approved or even entered clinical trials55. BMS-309403, FABP4-ap2, is a very strong, selective adipocyte fatty acid binding protein inhibitor with minimal inhibitory concentration ki = 2 nM (0.9 µg/l), 250(118 µg/l), and 350(165 µg/l) nM for FABP4, FABP3, and FABP5, respectively56. Comparatively, Methanolic acacia extract exhibited good inhibition ability with ki = 0.68 mg/l and being from natural source that may encourage its safety attitude.
Statistical analysis
The data were analyzed using the software SPSS V23 (IBM, USA) to perform Probit analyses to compute lethal concentration (LC) values and the one-way analysis of variance (ANOVA) (Post Hoc/Turkey's HSD test). The significance threshold was established at P < 0.05.
Results
Mosquito larvicidal activity
This study assessed A. nilotica plant extracts on Cx. pipiens larvae in their fourth instar. In this study, every tested plant extract shown strong larvicidal efficacy against Cx. pipiens mosquito larvae. The findings of this study demonstrated that methanol extracts of plant materials had stronger harmful effects than aqueous extracts against Cx. pipiens mosquito larvae.
The mortality percent (MO%) of Cx. pipiens treated with 1500 ppm methanol extracts of A. nilotica at 24 h post-treatment (PT) was 100% (Table 1) and the corresponding LC50 (50%, median lethal concentration) = 200.50 ppm (Table 2), while the corresponding values for aqueous extracts were 98 (MO%) and the LC50 values = 256.40 ppm.
The larval mortality was maximum after 48 h of PT, while the mortality reaching 100% at 1500 ppm in methanol and aqueous extracts. In terms of fatal concentrations, A. nilotica methanol extracts revealed to be the most effective against Cx. pipiens larvae (LC50 = 140.13 ppm), followed by aqueous extracts (LC50 = 172.68 ppm). The larval mortality rate for the traditional positive control (temephos) was 73.3 and 100% at 24 and 48 h post treatment, respectively in aqueous and Methanol extracts.
Effect of Acacia extract concentration on eggs hatching rate
We tested the oviposition and hatchability rate of eggs in the A. nilotica solution. Exactly 25 Cx. pipiens females were used for the test and mixed with 30 of the mosquito males, and consequently, 122 egg rafts were counted, and the detailed number of laid eggs and hatching rate were summarized in Table 3 and Fig. 1. At a higher concentration of A. nilotica extract (1000 ppm), about 65 egg rafts were laid and reached a hatching rate of 24.4%. At the moderate concentrations (500 ppm), a moderate number of egg rafts were laid (37 eggs) with a 48.4% hatching rate compared to the low-concentrated A. nilotica extract (250 ppm), which produced around 15 egg rafts with a hatching rate of 76.5% of the total eggs laid. The t-test demonstrated that these results were statistically significant (P < 0.05).

Efficacy of A. nilotica plant extracts concentration on the oviposition and hatching rates of female Cx. pipiens.
The Efficacy of plant extracts against non-target predators
The effects of the Acacia plant extracts on non-target insects (G. affinis and S. urinator) were tested. They were treated with LC90 values of A. nilotica aqueous (847.23), and A. nilotica methanol (711.24 ppm), respectively, against these predators (Table 4). The mean predation for A. nilotica methanol and aqueous with chosen predators did not differ significantly (P > 0.05). According to the data, mosquito fish, G. affinis (78.67, 80.67%) and S. urinator 26, 25.67%) bug were not affected in both methanol and aqueous plant extracts (Table 4).
Phytochemical analysis
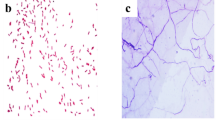
GC–MS analysis was used to perform a metabolic investigation of A. nilotica extracts and a comparison of aqueous and methanol extracts. Our study’s GC–MS analysis results allowed us to identify a variety of chemicals in the leaves of A. nilotica, including phenols, terpenes, esters, glycosides, fatty acids, piperazine, pyrrolidine and alkane (Tables 5 and 6). Methanolic leaf extract of A. nilotica showed the presence of 19 compounds (Table 5 and Fig. 2a), while aqueous leaf extract of A. nilotica showed 6 compounds (Table 6, Fig. 2b). Among them, 9,12,15-Octadecatrienoic acid (21.10%), squalene (18.25%), stigmasterol (14.25%), and 2,3-dimethoxy-anthracene-9,10-dione (5.65%) showed abundance in methanolic extract while, Cis-11-eicosenoic acid (68.10%), 11-Octadecenoic acid, methyl ester (20.14%), and 9,12-Octadecadienoic acid (z,z)-, methyl ester (8.80%) showed abundance in aqueous extract.

The TIC chromatograms of Acacia nilotica methanol (a) and aqueous extracts (b) chemical constituents detected by GC–MS.
FABP4 enzyme assay evaluation
The FABP4 enzyme inhibition assay for both methanolic and aqueous of acacia plant extract were estimated and orlistat was used as positive control and their half-maximal inhibitory concentration (IC50) 0.681, 2.311 and 0.535 µg/ml for the methanolic extract, aqueous extract and the lipase inhibitor, orlistat, respectively (Fig. 3).

FABP4 enzyme assay inhibition of the tested acacia extracts.
Docking study
The target protein Lm-FABP, PDB:2FLJ as fatty acid binding protein in Fig. 4 was chosen to perform docking study of twenty candidates of the most abundant active ingredient in aqueous and methanolic acacia extracts included: of Methoxyphenyl piperazine [1], 12-tridecynoic acid, methyl ester [2], pentadecanoic acid, 14-methyl-, methyl ester [3], hexadecanoic acid [4], methyl z-11-tetradecenoate [5], (z,z,z)-9,12,15-octadecatrienoic acid [6], stigmasterol [7], 7,10,13-eicosatrienoic acid, methyl ester [8], squalene [9], 1,8-diethyl-3,6-diazahomoadamantan-9-ol [10] Brenzkatechin [11], 2,3-dimethoxyanthracene-9,10-dione [12], (3á,5z,7e) 9,10-secocholesta-5,7,10(19)-triene-3,24,25-triol [13], Hexadecanoic acid, methyl ester [14], 11-Octadecenoic acid, methyl ester [15], 9,12-Octadecadienoic acid (z,z)-, methyl ester [16], Stearic acid methyl ester [17], Cis-11-eicosenoic acid [18], Ethyl iso-allocholate [19], 1,2,3-benzenetriol [20] were docked to the active center in the target protein presented in Fig. 4. The final simulation outcomes were listed in Table 7. The Candidate Methoxy phenyl piperazine [1] reacted similarly like the co-crystallized ligand, OLA presented in (Fig. 5, and revealed three different dipole–dipole interactions with the residues arginine and tyrosine with bond lengths between 2.15 and 2.48 Å as shown in Fig. 6. Compound [4], hexadecanoic acid, furnished three hydrogen bonds Fig. 7 with Gln98, thr76 and Asp75, at bond length between 2.01 to 3.41 Å. All Compounds (z,z,z)-9,12,15-octadecatrienoic acid [6], 10,13-eicosatrienoic acid, methyl ester [8], 9,12-Octadecadienoic acid (z,z)-, methyl ester [16], Stearic acid methyl ester [17], Cis-11-eicosenoic acid [18] and Ethyl iso-allocholate [19] shared two intermolecular hydrogen interactions with different residues including arginine and tyrosine as showen in Figs. S1 and S2. Some compounds like 12-tridecynoic acid, methyl ester [2], 1,8-diethyl-3,6-diazahomoadamantan-9-ol [10], Brenzkatechin [11], of 2,3-dimethoxyanthracene-9,10-dione [12], (3á,5z,7e)9,10-secocholesta-5,7,10(19)-triene-3,24,25-triol [13] 11-Octadecenoic acid, methyl ester [15] and 1,2,3-benzenetriol [20] shared less number of hydrogen bonds, only one electrostatic force, as shown in Figs. S3 and S4. Some other compounds have no compatibility to bind to this pocket like pentadecanoic acid, 14-methyl-, methyl ester [3], stigmasterol [7] squalene, and Hexadecanoic acid, methyl ester [14] as showen in Fig. S5 (Fig. 7).

Three-dimensional structure of 2flj and its active pocket.

Self-Docking of the co-crystallized ligand (Oleic acid or OLA) interior 2FLJ pocket: (a) two-dimensional receptor interactions; (b) three-dimensional receptor interactions; (c) three-dimensional positioning in the receptor pocket.

Ligand-Docking of compound 1 (Methoxy phenylpiperazine) interior 2FLJ pocket: (a) two-dimensional receptor interactions; (b) three-dimensional receptor interactions; (c) three-dimensional positioning in the receptor pocket.

Ligand-Docking of compound 4 (hexadecanoic acid) interior 2FLJ pocket: (a) two-dimensional receptor interactions; (b) three-dimensional receptor interactions; c) three-dimensional positioning in the receptor pocket.
Discussion
Plant extracts usually combines of various safe, and biodegradable essential oils (EOs) active ingredients may use as pests and disease control that killing harmful pests effectively. Merely 5% of pesticides worldwide are biopesticides, despite their advantageous effects as insecticides57. However, due to their special qualities that promote their application, such as their non-toxicity to the environment, biopesticides are growing quickly and are predicted to surpass chemical pesticides in the near future at an average annual growth rate of 9–20%58,59,60.
All the Acacia plant extracts that were evaluated in this study demonstrated a high level of insecticidal efficacy against mosquito larvae. Acacia methanol extracts killed mosquito larvae at 100%, 24 and 48 h post-treatment, and they were more effective than Aqueous extract. Our findings corroborated those of Baeshen and Baz35 research on the effects of extracts from Salix safsafs, Eucalyptus camaldulensis, and A. nilotica on Cx. pipiens larvae. They discovered that the extract from A. nilotica reduced the number of female eggs and killed mosquito larvae (100% mortality). Similarly, A. nilotica acetone leaf extracts were found to be acutely toxic at 212.1 mg/l and chronically toxic at 144.2 mg/l by Zaitoun et al.61, also who referred to the ability of A. nilotica extracts on inhibit adult emergence and egg hatchability.
Many multifunctional active ingredients have apparently been discovered in the seeds and leaves of A. nilotica62,63. Consequently, extracts from A. nilotica's fruits and leaves have demonstrated potential as fungicides, bactericides, insecticides, and molluscicides62,64. Vivekanandhan et al.65 assessed the bio efficacy of A. nilotica seed essential oil and seed pod solvent extracts against three significant mosquito species 24 h after treatment. Strong larvicidal efficacy against Anopheles stephensi (LC50 = 5.239 mg/L), Aedes aegypti (LC50 = 3.174 mg/L), and Cx. quinquefasciatus (LC50 = 4.112 mg/l) was demonstrated by the hydrodistilled seed oil. In contrast, the smoke toxicities for Cx. quinquefasciatus, Ae. aegypti, and An. stephensi adults were 82%, 90%, and 80%, respectively65.
In addition to their effectiveness on mosquito larvae, A. nilotica's methanol and acetone bark extracts were examined using an artificial diet bioassay for their potential to inhibit the growth of melon fly (Bactrocera cucurbitae) larvae in their first, second, and third instars. Also, pupation and emergence percentages were reported to be markedly decreased66. Studies have shown that extracts from A. nilotica can be effective in killing mosquito larvae. These extracts can disrupt the larvae's life cycle and prevent them from developing into adult mosquitoes14,67,68. In addition to being highly effective against other insects65,69 and microbes63,70.
A. nilotica extracts have been shown to be safe in research studies, meaning they can be used in aquatic environments where mosquitoes live. To ensure they are safe for existing organisms and to see what would happen if these extracts were used extensively, we evaluated these extracts on some non-target aquatic organisms common in mosquito larval habitats. It was found that there were no significant differences (P > 0.05) when applying aqueous extracts to predators, whether with aqueous or ethanol extracts.
Three of the non-target aquatic biocontrol agents were less hurt after being treated with A. caesia extracts and Ag NPs than mosquito larvae. The A. caesia LC50 ranged from 684 to 2245 μg/ml. Overall, our study shows that A. caesia has only a moderate effect on non-target aquatic biocontrol agents like A. bouvieri, D. indicus, and G. affinis fishes71. Specifically, the literature articles discussing the effects of the extract from the Acacia plant on fish or non-target organisms are rarely. However, generally speaking, a large number of literary articles support the idea that medicinal plants are not toxic to fish or non-target organisms. Promsiri et al.72 found that the guppy fish (Poecilia reticulata) was a common non-target organism found in Ae. aegypti habitats at levels that were effective at killing mosquito larvae. This is an example of this. They discovered that extracts from 112 different medicinal plant species showed no or very low toxicity to guppy fish. In another study that conducted the toxicological effect of Plumbago zeylanica and Cestrum nocturnum plant extracts against the fish species P. reticulata, which are the most widespread non-target organisms in A. aegypti environments, almost no significant harm was reported at doses of LC50 and LC9073.
As an attractive point, the descending concentration gradient leads to a descending rate of oviposition and hatching rates. At high concentrations of A. nilotica extract, the oviposition rate recorded its maximum value, and more than 50% of the total eggs were laid. Similarly, oviposition attitude was observed at moderate concentration, and less than 31% of the total eggs were counted. In agreement, the use of a very low concentration of A. nilotica showed a noticeable downfall of the number of eggs laid to less than 13% and 5% of the total eggs laid for the concentrations 250 ppm and zero (control without extract), respectively, confirming the direct relationship and oviposition rate. Moreover, the hatching rate is inconsistent with the oviposition rate. The hatchability rates were affected by the concentration gradient, and at zero concentration, the hatching ability was raised to 95% of the eggs laid at this concentration, whereas the higher concentration revealed a lower hatchability rate, confirming the inverse relationship between the concentration and the hatching rate. Based on the experimental data, the concentration of A. nilotica extract is directly proportional to oviposition rate and inversely proportional to hatching ability, which may introduce a reasonable interpretation for the dual benefit of acacia. Through what some active ingredients do to attract insects74,75, perhaps through extracting odorant chemical code, at the same time, the rest of the active ingredients help kill the larvae and reduce the chances of eggs hatching76.
Bioactive plant chemicals, bacteria, fungus, viruses, and protozoa are natural sources of bioinsecticides, as are pheromones and microorganisms. According to their source, the four main classes of bioinsecticides are phytochemicals, plant-incorporated protectants, microbiological pesticides, and pheromones77. They have been successfully used to manage and control pests78. Because they are more potent at very low doses, being biodegradable, less toxic, and target-specific than synthetic compounds, they are superior to them.
Plants are biological factories that generate a wide range of compounds referred as secondary metabolites. The main pharmacological effects of all medicinal plants are attributable to those secondary metabolites, which include alkaloids, carbohydrates, flavonoids, saponins, tannins, and terpenoids. Several different types of bioactive compounds that are isolated from plants or added to botanical pesticides may have negative effects on pests or other species that eat them or come into contact with them79.
The study examined the organic compounds present in the methanol and aqueous extracts of A. nilotica. The results indicated that the methanol extract had a greater variety of organic compounds, including terpenes, steroids, polyethylene glycol, and fatty acid compounds, whereas the aqueous extract had a higher concentration of fatty acid contaminants. The extracting solvents’ polarity had an impact on the antioxidant qualities, phytochemical content, and extraction yield80. Different extraction yields were obtained with different solvents. This is because the plant materials include high quantities of polar chemicals that are soluble in high polarity solvents, which might lead to a considerable fluctuation in the quantity of bioactive compounds in the extract due to changes in the polarity of the solvents. The findings showed that acetone contained more phytochemicals than aqueous extract, including phenols, terpenes, flavonoids, alkanes, and ketone81.
We find that depending on the type of solvent used in extraction and the presence of the Acacia plant in a specific area with specific climate, a clear variation appears in the secondary compounds. According to Edriss et al.68, the A. nilotica water and ethanol extracts revealed a broad spectrum of secondary metabolites, primarily alkaloids, saponins, flavones, and tannins, but the petroleum ether extracts were primarily composed of triterpenes and sterols. Aqueous and alcohol extracts of the plant A. nilotica were subjected to phytochemical screening. Acacia nilotica fruit showed the existence of phenol, terpenoid, flavonoid, reducing sugar, steroid, and flavonoids82.
A. nilotica contains various bioactive compounds, including tannins, alkaloids, and flavonoids. These compounds are believed to be responsible for its larvicidal activity, where in the Table 4 there are some of the active ingredients that are used in medical treatments, such as Methoxyphenyl piperazine (Piperazine).
Piperazine and its derivatives have attained a unique position as imperative pharmacophores in several therapeutic areas. Researchers have always been interested in using piperazine molecules as a building block to make new drug molecules that have a lot of different biological activities83 because of their flexible structure. Neophytadiene is a good analgesic, antipyretic, anti-inflammatory, antimicrobial, and antioxidant compound84.
The mission of carrying lipids and poorly soluble molecules such as retinoids, fatty acids and bile acids in addition to all hydrophobic constituents transported by the cytosol for variant purposes of storage or even utilization, is one of the intracellular binding proteins’s roles in the living biology. Such proteins are a family of low-molecular weight and single chain polypeptides binds to the hydrophobic components, from different origins, forming a matrix of complex capable of carrying poorly soluble compartments into cytosol or any organelles in the cell, this complex keep the sensitive poorly soluble components shielded from decomposition. Fatty acid binding proteins, FABPs, aids to synthesize phospholipids, lipid metabolism and mitochondrial beta oxidation furthermore a large number of functional roles of FABPs have been proposed based on physiological disorder in both animal and insect kingdoms85. FABPs usually expressed largely in both vertebrates and invertebrate86. FABPs classified into main categories in mammels constrained epidermal (E-FABP), intestinal (I-FABP), brain (B-FABP), haert (H-FABP), myelin (m-FABP), testis (T-FABP), retina (FABP12) and liver FABPs87. Majority of FABP proteins share high amino acid sequence similarities more than 70% however, their three-dimensional structure was highly constrained β-barrel structure with ligand binding cavity88,89,90.
The first FABP discovered was desert flight muscle locust of Schistocerca gregaria and Haunerland et al.91 followed by two abundant MFB1 and MFB2 were isolated from the midgut of Manduca sext larvae92. FABPs from different and variant insects heavily affects insect physiological metabolism by regulating intracellular fatty acids, it regulates sleep and long term memory consolidation, affects lipid accumulation in addition to their role in nutritional exploitation and caste division of many insects93. The change in FABPs chemistry will promote extensive physiological and behavioral attitude of insects. Hereon we tried to introduce a rationalized mode of action describing the activity of acacia extract, and how it affects Cx. Pipiens, taking the advantages of the great structure similarity of their fatty acid binding proteins (FABPs) for most insects. We used lm-FABP (PDB code:2FLJ) as model to evaluate each single component of the activated ingredients in acacia extract and how it binds to the target FABP.
Qualitatively, the docking results of most of the activate ingredients detected by the GC/MS were found to be close to the positive control, OLA, Due to the structure similarity between most of detected compounds and OLA. The results divided into four main categories the first one is some of tested compound revealed the same number of bindings with great similarity of amino acids residues like compound 1, methoxyphenyl piperazine, and compound 4, hexadecenoic acid, each one on them revealed three dipole–dipole interactions. The second category was recognized with candidates bonded with two hydrogen bonds like compounds (z,z,z)-9,12,15-octadecatrienoic acid [6]; 7,10,13-eicosatrienoic acid, methyl ester [8]; 9,12-Octadecadienoic acid (z,z)-, methyl ester [16]; Stearic acid methyl ester [17]; Cis-11-eicosenoic acid [18] and Ethyl iso-allocholate [19]. The nature of hydrogen bond shares the same properties of length, they have comparable length, and with the same amino acids’ residues e.g., the ester group in compound 8 interacted with Try130 and Arg128 amino acids, this is quite similar to the reference drug OLA.
Similarly, compound 18 and 19 revealed the same number of interactions. As shown in Fig. 8 (supplementary file) compound 18 has relatively high and not preferable RMSD. Such value determines whatever the tested ligand could simulates the co-crystallized ligand or not. Energetically the accepted value of RMSD should not exceed 1.7 and higher values are energetically not favored41,42. On the other side, a group of six candidates of 10, 11, 12, 13, 15 and 20 showed a smaller number of bonds however some of these compounds interacted with the same residual amino acid as the reference drug e.g., compound 10. In addition to RMSD values, the scoring energy is a very beneficial physical property and all the selected poses revealed negatively charged scoring energy. The larger the negative value of the scoring energy becomes, the more stability it indicates, and for this compounds 8 and 17 have the lowest scoring energy of -7.4504 and -7.3073 kcal/mol, respectively.

Ligand-Docking and two-dimensional receptor interactions of compound 15 (9,12-Octadecadienoic acid (z,z)-, methyl ester) interior 2FLJ pocket.
From the docking results, most of the tested structures such as 2, 4, 5, 6, 8, 15, 16 17 and 18, showed at least one interaction resemble to the OLA. The most effective factor is the structure of that compounds and their structure-resemblance with oleate (OLA) co-crystallized ligand. Such compounds have long hydrocarbon chain prolonged from 14 to 20 carbon atoms (Fig. 9) and the reference drug consist of exactly 14 carbon backbone. Such resemblance makes the size of tested ligand fitting to the protein receptor pocket and that interprets the low RMSDs which strengthen the idea of compatibility. Additionally, the carboxylic (COOH) group and /or ester group (COOMe) presented in the tested compounds structurally agreed with the carboxylate (COO–) terminated the OLA.

Chemical structure of the most active and abundant ingredients of open chain acid and esters of acacia extract compartments and the structure similarity relationship compared to the co-crystallized ligand, OLA.
Extending carbon backbone more over e.g., squalene, even if it contains active part like multiple double bonds, but the size of the structure becomes non-compatible to the receptor pocket. Replacing the long chain hydrocarbon with polycyclic moieties as showed in Fig. 10 such as compounds 7 and 13 did not achieve the same purpose of binding ability.

chemical structure of the most active and abundant activate ingredients of polycyclic components of acacia extract.
Conclusion
Molecular hybridization and docking simulation were used to introduce a convenient mode of action and to suggest how is acacia extracts affects Cx. pipiens via studying the binding compatibility of each single component detected by the GC/MS to the FABP, as one the effective proteins the invasively affect not only Culex but all vertebrate and invertebrates. Three components manifested unique binding ability like OLA co-crystallized ligand. Some other candidates presented exceptionally good binding abilities with higher chances of replacing OLA. The activity prediction by simulation keeps the ways smoothed for the researchers to continue exploring the potential of A. nilotica and its extracts as a natural and sustainable means of controlling mosquito populations and reducing the spread of mosquito-borne diseases. However, the effectiveness of these extracts may vary depending on factors such as concentration, application method, and local environmental conditions, so further research and testing are necessary to refine and optimize their use.
Data availability
This article includes all data produced or analysed during this investigation.
References
Coetzee, M. How important are Dipteran vectors of disease in Africa?. Trans. R. Soc. Trop. Med. Hyg. 108(4), 179–180 (2014).
Selim, A., Megahed, A., Kandeel, S., Alouffi, A. & Almutairi, M. M. West Nile virus seroprevalence and associated risk factors among horses in Egypt. Sci. Rep. 11, 20932. https://doi.org/10.1038/s41598-021-00449-6 (2021).
Selim, A. & Abdelhady, A. The first detection of anti-West Nile virus antibody in domestic ruminants in Egypt. Trop. Anim. Health Prod. 52, 3147–3151. https://doi.org/10.1007/s11250-020-02339-x (2020).
Selim, A., Manaa, E. & Khater, H. Molecular characterization and phylogenetic analysis of lumpy skin disease in Egypt. Comp. Immunol. Microbiol. Infect. Dis. 79, 101699. https://doi.org/10.1016/j.cimid.2021.101699 (2021).
Selim, A., Manaa, E. & Khater, H. Seroprevalence and risk factors for lumpy skin disease in cattle in Northern Egypt. Trop. Anim. Health Prod. 53, 350. https://doi.org/10.1007/s11250-021-02786-0 (2021).
Selim, A. et al. Prevalence and animal level risk factors associated with Trypanosoma evansi infection in dromedary camels. Sci. Rep. 12, 8933. https://doi.org/10.1038/s41598-022-12817-x (2022).
Hamady, D. et al. Colonized Aedes albopictus and its sexual performance in the wild: Implications for SIT technology and containment. Parasit. Vect. 6, 1–9 (2013).
Conway, M. J., Colpitts, T. M. & Fikrig, E. Role of the vector in arbovirus transmission. Ann. Rev. Virol. 1, 71–88 (2014).
Baz, M. M., Selim, A., Radwan, I. T., Alkhaibari, A. M. & Khater, H. F. Larvicidal and adulticidal effects of some Egyptian oils against Culex pipiens. Sci. Rep. 12, 4406. https://doi.org/10.1038/s41598-022-08223-y (2022).
Gibbons, R. V. Dengue conundrums. Int. J. Antimicrob. Agents 36, S36–S39 (2010).
Selim, A., Almohammed, H., Abdelhady, A., Alouffi, A. & Alshammari, F. A. Molecular detection and risk factors for Anaplasma platys infection in dogs from Egypt. Parasit. Vect. 14, 429. https://doi.org/10.1186/s13071-021-04943-8 (2021).
WHO. Dengue and severe dengue. Geneva: World Health Organisation (2014). http://www.who.int/mediacentre/factsheets/fs117/en/. Accessed 6th June 2014.
Gingrich, J. B., Harlan, H. J., Perkins, P. V., Trosper, J. H. & DC, A. F. P. M. B. W. Regional Disease Vector Ecology Profile East Asia. Armed Forces Pest Management Board Washington DC (2002).
Elkhidr, M. E. et al. Toxicity of water extract of Acacia nilotica fruits against mosquito larvae: An experimental study. Entomol. Appl. Sci. Lett. 7, 84–90 (2020).
Sudo, M., Takahashi, D., Andow, D. A., Suzuki, Y. & Yamanaka, T. Optimal management strategy of insecticide resistance under various insect life histories: Heterogeneous timing of selection and interpatch dispersal. Evolut. Appl. 11, 271–283 (2018).
Bakkali, F., Averbeck, S., Averbeck, D. & Idaomar, M. Biological effects of essential oils: A review. Food Chem. Toxicol. 46, 446–475 (2008).
Pavela, R. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crops Prod. 76, 174–187 (2015).
Suman, T., Elumalai, D., Vignesh, A., Kaleena, P. & Murugesan, K. Evaluation of larvicidal activity of the aerial extracts of a medicinal plant, Ammannia baccifera (Linn) against two important species of mosquitoes, Aedes aegypti and Culex quinquefasciatus. Asian Pac. J. Trop. Dis. 2, S352–S355 (2012).
Radwan, I. T., Baz, M. M., Khater, H. & Selim, A. M. Nanostructured Lipid Carriers (NLC) for Biologically active green tea and fennel natural oils delivery: Larvicidal and adulticidal activities against Culex pipiens. Molecules 27, 1939 (2022).
Baz, M. M., Hegazy, M. M., Khater, H. F. & El-Sayed, Y. A. Comparative evaluation of five oil-resin plant extracts against the mosquito larvae, Culex pipiens Say (Diptera: Culicidae). Pak. Vet. J. https://doi.org/10.5555/20210221793 (2021).
Wiesman, Z. & Chapagain, B. P. Larvicidal activity of saponin containing extracts and fractions of fruit mesocarp of Balanites aegyptiaca. Fitoterapia 77, 420–424 (2006).
Chaudhuri, A. et al. EFNS guideline on the management of community-acquired bacterial meningitis: Report of an EFNS task force on acute bacterial meningitis in older children and adults. Eur. J. Neurol. 15, 649–659 (2008).
Joseph, C., Ndoile, M., Malima, R. & Nkunya, M. Larvicidal and mosquitocidal extracts, a coumarin, isoflavonoids and pterocarpans from Neorautanenia mitis. Trans. R. Soc. Trop. Med. Hyg. 98, 451–455 (2004).
Cavalcanti, E. S. B., Morais, S. M. D., Lima, M. A. A. & Santana, E. W. P. Larvicidal activity of essential oils from Brazilian plants against Aedes aegypti L. Memórias do Instituto Oswaldo Cruz 99, 541–544 (2004).
Martha Perez Gutierrez, R., Maria Neira Gonzalez, A. & Hoyo-Vadillo, C. Alkaloids from piper: A review of its phytochemistry and pharmacology. Mini Rev. Med. Chem. 13, 163–193 (2013).
Xing, F., Cheng, G., Yang, B. & Ma, L. Microencapsulation of capsaicin by the complex coacervation of gelatin, acacia and tannins. J. Appl. Polym. Sci. 91, 2669–2675 (2004).
Shabir, G. et al. Antioxidant and antimicrobial attributes and phenolics of different solvent extracts from leaves, flowers and bark of Gold Mohar [Delonix regia (Bojer ex Hook.) Raf.]. Molecules 16, 7302–7319 (2011).
Modi, A. et al. Delonix regia: Historic perspectives and modern phytochemical and pharmacological researches. Chin. J. Nat. Med. 14, 31–39 (2016).
Bala, A. & Rani, G. Green synthesis of AgNPs using Delonix regia bark for potential catalytic and antioxidant applications. Microsc. Res. Tech. 86(8), 911–922 (2023).
Mittal, P. Biolarvicides in vector control: challenges and prospects. J. Vect. Borne Dis. 40, 20 (2003).
Samada, L. H. & Tambunan, U. S. F. Biopesticides as promising alternatives to chemical pesticides: A review of their current and future status. Online J. Biol. Sci 20, 66–76 (2020).
Baz, M. M. et al. Novel pesticidal efficacy of Araucaria heterophylla and Commiphora molmol extracts against camel and cattle blood-sucking ectoparasites. Plants 11, 1682 (2022).
Radwan, I. T., Baz, M. M., Khater, H., Alkhaibari, A. M. & Selim, A. M. Mg-LDH nanoclays intercalated fennel and green tea active ingredient: Field and laboratory evaluation of insecticidal activities against Culex pipiens and their non-target organisms. Molecules 27, 2424 (2022).
Venkadachalam, R. et al. Mosquito larvicidal and pupicidal activity of Tephrosia purpurea Linn. (Family: Fabaceae) and Bacillus sphaericus against, dengue vector, Aedes aegypti. Pharmacogn. J. https://doi.org/10.5530/pj.2017.6.116 (2017).
Baeshen, R. S. & Baz, M. M. Efficacy of Acacia nilotica, Eucalyptus camaldulensis, and Salix safsafs on the mortality and development of two vector-borne mosquito species, Culex pipiens and Aedes aegypti, in the laboratory and field. Heliyon https://doi.org/10.1016/j.heliyon.2023.e16378 (2023).
Bolje, A. & Gobec, S. Analytical techniques for structural characterization of proteins in solid pharmaceutical forms: An overview. Pharmaceutics https://doi.org/10.3390/pharmaceutics13040534 (2021).
Dhakal, A., McKay, C., Tanner, J. J. & Cheng, J. Artificial intelligence in the prediction of protein-ligand interactions: Recent advances and future directions. Brief. Bioinform. https://doi.org/10.1093/bib/bbab476 (2022).
Pinzi, L. & Rastelli, G. Molecular docking: Shifting paradigms in drug discovery. Int. J. Mol. Sci. https://doi.org/10.3390/ijms20184331 (2019).
Batool, M., Ahmad, B. & Choi, S. A structure-based drug discovery paradigm. Int. J. Mol. Sci. https://doi.org/10.3390/ijms20112783 (2019).
Mohareb, R. M., Bagato, N. M. A. & Radwan, I. T. Design, synthesis, molecular docking, and biological studies of new heterocyclic compounds derived from β-diketones as novel EGFR and pim-1 inhibitors endowed with antitumor activity. Anti-Cancer Agents Med. Chem. Chem. (Form. Curr. Med. Chem.-Anti-Cancer Agents) 22, 2558–2576 (2022).
Radwan, I. T. et al. Design, synthesis, docking study, and anticancer evaluation of novel bis-thiazole derivatives linked to benzofuran or benzothiazole moieties as PI3k inhibitors and apoptosis inducers. J. Mol. Struct. 1265, 133454 (2022).
Zain-Alabdeen, A. I. et al. Synthesis and anticancer activity of new benzensulfonamides incorporating s-triazines as cyclic linkers for inhibition of carbonic anhydrase IX. Sci. Rep. 12, 16756 (2022).
Radwan, I. T. et al. Effect of nanostructure lipid carrier of methylene blue and monoterpenes as enzymes inhibitor for Culex pipiens. Sci. Rep. 13, 12522 (2023).
Elanany, M. A., Osman, E. E. A., Gedawy, E. M. & Abou-Seri, S. M. Design and synthesis of novel cytotoxic fluoroquinolone analogs through topoisomerase inhibition, cell cycle arrest, and apoptosis. Sci. Rep. 13, 4144 (2023).
Zhang, B. et al. Design, synthesis and biological evaluation of substituted 2-(thiophen-2-yl)-1, 3, 5-triazine derivatives as potential dual PI3Kα/mTOR inhibitors. Bioorgan. Chem. 95, 103525 (2020).
Baz, M. M., Selim, A. M., Radwan, I. T. & Khater, H. F. Plant oils in the fight against the West Nile Vector, Culex pipiens. Int. J. Trop. Insect Sci. 42, 2373–2380 (2022).
WHO. Instruction fordetermining the susceptibilityor resistance of mosquito larvaetoinsecticides. WHO/VBC/ 81.807 (1981).
Abai, M. R., Hanafi-Bojd, A. A. & Vatandoost, H. Laboratory evaluation of Temephos against anopheles Stephensi and Culex Pipiens Larvae in Iran. J. Arthropod-Borne Dis. 10, 510–518 (2016).
Zheng, M.-L., Zhang, D.-J., Damiens, D. D., Lees, R. S. & Gilles, J. R. Standard operating procedures for standardized mass rearing of the dengue and chikungunya vectors Aedes aegypti and Aedes albopictus (Diptera: Culicidae)-II-Egg storage and hatching. Parasit. Vect. 8, 1–7 (2015).
Salem, M. et al. Antibacterial activity of extracted bioactive molecules of Schinus terebinthifolius ripened fruits against some pathogenic bacteria. Microb. Pathog. 120, 119–127 (2018).
Zimmerman, A. W. & Veerkamp, J. H. New insights into the structure and function of fatty acid-binding proteins. Cell. Mol. Life Sci.: CMLS 59, 1096–1116. https://doi.org/10.1007/s00018-002-8490-y (2002).
Metwally, N. H., Radwan, I. T., El-Serwy, W. S. & Mohamed, M. A. Design, synthesis, DNA assessment and molecular docking study of novel 2-(pyridin-2-ylimino) thiazolidin-4-one derivatives as potent antifungal agents. Bioorgan. Chem. 84, 456–467 (2019).
Thompson, B. R., Mazurkiewicz-Muñoz, A. M., Suttles, J., Carter-Su, C. & Bernlohr, D. A. Interaction of adipocyte fatty acid-binding protein (AFABP) and JAK2: AFABP/aP2 as a regulator of JAK2 signaling. J. Biol. Chem. 284, 13473–13480. https://doi.org/10.1074/jbc.M900075200 (2009).
Adida, A. & Spener, F. Adipocyte-type fatty acid-binding protein as inter-compartmental shuttle for peroxisome proliferator activated receptor gamma agonists in cultured cell. Biochim. Biophys. acta 1761, 172–181. https://doi.org/10.1016/j.bbalip.2006.02.006 (2006).
Floresta, G. et al. Adipocyte fatty acid binding protein 4 (FABP4) inhibitors. A comprehensive systematic review. Eur. J. Med. Chem. 138, 854–873. https://doi.org/10.1016/j.ejmech.2017.07.022 (2017).
He, Y. et al. Optimization of potent, selective and orally bioavailable biphenyl scaffold as FABP4 inhibitors for anti-inflammation. Eur. J. Med. Chem. 253, 115319. https://doi.org/10.1016/j.ejmech.2023.115319 (2023).
Rakshit, A. et al. Biopesticide: Volume 2: Advances in Bio-Inoculants (Woodhead Publishing, 2021).
Kumar, J., Ramlal, A., Mallick, D. & Mishra, V. An overview of some biopesticides and their importance in plant protection for commercial acceptance. Plants 10, 1185 (2021).
Lahlali, R., El Hamss, H., Mediouni-Ben Jemâa, J. & Barka, E. A. The use of plant extracts and essential oils as biopesticides. Front. Agron. 4, 921965 (2022).
Selim, A., Abdelrahman, A., Thiéry, R. & Sidi-Boumedine, K. Molecular typing of Coxiella burnetii from sheep in Egypt. Comp. Immunol. Microb. Infect. Dis. 67, 101353. https://doi.org/10.1016/j.cimid.2019.101353 (2019).
Zaitoun, A., Madkour, M. & Shamy, M. Effect of three plants extracts on some bacterial strains and Culex pipiens L. stages. J. Environ. Sci. Technol. 5, 54–63 (2012).
Chaubal, R. et al. Larvicidal activity of acacia nilotica extracts and isolation of D-Pinitol–A bioactive carbohydrate. Chem. Biodivers. 2, 684–688 (2005).
Abbassy, M., Ibrahim, H. & Gab Alla, M. A. Evaluating the insecticidal and fungicidal efficiency of Acacia nilotica pods extract. J. Plant Protect. Pathol. 9, 283–289 (2018).
Rojas-Sandoval, J. Acacia nilotica (gum arabic tree). (2023).
Vivekanandhan, P. et al. Comparative analysis of major mosquito vectors response to seed-derived essential oil and seed pod-derived extract from Acacia nilotica. Int. J. Environ. Res. Public Health 15, 388 (2018).
Vasudev, A., Kaur, J., Punj, I., Gill, P. K. & Sohal, S. K. Evaluation of methanol and acetone bark extracts from Acacia nilotica (Linn.) as a source of growth inhibitors against Bactrocera cucurbitae (Diptera: Coquillett). J. Entomol. Zool. Stud. 3(4), 260–266 (2015).
Taura, D., Mukhtar, M. & Adoum, O. Lethality of the aqeous extracts of Acacia nilotica, Guiera senegalensis, Kigelia africana and Securidaca longepedunculata on Culex mosquito larva. Ife J. Sci. 6, 115–118 (2004).
Edriss, A. E., Satti, A. A. & Alabjar, Z. A. Preliminary studies on phytochemicals and larvicidal effects of Acacia nilotica L. extracts against Anopheles arabiensis Patton. Sci. Res. Essays 7, 4253–4258 (2012).
Ramesh Babu, S., Subrahmanyam, B., Srinivasan, & Santha, I. In vivo and in vitro effect of Acacia nilotica seed proteinase inhibitors on Helicoverpa armigera (Hübner) larvae. J. Biosci. 37, 269–276 (2012).
Banso, A. Phytochemical and antibacterial investigation of bark extracts of Acacia nilotica. J. Med. Plants Res. 3, 082–085 (2009).
Benelli, G., Kadaikunnan, S., Alharbi, N. S. & Govindarajan, M. Biophysical characterization of Acacia caesia-fabricated silver nanoparticles: Effectiveness on mosquito vectors of public health relevance and impact on non-target aquatic biocontrol agents. Environ. Sci. Pollut. Res. 25, 10228–10242 (2018).
Promsiri, S., Naksathit, A., Kruatrachue, M. & Thavara, U. Evaluations of larvicidal activity of medicinal plant extracts to Aedes aegypti (Diptera: Culicidae) and other effects on a non target fish. Insect Sci. 13, 179–188 (2006).
Patil, C. D., Patil, S. V., Salunke, B. K. & Salunkhe, R. B. Bioefficacy of Plumbago zeylanica (Plumbaginaceae) and Cestrum nocturnum (Solanaceae) plant extracts against Aedes aegypti (Diptera: Culicide) and nontarget fish Poecilia reticulata. Parasitol. Res. 108, 1253–1263 (2011).
Conchou, L. et al. Insect odorscapes: From plant volatiles to natural olfactory scenes. Front. Physiol. 10, 972. https://doi.org/10.3389/fphys.2019.00972 (2019).
Spanoudis, C. G. et al. The chemical code for attracting Culex mosquitoes. Front. Ecol. Evolut. https://doi.org/10.3389/fevo.2022.930665 (2022).
Ahmad, A. et al. Evaluation of different high doses aqueous plant extracts for the sustainable control of Aedes aegypti mosquitoes under laboratory conditions. J. King Saud Univ. Sci. 35, 102991. https://doi.org/10.1016/j.jksus.2023.102991 (2023).
Chengala, L. & Singh, N. Botanical pesticides: A major alternative to chemical pesticides—a review. Int. J. Life Sci 5, 722–729 (2017).
Prabha, S. et al. Biopesticides: An alternative and eco-friendly source for the control of pests in agricultural crops. Plant Arch. 16, 902–906 (2016).
Souto, A. L. et al. Plant-derived pesticides as an alternative to pest management and sustainable agricultural production: Prospects, applications and challenges. Molecules 26, 4835 (2021).
Nawaz, H., Shad, M. A., Rehman, N., Andaleeb, H. & Ullah, N. Effect of solvent polarity on extraction yield and antioxidant properties of phytochemicals from bean (Phaseolus vulgaris) seeds. Braz. J. Pharm. Sci. 56, e17129 (2020).
Baz, M. M. et al. Evaluation of four ornamental plant extracts as insecticidal, antimicrobial, and antioxidant against the West Nile vector, Culex pipiens (Diptera: Culicidae) and metabolomics screening for potential therapeutics. (2023).
Das, N. & Roy, U. S. Studies on phytochemical analysis, antioxidant, antibacterial and Larvicidal properties of the acacia Nilotica fruit extracts. Mapana J. Sci. 21 (2022).
Sadalge, P. R. et al. Synthesis, characterization, and biological activity of novel azole piperazine congeners. J. Appl. Pharm. Sci. 13, 053–069 (2023).
Raman, B. V. et al. Antibacterial, antioxidant activity and GC-MS analysis of Eupatorium odoratum. Asian J. Pharm. Clin. Res. 5, 99–106 (2012).
Ali, M. M., Ramadan, M. A., Ghazawy, N. A., Afify, A. & Mousa, S. A. Photochemical effect of silver nanoparticles on flesh fly larval biological system. Acta Histochem. 124, 151871 (2022).
Haunerland, N. H. & Spener, F. Fatty acid-binding proteins–insights from genetic manipulations. Prog. Lipid Res. 43, 328–349 (2004).
Liu, R.-Z., Li, X. & Godbout, R. A novel fatty acid-binding protein (FABP) gene resulting from tandem gene duplication in mammals: Transcription in rat retina and testis. Genomics 92, 436–445 (2008).
Schaap, F. G., Van der Vusse, G. J. & Glatz, J. F. Evolution of the family of intracellular lipid binding proteins in vertebrates. Cell Lipid Bind. Proteins https://doi.org/10.1007/978-1-4419-9270-3_9 (2002).
Zimmerman, A. & Veerkamp, J. New insights into the structure and function of fatty acid-binding proteins. Cell. Mol. Life Sci. CMLS 59, 1096–1116 (2002).
Marcelino, A. M. C., Smock, R. G. & Gierasch, L. M. Evolutionary coupling of structural and functional sequence information in the intracellular lipid-binding protein family. Prot. Struct. Funct. Bioinform. 63, 373–384 (2006).
Haunerland, N. H., Andolfatto, P., Chisholm, J. M., Wang, Z. & Chen, X. Fatty-acid-binding protein in locust flight muscle: Developmental changes of expression, concentration and intracellular distribution. Eur. J. Biochem. 210, 1045–1051 (1992).
Smith, A. F., Tsuchida, K., Hanneman, E., Suzuki, T. C. & Wells, M. A. Isolation, characterization, and cDNA sequence of two fatty acid-binding proteins from the midgut of Manduca sexta larvae. J. Biol. Chem. 267, 380–384 (1992).
Caccia, S. et al. Functional analysis of a fatty acid binding protein produced by Aphidius ervi teratocytes. J. Insect Physiol. 58, 621–627 (2012).
Author information
Authors and Affiliations
Contributions
Conceptualization, methodology, investigation, data curation, formal analysis, resources, writing-original draft preparation, M.M.B., N.M.E., A.M.A., H.S.G., M.H.A., A.S. and I.T.R.; editing and writing-review, M.M.B., N.M.E., A.M.A., H.S.G., M.H.A., A.S. and I.T.R.; project administration, A.S.; funding achievement, M.M.B., N.M.E., A.M.A., H.S.G., M.H.A., A.S. and I.T.R. All authors have read and approved to the published version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Baz, M.M., El-Shourbagy, N.M., Alkhaibari, A.M. et al. Larvicidal activity of Acacia nilotica extracts against Culex pipiens and their suggested mode of action by molecular simulation docking. Sci Rep 14, 6248 (2024). https://doi.org/10.1038/s41598-024-56690-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-56690-2
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
