
Abstract
Prunus scoparia (Spach) C. K. Schneid is among the most prevalent species which has the potential of being used as a dwarf rootstock for the cultivated almond. In the present study, the phenotypic variation of 521 wild accessions of this species naturally grown in 29 areas of 11 provinces in Iran was assessed. The accessions investigated showed significant differences based on the measured traits. The majority of the characters measured (90 out of 100) exhibited a coefficient of variation of higher than 20.00%, indicating considerable variation among the accessions. The range of nut-related characters was as follows: nut length: 9.72–22.87 mm, nut width: 5.81–15.54 mm, nut thickness: 5.67–12 mm, and nut weight: 0.18–0.99 mm. The range of kernel-related characters was as follows: kernel length: 6.83–19.23 mm, kernel width: 4.28–10.32 mm, kernel thickness: 2.16–7.52 mm, and kernel weight: 0.03–0.37 g. Kernel weight exhibited positive and significant correlations with nut length (r = 0.57), nut width (r = 0.54), nut thickness (r = 0.42), nut weight (r = 0.69), kernel length (r = 0.75), kernel width (r = 0.78), and kernel thickness (r = 0.58). Cluster analysis based on Ward’s method showed two different major clusters among all the accessions. Based on the bi-plot created using principal component analysis of population analysis, the studied 29 natural habitats formed four groups. The studied accessions showed considerable variation in terms of the measured traits within and among populations. This variation is due to cross-pollination, cross-incompatibility, natural hybridization, propagation by seeds, gene flow, and exchange of plant material between the study areas. By using crosses between accessions of different regions, it is possible to increase the amount of variability in different traits of wild almonds.
Similar content being viewed by others


Introduction
Iran is located in arid and semi-arid areas, and more than 60% of this country includes these areas. Wild species are a valuable genetic resource in terms of desired traits in breeding programs, which include traits related to trees and fruits. Accordingly, using plants resistant to such weather conditions should be prioritized1. Also, the valuable features of resistance to biotic and abiotic stresses are evident in these species, which can be used in breeding programs to improve domesticated plants2. The diversity of wild species of fruit trees in Iran is considerable, and there are reserves rich in more almond species than 20 species, whose distribution has been reported here, some of which are endemic2,3.
Some of the wild species of almonds can have high survival in water shortage due to having some characteristics, such as defoliation during the hot season and high ability in absorption and storage of water, and the useful features of drought resistance in them can be evident in breeding programs4. Also, these species grow in shallow and rocky soils, and sometimes they grow in rocks5. Wild species of almonds have been used in Iran since 300 years ago as a rootstock for almonds or related species6. In different regions of Iran, including Hormozgan, Bushehr, Kerman, and Fars provinces, there are many orchards of almonds that are grafted onto wild-related species2. Wild almond species can be used in economics and ecology. Their kernels and oil are used by local people. In addition, wild almond species play a role of physical soil protection and have a high ability to prevent soil erosion3,7.
The resistance of wild almond species to hot and dry weather conditions as well as salinity and cold stress is high8. Other important characteristics of this valuable gene pool are late late-blooming, self-fertilization, and dwarfing9. Therefore, this important genetic resource can be used in future breeding programs, landscape, and reforestation. They can also be used for air purification in polluted areas9,10,11.
One of the wild almond species that is widely distributed throughout Iran is Prunus scoparia (Spach) C. K. Schneid (Fig. 1)12. This wild species has multi-purpose importance, the most important of which is its use as a rootstock for domesticated almonds7. It can also be used to stabilize and prevent soil erosion in arid and semi-arid regions13. High resistance to drought stress and infertile soils, dense and green canopy, beautiful flowers, long-lasting green branches, and long flowering period make this plant a suitable choice for the landscape in arid and semi-arid areas, especially in mountains around cities3,7.

Geographic distribution of P. scoparia in Iran12.
Successful plant breeding programs are highly dependent on their genetic diversity. Investigating and determining genetic diversity is very important. Wild species are valuable genetic resources that are abundant and have a high potential for crop improvement in breeding programs. Therefore, identifying their promising accessions is needed9. Analysis of morphological traits is one of the first steps and the most common methods for germplasm description and identification. In the present study, the phenotypic variation of 521 wild accessions of P. scoparia collected from 29 regions of 11 provinces in Iran was evaluated. The findings of the present study can contribute to effective breeding programs.
Materials and methods
Plant material
The phenotypic variation of 521 wild accessions of P. scoparia collected from 29 regions of 11 provinces in Iran was evaluated for two consecutive years (2021 and 2022). Table 1 contains the geographical characteristics of the studied areas. The identification of the specimens was performed by Prof. Dr. Ali Khadivi. A herbarium voucher specimen with sediment number PS-2443 was donated to a public available herbarium of the Faculty of Agriculture and Natural Resources of Arak University, Iran. Permits required to collect the studied plant samples were obtained from the Ministry of Agriculture and Natural Resources of Iran. For correct sampling, a proper distance of at least 200 m between the accessions of each area was regarded so that the clone samples were not collected.
The characters evaluated
In total, 100 morphological traits related to flowers, branches, leaves, and fruits were recorded using 50 replications for each organ. Traits related to the size and weight of different organs were measured through digital calipers and electronic scales, respectively. To estimate the qualitative attributes in the form of code and rank, the almond descriptor (IPGRI) was used14.
Statistical analysis
The average data were used for analyses. To determine the significance between accessions, analysis of variance was done using SAS software15. The SPSS software16 was used to determine the correlation between the traits as well as principal component analysis (PCA). Cluster analysis based on Ward's method and Euclidean distance and creating a scatter plot based on PC1 and PC2 were done using PAST software17.
Statement specifying permissions
For this study, we acquired permission to collect P. scoparia specimens issued by the Agricultural and Natural Resources Ministry of Iran.
Statement on experimental research and field studies on plants
All methods performed on plants (either cultivated or wild), including the collection of plant material comply with relevant institutional, national, and international guidelines and domestic legislation of Iran.
Results and discussion
The accessions investigated showed significant differences based on the traits (ANOVA, P < 0.01). The majority of the characters (90 out of 100) exhibited a coefficient of variation (CV) of higher than 20.00%, indicating considerable variation among the accessions. The lowest CVs belonged to nut thickness (11.12%), sepal shape (14.43%), nut width (14.43%), current branch leaf apex shape (14.76%), and kernel width (15.57%), while suture opening of the shell showed the highest CV (685%), followed by the marking of the outer shell (171.28%), fruit stigma retention (161.07%), kernel shriveling (136.80%), shell surface pubescent (136.38%), shell ornamentation (133.33%), shell ornamentation present (130.27%), shell back line shape (117.34%), peduncle color (110.96%), and shell short furrows starting from the base (106.38%) (Table 2). Variation in the species and cross-pollination that lead to heterozygosity and increased genetic diversity in almonds during development and evolution can increase the CV value of different traits among accessions18.
Peduncle length ranged from 0.77 to 3.95 mm, while peduncle width varied from 0.95 to 3.36 mm. Petal length varied from 4.30 to 16.37 mm, and petal width ranged from 2.56 to 15.01 mm. Sepal length ranged from 1.50 to 6.66 mm, while sepal width varied between 0.60 and 5.65 mm.
Tree height was moderate (1–2 m) and then low (< 1 m) in the majority of accessions (263 and 125 accessions) (Table 3). In breeding programs, low tree height is considered a useful trait for introducing dwarfing rootstocks7,19.
Tree growth vigor was high in most accessions (259) (Table 3). Trunk diameter was moderate and then high in the majority of accessions (238 and 155, respectively). The stem diameter in P. scoparia is very important for the production of gum and resin. It has been reported that most resin-producing plants form resin-producing ducts for self-defense20. One of the most important reasons that wild almond trees show high resistance to pests and diseases can be attributed to the unique feature of gum production in them21. In addition, hydraulic conductivity in most plant species is enhanced by increasing the thickness of stems and branches. The movement of water and nutrients needed towards the fruit is done better by the thick branches, which increases the growth and quality of the fruit22.
Annual branch color was light green in 356, green in 299, dark green in 64, and brown in 3 accessions. Also, current branch color in summer was predominantly light green (427 accessions) and then green (245), while it was dark green in 31, crimson in 12, and purple-green in 7 accessions. The leaves of P. scoparia normally fall in early summer and then the green branches continue photosynthesis to provide carbohydrates for root and branch growth and development for the rest of the growing season. In this case, the green branches compensate for the lack or absence of leaves. Considering that cytokinins and gibberellins are produced in the root23, the transfer of these substances to the branches, instead of forming new leaves, causes internode elongation in P. scoparia3,4.
The range of related characters of leaves on the annual branch (branch of the previous year) was as follows: leaf length: 8.11–54.35 mm, leaf width: 1.23–10.15 mm, petiole length: 0.60–9.55 mm, and petiole width: 0.25–1.10 mm. The range of related characters of leaves on the current branch was as follows: leaf length: 8.00–40.25 mm, leaf width: 0.74–10.15 mm, petiole length: 0.46–10.31 mm, and petiole width: 0.30–0.91 mm (Table 2). In general, leaf area in P. scoparia is low, or in other words, its leaves are small, which indicates its greater adaptation to drought stress. Previous studies also reported that reduction of leaf area is an initial response of plant adaptation to drought conditions. Considering that the leaves of P. scoparia fall in early summer and the green shoots continue photosynthesis, this species can be a better choice as a rootstock. Also, the pubescence amount on the upper and lower surfaces of P. scoparia leaves is high, which is one of the responses to improve resistance to drought stress24.
Although the leaf size of P. scoparia is smaller, it is interesting that the dry matter content in this species is higher than that of domestic almonds. Dry and fresh weight in a plant determines its biomass production25. Interestingly, the cheapest and easiest method to track the performance and adaptation of plants to drought is to measure the accumulation of ash and mineral content26. It has been reported that plants with more dry matter have higher yields under salinity-stress conditions27. Previously, positive and significant correlations between leaf ash content and yield28,29 and also between leaf life span and dry matter content have been reported30.
The range of fruit stalk length and diameter was 1.85–5.53 mm and 0.88–3.48 mm, respectively. The range of nut-related characters was as follows: nut length: 9.72–22.87 mm, nut width: 5.81–15.54 mm, nut thickness: 5.67–12.00 mm, and nut weight: 0.18–0.99 mm. The suture opening of the shell was absent in 511 out of 521 accessions studied. The well-sealed shell is common in P. scoparia and is reported to be more resistant to fungus and insect infestation31. This offers new opportunities in breeding already not readily available in domesticated almond genetic resources. Shell thickness ranged from 0.40 to 1.76 mm. The presence of a relatively high variation in shell thickness among populations of P. scoparia offers the opportunity to select thin-shell nuts, which is important for the local production of this species as a nut crop32.
The range of kernel-related characters was as follows: kernel length: 6.83–19.23 mm, kernel width: 4.28–10.32 mm, kernel thickness: 2.16–7.52 mm, and kernel weight: 0.03–0.37 g. Empty nuts were observed in 79 out of 521 accessions. Variations in kernel size and the occurrence of empty nuts can be due to variations in humidity and rainfall occurring in the natural habitats. When the almond species are subjected to drought stress, they will start to use the kernel moisture which then results in the shrinking of the kernel and the decrease in nut size3,33. Kester et al.34 observed a highly significant effect of the environment on the occurrence of empty nuts, while Sanchez-Perez et al.35 indicated a diminutive annual variation in this particular trait.
Many the almond characteristics are genetically controlled36. Also, the differences in the characters of accessions of different areas could be mainly because of the wider geographic regions and climatic zones covered in this study. Another reason behind these differences could arise from the variation in climatic conditions, especially in rainfall. The variation in annual precipitation is very common in the arid and semi-arid climate of Iran33. The pictures of leaves, flowers, and nuts of P. scoparia accessions studied are shown in Fig. 2.

The leaves, flowers, and nuts of P. scoparia accessions studied.
There were significant correlations between some characters (Table 4). Sepal length showed positive and significant correlations with peduncle length (r = 0.24), petal length (r = 0.60), petal width (r = 0.54), hypanthium length (r = 0.34), hypanthium diameter (r = 0.55), and sepal width (r = 0.73). Annual branch leaf length exhibited positive and significant correlations with annual branch leaf width (r = 0.43), annual branch petiole length (r = 0.57), and annual branch petiole width (r = 0.48). Current branch leaf length was positively and significantly correlated with current branch leaf width (r = 0.47), current branch petiole length (r = 0.71), and current branch petiole width (r = 0.51), in agreement with the previous findings3,11,19,24,33,37.
Nut weight was positively and significantly correlated with nut length (r = 0.77), nut width (r = 0.84), nut thickness (r = 0.70), and shell thickness (r = 0.44). Kernel weight exhibited positive and significant correlations with nut length (r = 0.57), nut width (r = 0.54), nut thickness (r = 0.42), nut weight (r = 0.69), kernel length (r = 0.75), kernel width (r = 0.78), and kernel thickness (r = 0.58), in agreement with the previous findings3,11,19,24,37,38.
PCA placed the traits in 26 components that explained 72.44% of the total variance. PC1 accounted for 6.10% of the total variance and showed significant correlations with petal length, petal shape, petal width, hypanthium diameter, sepal length, and sepal width. Nut length, nut thickness, nut width, nut weight, kernel weight, kernel width, and kernel length were placed in PC2 and explained 6.06% of the total variance. Tree form, trunk type, tree height, and trunk diameter were placed in PC3 and explained 4.58% of the total variance (Table 5). It has been reported that fruit-related traits are important for distinguishing accessions of almond species9,33.
In the scatter plot, the accessions were widely distributed on the plot level (Fig. 3). The results of the plot showed that the accessions have considerable variation so that a large number were placed outside the oval, which indicates their high differences with other accessions in terms of traits in PC1 and PC2.

Scatter plot for the studied P. scoparia accessions based on PC1/PC2. The symbols represent the accessions of each area in the plot (for an explanation of accession symbols, see Table 1).
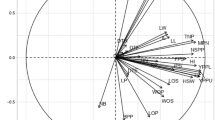
The dendrogram created through Ward's method and Euclidean distance divided the accessions into two groups, each group having several subgroups, which indicates the high variation among accessions (not shown). Also, the studied 29 populations were placed into four groups in the bi-plot generated with PCA of population analysis (Fig. 4). Maharlo, Taft, Hesarsorkh, Bazman, and Esfid populations were placed into the first group, and Khash, Hajiabad, and Rokh populations were placed in the second group. Also, 11 populations, including Chahbot, Sharbabak, Hajiabad, Kalmand, Bahramgoor, Zarju, Barzok, Moorchekhort, Kapise, Kelestan, and Tangestan formed the third group, while the fourth group consisted of the rest 10 populations, including Zavieh, Alvar, Saidabad, Zavarian, Sarrak, Babamir, Shilan, Sargach, Kheirabad, and Shoorab.

Bi-plot for the studied populations of P. scoparia based on the morphological characters.
The studied accessions showed considerable variation in terms of the measured traits within and among populations. This variation is due to cross-pollination, natural hybridization, cross-incompatibility, propagation by seeds, gene flow, and exchange of plant material between the study areas39. The traditional method of propagation and distribution of almonds is through seeds, which has caused the differentiation of traits and increased diversity over time. Also, the exchange of almond germplasm has been done in this country for millennia due to communication and interactions, which has caused interbreeding between populations. Thus, it is possible to justify the grouping of accessions of some distant populations40,41,42. Also, the dissimilarity between accessions of the species denotes the capability of generating new progenies and producing different associations or segregations of genes, thereby facilitating a partial removal of former linkages or the creation of new ones that can be applied in both classical and modern breeding methods. To generate new progenies in a subsequent generation (with new linkage groups or new population properties), it is a common practice to use distant genotypes40,41,42.
Frost resistance is a major breeding goal for almond cultivars in many production areas because of their early flowering time during late winter and early spring. Some accessions of P. scoparia showed late flowering time. The possibility of use of almond related species with a very late-flowering date (high chilling requirements) to develop new cultivars with late-flowering would not only reduce frost damage, but reduce disease damage if flowering is delayed beyond the rainy season, and would allow more efficient use of increasingly scarce insect pollinators43.
Conclusion
A wide range of variations was detected within and among the populations studied of P. scoparia. The obtained results can be important for the management and protection of the gene pool. Also, these findings can be used to develop and introduce new rootstocks for almonds and other stone fruits. The traits, such as late flowering time, suitable trunk diameter, low tree height, suitable tree growth vigor, small leaf size, high nut weight, and high kernel weight are desirable traits that can be considered in almond breeding programs. Also, local cultivation of P. scoparia can be considered for the production of nuts and oil extraction. By using crosses between accessions of different regions, it is possible to increase the amount of variability in different traits of wild almonds.
Data availability
The findings supporting the present study, when reasonable request, are available from the corresponding author.
References
Imani, A. et al. Diversity and broad sense heritability of phenotypic characteristic in almond cultivars and genotypes. Int. J. Hortic. Sci. Technol. 8(3), 261–269 (2021).
Gharaghani, A., Solhjoo, S. & Oraguzie, N. A review of genetic resources of almonds and stone fruits (Prunus spp.) in Iran. Genet. Resour. Crop Evol. 64(3), 611–640 (2017).
Rahimi-Dvin, S., Gharaghani, A., Eshghi, S., Avanzato, D. & Ansari, A. Diversity in the nut and kernel characteristics of seven populations of Prunus scoparia from the central and southern Zagros regions of Iran by comparison with three other almond species. Fruits 72(6), 370–381 (2017).
Madam, B., Rahemi, M., Mousavi, A. & Martines, G. P. Evaluation of the behavior of native Iranian almond species as rootstocks. Int. J. Nuts Relat. Sci. 2(3), 29–34 (2011).
Ali, J. J. M., Nori, I. M., Hama, S. J. & Rashed, S. O. Water harvesting through utilization of wild almond as rootstocks for production of peach, apricot and plum under dry land farming in Sulaymaniyah region. Int. J. Innov. Sci. Eng. Technol. 2(8), 705–724 (2015).
Madani, F. Afforestation trial with different varieties of the almond species in the forestlands of the Kurdistan province—First Phase. In IV International Symposium on Pistachios and Almonds. Acta Horticulturae, Vol 726, 99–104 (2006).
Gharaghani, A. & Eshghi, S. Prunus scoparia, a potentially multi-purpose wild almond species in Iran. Acta Hortic. 1074, 67–72 (2015).
Gholami, M., Rahemi, M. & Kholdebarin, B. Effect of drought stress induced by polyethylene glycol on seed germination of four wild almond species. Aust. J. Basic Appl. Sci. 4(5), 785–791 (2010).
Khadivi-Khub, A. & Anjam, K. Morphological characterization of Prunus scoparia using multivariate analysis. Plant Syst. Evol. 300, 1361–1372 (2014).
Sorkheh, K. et al. Phenotypic diversity within native Iranian almond (Prunus spp.) species and their breeding potential. Genet. Resour. Crop Evol. 56(7), 947–961 (2009).
Moradi, Y., Khadivi, A., Mirheidari, F. & Paryan, S. Morphological variability of naturally grown Prunus scoparia Spach accessions. Sci. Hortic. 267, 109331 (2021).
Yazbek, M. M. Systematics of Prunus Subgenus Amygdalus Monograph and Phylogeny 245 (Cornell University, 2010).
Mozaffarian, V. Trees and Shrubs of Iran 642–664 (Farhang Moaser Publisher, 2005) (in Persian).
Gulcan, R. (ed.) Descriptor List for Almond (Prunus amygdalus) (International Board for Plant Genetic Resources, 1985).
SAS® Procedures. Version 6 3rd edn. (SAS Institute, 1990).
Norusis, M. J. SPSS/PC Advanced Statistics (SPSS Inc., 1998).
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4(1), 9 (2001).
Kester, D. E. & Gradziel, T. M. Almonds. In Fruit Breeding Vol. 3 (eds Janick, J. & Moore, J. N.) 1–97 (Wiley, 1996).
Khadivi-Khub, A. & Anjam, K. Prunus scoparia, a suitable rootstock for almond (Prunus dulcis) under drought condition based on vegetative and fruit characteristics. Sci. Hortic. 210, 220–226 (2016).
Lombardero, M. J., Ayres, M. P. & Ayres, B. D. Effects of fire and mechanical wounding on Pinus resinosa resin defenses, beetle attacks, and pathogens. For. Ecol. Manag. 225(1–3), 349–358 (2006).
Daryaei, M. G., Hoseiny, S. K., Taheri, K., Mirzaei, J. & Mzbani, A. Effect of morphological variables of Pistacia atlantica on gum and seed production. Iran. J. Biol. 25(2), 303–315 (2012).
Genard, M. & Bruchou, C. Multivariate analysis of within-tree factors accounting for the variation of peach fruit quality. Sci. Hortic. 52(1–2), 37–51 (1992).
Arteca, R. N. Plant Growth Substances: Principles and Applications (Springer, 2013).
Khadivi-Khub, A., Sarooghi, F. & Abbasi, F. Phenotypic variation of Prunus scoparia germplasm: Implications for breeding. Sci. Hortic. 207, 193–202 (2016).
Barzegargolchini, B., Movafeghi, A., Dehestani, A. & Mehrabanjoubani, P. Morphological and anatomical changes in stems of Aeluropus littoralis under salt stress. J. Plant Mol. Breed. 5(1), 40–48 (2017).
Cabrera-Bosquet, L., Sánchez, C. & Araus, J. L. How yield relates to ash content, Δ13C and Δ18O in maize grown under different water regimes. Ann. Bot. 104(6), 1207–1216 (2009).
Wright, P. R., Morgan, J. M. & Jessop, R. S. Turgor maintenance by osmoregulation in Brassica napus and B. juncea under field conditions. Ann. Bot. 80(3), 313–319 (1997).
Voltas, J., Romagosa, I., Muñoz, P. & Araus, J. L. Mineral accumulation, carbon isotope discrimination and indirect selection for grain yield in two-rowed barley grown under semiarid conditions. Eur. J. Agron. 9(2–3), 147–155 (1998).
Monneveux, P., Reynolds, M. P., Trethowan, R., Peña, J. & Zapata, F. Carbon isotope discrimination, leaf ash content and grain yield in bread and durum wheat grown under full-irrigated conditions. J. Agron. Crop Sci. 190(6), 389–394 (2004).
Wilson, P. J., Thompson, K. E. N. & Hodgson, J. G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 143(1), 155–162 (1999).
Ledbetter, C. A. & Shonnard, C. B. Evaluation of selected almond (Prunus dulcis (Miller) D.A. Webb) germplasm for several shell and kernel characteristics. Fruit Var. J. 46, 79–82 (1992).
Gradziel, T. M. & Martinez-Gomez, P. Shell seal breakdown in almond is associated with the site of secondary ovule abortion. J. Am. Soc. Hortic. Sci. 127, 69–74 (2002).
Rahimi-Dvin, S., Gharaghani, A. & Eshghi, S. Diversity of nut and seedling characteristics and its relationship to habitat climate in some almond species. J. Nuts 12, 113–136 (2021).
Kester, D. E., Hansche, P. E., Beres, V. & Asay, R. N. Variance components and heritability of nut and kernel traits in almond. J. Am. Soc. Hortic. Sci. 102, 264–266 (1977).
Sanchez-Perez, R., Ortega, E., Duval, H., Martinez-Gomez, P. & Dicenta, F. Inheritance and relationships of important agronomic traits. Euphytica 155, 381–391 (2007).
Arteaga, N., Socias, I. & Company, R. Heritability of fruit and kernel traits in almond. Acta Hortic. 591, 269–274 (2002).
Hashemi, S.-M. & Khadivi, A. Morphological variability of Prunus lycioides Spach germplasm using multivariate analysis. Sci. Hortic. https://doi.org/10.1016/j.scienta.2019.108973 (2020).
Dicenta, F. & Garcia, J. E. Phenotypical correlations among some traits in almond. J. Genet. Breed. 46, 241–246 (1992).
Sefc, K. M., Lopez, M. S., Lefort, F. & Botta, R. Microsatellites variability in grapevine cultivars from different European regions and evaluation of assignment testing to assess the geographic origin of cultivars. Theor. Appl. Genet. 100, 498–505 (2000).
Holsinger, K. E. & Weir, B. S. Genetics in geographically structured populations: Defining, estimating and interpreting F ST. Nat. Rev. Genet. 10(9), 639–650 (2009).
Cruz, C. D. Genes: a software package for analysis in experimental statistics and quantitative genetics. Acta Sci. Agron. 35, 271–276 (2013).
Xiong, J. S., Ding, J. & Li, Y. Genome-editing technologies and their potential application in horticultural crop breeding. Hortic. Res. 2, 15019 (2015).
Rickter, A. A. L’amandier (Academie Sciences Agricoles, Jardin Botanique de Nikits, 1972).
Author information
Authors and Affiliations
Contributions
F.M. and Y.M. experimented and collected data. A.K. guided all stages of the research, analyzed data, and wrote and edited the article. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Khadivi, A., Mirheidari, F. & Moradi, Y. Morphological characterization of wild Prunus scoparia Spach accessions in 11 provinces of Iran. Sci Rep 13, 15864 (2023). https://doi.org/10.1038/s41598-023-43146-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-43146-2
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
