
Abstract
Jatropha curcas is an oilseed crop with biorefinery applications. Whilst cake generated following oil extraction offers potential as a protein source for animal feed, inactivation of toxic phorbol esters present in the material is necessary. Pleurotus pulmonarius is a detoxifying agent for jatropha cake with additional potential as animal feed, edible mushroom and for enzyme production. For the characterization of fungal genes involved in phorbol ester degradation, together with other industrial applications, reverse transcription-quantitative PCR (RT-qPCR) is a tool that enables accurate quantification of gene expression. For this, reliable analysis requires reference genes for normalization of mRNA levels validated under conditions employed for target genes. The stability of potential reference genes β-TUB, ACTIN, GAPDH, PHOS, EF1α, TRPHO, LAC, MNP3, MYP and VP were evaluated following growth of P. pulmonarius on toxic, non-toxic jatropha cake and a combined treatment, respectively. NormFinder and geNorm algorithms for expression stability analysis identified PHOS, EF1α and MNP3 as appropriate for normalizing gene expression. Reference gene combinations contrasting in ranking were compared following normalization of relative expression of the CHU_2040 gene, encoding an esterase enzyme potentially involved in phorbol ester degradation. The reference genes for P. pulmonarius will facilitate the elucidation of mechanisms involved in detoxification of phorbol esters as well as analysis of target genes for application in biorefinery models.
Similar content being viewed by others


Introduction
The current global drive to reduce the negative impacts of industry on the environment has led to the development of the bioeconomy concept, where economic sectors are adapted towards sustainability utilizing renewable biological resources for food, materials and energy production. Agro-industries are undergoing a transformation regarding renewable raw materials, low-carbon production, and land-use optimisation within integrative models such as biorefineries. In this context, Jatropha curcas L., an oilseed crop with potential industrial and agricultural applications, is recognized primarily as a potential sustainable biofuel crop1,2,3. Difficulties exist, however, in adding value to the by-product of jatropha cake, which is a lignocellulosic residue rich in proteins, nitrogen, phosphorus, potassium, and carbon. Application of this residue in animal feed is currently limited due to the presence of toxic and thermostable antinutritional factors such as phorbol ester diterpenes4,5,6.
Terpenes are chemical compounds derived from secondary plant metabolism that play a fundamental role in plant development, ecological interactions and defense responses against pests and pathogens7,8. These chemical compounds, whilst widely used in industrial processes such as in the production of cosmetics, pharmaceuticals, medicinal products, and insecticides7,9,10, are also among the main pollutants accumulating from industrial pulp and paper production11. Given this, the bioprospecting of microorganisms capable of degrading such substances and their derivatives is of fundamental importance in bioremediation. The bio-detoxification of diterpene phorbol esters in jatropha cake offers potential for downstream application of the residue as a fertilizer or animal feed supplement. Appropriate micro-organisms may enable not only the degradation of toxic compounds present, but potentially also concomitantly increase nutritional value, or generate other value-added by-products such as enzymes, bioactive compounds and/or edible mushrooms. The genes and metabolic pathways underlying the biodegradation mechanism of phorbol ester in the cake generated after J. curcas oil extraction are currently poorly explored. However, it has been demonstrated that specific microbial growth can lead to the secretion of extracellular enzymes such as esterases, lipases, and proteases, which play roles in the degradation of the toxic diterpene3,12.
Fungal species of the genus Pleurotus are today employed in the bioeconomy, in green chemistry and in the production of edible protein substitutes. These cultivated mushrooms, known as oyster mushrooms, are among the most cultivated globally13. Success in production is mainly due to the efficiency of fruiting and cultivation on various substrates14. Biotechnological products are numerous, with applications in bioremediation, mycoremediation, biotreatment and enrichment of lignocellulosic waste, biological control, development of catalytic products and in production of polysaccharides such as ergosterol15,16,17,18. Among the oyster mushrooms, Pleurotus pulmonarius is efficient in the degradation of phorbol esters in jatropha cake, with additional potential for application in biorefinery models3. The identification of genes in this basidiomycete fungus that are involved in the degradation of phorbol esters has yet to be conducted and represents an important step towards efficient enzymatic degradation and solving bottlenecks related to biodegradation. In this context, high-throughput sequencing approaches for analysis of the fungal transcriptome will be appropriate for the identification of expressed genes and metabolic pathways activated during the degradation of this toxic compound, as well as additional genes encoding and regulating the expression of enzymes with diverse industrial application3,12.
For accurate quantification and validation of gene expression data originating from high-throughput transcriptome sequence data, reverse transcription-quantitative PCR (RT-qPCR) is a benchmark analytical tool for application across different experimental conditions19,20,21,22,23,24. Robust analysis, however, requires the employment of a suitable set of reference genes for transcript expression normalization, correcting data for variation across samples. In accordance with the Minimum Information for Publication of Real-Time Quantitative PCR Experiments (MIQE) guidelines25, normalization of expression of each target gene in relation to a reference gene is required to correct data for potential variations across sample replicates that often occur during cDNA preparation. For such normalization, specific reference genes are necessary for each organism that consider microbial growth conditions, tissue type and expression time points25,26,27. A suitable reference gene for gene expression normalization should exhibit constant expression, independent of the sample, condition, treatment, cell, and tissue type28,29. To determine the most stable normalizing genes, different algorithms with distinct statistical methods are generally applied, which will determine the best sets of stable genes30,31,32.
The genetic stability of reference genes has previously been investigated in different species of the genus Pleurotus, including Pleurotus ostreatus19,33,34,35; Pleurotus eryngii36, and Pleurotus tuber-regium37. Given the potential in P. pulmonarius for both the degradation of phorbol esters and concomitant application in biorefinery models using jatropha cake as carbon source, analysis of fungal gene expression is warranted. For accurate future analysis of target gene expression, reference genes for RT-qPCR are therefore required for this species. This study investigates the reliability of potential reference genes for P. pulmonarius during growth on jatropha cake, in the presence and absence of toxic phorbol esters.
Results
Specificity and efficiency analysis of primers
Fifteen candidate genes were tested for specificity and stability as potential reference genes for P. pulmonarius cultivated on jatropha cake in the presence and absence of phorbol esters. Primer information for each evaluated gene is described in Supplementary Table S1. Out of the initial 15 candidate genes, five genes, namely PEP, ACTIN2, LAC2, CHS, and α-TUB did not display specific amplification via RT-PCR and were therefore not subsequently tested via RT-qPCR. For the remaining 10 genes, the specificity of primer amplification for single gene loci was confirmed by analysing melting curves, as shown in Supplementary Fig. S1, with single peaks obtained for each primer pair. PCR amplification efficiency values for the candidate genes ranged from 1.956 to 1.85, following calculation with the software LinRegPCR (Table 1).
Analysis of Cq values of candidate reference genes
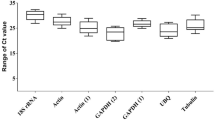
Variation in Cq (cycle quantification) values for the 10 candidate genes was analysed and presented using a Box-plot representation, where it is possible to observe expression intervals, average Cq values (mean and median) and discrepant outlier values. Analysis of each gene was carried out under distinct conditions, namely P. pulmonarius grown on jatropha cake with the presence of toxic phorbol ester (treatment T) (Fig. 1A), P. pulmonarius grown on jatropha cake without toxic phorbol ester (treatment NT) (Fig. 1B), and via consideration of P. pulmonarius under both conditions (referred to as a combined treatment (C), representing a dataset of combined T and NT data) (Fig. 1C). In the presence of toxic phorbol esters, Cq values ranged from 16.9 to 29.1 across the candidate reference genes. Under this condition, the ACTIN gene presented the lowest number of cycles, indicating greater expression than the other candidate genes, whilst the peroxidase (VP) gene presented the highest Cq value, indicating lowest gene expression. Greatest variation of Cq was observed for the gene VP, with GAPDH presenting the lowest variation of Cq, with a mean value and median similar to the EF1α gene. The LAC gene was the only candidate to present outliers for this condition. For the NT condition of P. pulmonarius cultivation on jatropha cake without toxic phorbol esters, Cq values ranged from 15.6 to 30.8. For this condition, the MYP gene displayed the highest Cq value, whilst the lowest Cq value was observed for the ACTIN gene, where the highest variation of Cq was also observed. The gene VP was the only candidate in this condition to present outliers, with the lowest variation of Cq observed for the GAPH gene. For the combined condition, ACTIN and VP genes showed the lowest and highest Cq values, respectively. Both genes, however, showed a large variation in Cq, as also observed for the MYP gene. The lowest variation of Cq for this condition was observed for GAPDH and EF1α genes.

Box plot presentation of the variations in the Cq values for the ten candidate genes tested for stabilizing gene expression of Pleurotus pulmonarius cultivated on toxic and non-toxic jatropha cake. (A) Toxic, (B) non-toxic, (C) combined.
Expression stability analysis
Stability of candidate reference gene expression stability for P. pulmonarius cultivated on jatropha cake representing T, NT and combined data for treatments was assessed using the algorithms BestKeeper, geNorm, and NormFinder. Ranking of expression stability according to each biological treatment is summarized in Table 2.
Analysis with the geNorm algorithm revealed stability for the P. pulmonarius candidate genes β-TUB, PHOS, EF-1 and MNP3 across all treatment conditions, with stability values below the default limit of 0.5, representing stability (Fig. 2). In contrast, the VP gene showed values above 0.5 across all the treatments. For MYP, TRPHO and GAPDH genes, stability was observed only during fungal growth during treatment T, with values observed of 0.11, 0.12, and 0.472, respectively. The LAC and ACTIN genes displayed stability only during treatment NT, with values of 0.116 and 0.394, respectively. Pairwise variation analysis (V) was conducted using geNorm to determine the minimum number of genes necessary for normalizing gene expression in P. pulmonarius on J. curcas cake. Data presented in Fig. 3 shows a pairwise variation value of V2/3 of V < 0.15 for all fungal growth conditions, indicative that two reference genes are sufficient for normalization of gene expression across the treatments.

GeNorm algorithm-derived data on the stability of candidate reference genes for normalizing gene expression via qPCR in Pleurotus pulmonarius under the three analysed conditions: toxic, non-toxic, and combined. Values of M below a 0.5 cut-off indicate a high stability rate.

GeNormV-based determination of optimal numbers of reference genes for accurate RT-qPCR normalization of target gene expression. For all data values below a cut-off value of 0.15, additional reference genes do not contribute significantly to gene expression data normalization.
Gene stability ranking data following analysis with the algorithm NormFinder is presented in Fig. 4. In agreement with ranking data obtained using geNorm, PHOS and EF1α genes displayed greatest stability across all treatment conditions. In the case of the MNP3 gene, analysis with NormFinder indicated high stability in expression in P. pulmonarius during cultivation conditions on toxic jatropha cake and when considering data for the combined conditions. Also in agreement with geNorm data, the genes TRPHO and MYP, displayed greater stability during growth under only the toxic condition, whilst β-TUB, ACTIN and LAC genes, which also displayed stability only during the toxic condition, showed a NormFinder stability profile diverging from geNorm ranking data.

NormFinder algorithm-derived data on the stability of candidate reference genes for normalizing gene expression via qPCR in Pleurotus pulmonarius under the three analysed conditions: toxic, non-toxic, and combined. Values of M below a 0.5 cut-off indicate a high stability rate.
Analysis using the program BestKeeper revealed only the GAPDH gene displaying a standard deviation value lower than 1 for P. pulmonarius under the three distinct conditions, and the EF1α gene displaying a value of 0.94 in P. pulmonarius during treatment NT (Table 2). Results were generally discrepant in relation to those obtained with the algorithms geNorm and NormFinder, with the exception of the gene EF1α, which showed stability with all three algorithms employed.
Testing selected reference genes for normalization of a P. pulmonarius esterase gene
To validate reference gene rankings, relative expression of the CHU_2040 gene was normalized against different reference gene combinations. This gene encodes an esterase enzyme, potentially involved in phorbol ester degradation. Analysis of RNA-seq data from P. pulmonarius on J. curcas (unpublished), has revealed that the CHU_2040 gene exhibits differential expression across 3, 7, and 11 days of cultivation on toxic jatropha cake when compared to cultivation on non-toxic cake. For normalization tests using the CHU_2040 gene, various combinations of genes ranked as most stable (PHOS and EF1α; EF1α and MNP3) and as least stable (LAC and VP) were compared. Figure 5 demonstrates that normalizing the relative expression of the CHU_2040 gene using the two most stable gene pair combinations revealed a similar positive expression profile of the gene over the time course investigated, albeit with variations in expression levels, and with RT-qPCR data supporting in silico data. Normalization with the two least stable gene candidates, in contrast, resulted in distinct gene expression profiles of the CHU_2040 gene over the time course, with greater standard deviation.

Relative expression levels of the esterase gene CHU_2040 in Pleurotus pulmonarius using the most (EF1α and PHOS; EF1α and MNP3) and the least (LAC and VP) stable pairs of reference genes. RT-qPCR was performed with three biological replicates, considering the combined condition C (datasets for P. pulmonarius cultivated on toxic and non-toxic jatropha cake). (A) EF1α and PHOS; (B) EF1α and MNP3; (C) LAC and VP.
Discussion
Given the potential application of P. pulmonarius in the degradation of phorbol esters and concomitant enrichment of lignocellulosic residues3, the objective of this study was to identify reference genes for normalizing gene expression data for P. pulmonarius cultivated on jatropha cake in the presence and absence of the toxic diterpene phorbol ester.
Several potential reference genes were selected for screening in P. pulmonarius during growth on toxic and non-toxic jatropha cake. These encoded proteins involved in constitutive fungal activities, such as β-tubulin and Actin 1, which are both involved in structural activity, GAPDH, which is involved in glycolysis, PHOS, that plays an important role in the synthesis of amino acids and nitrogenous bases, EF1α, involved in translation processes, and TRPHO, involved in membrane protection19,38,39,40.
The mathematical algorithms of geNorm28, NormFinder29 and BestKeeper41 have been widely employed for reference gene development across different kingdoms, with combined data generally enabling accurate identification of stable normalizing genes. Here, data obtained using NormFinder and geNorm showed an overlap for the most stable and the least stable genes, in contrast to the results obtained using BestKeeper. Discrepancies in results obtained with the different software are common19. BestKeeper is an algorithm that may be inadequate for in vivo samples, since the calculation model applied (geometric mean of reference genes with a standard deviation > 1) does not fit biological system variations and is limited to controlled conditions23,42.
Based on the results obtained using geNorm and NormFinder, highest stability among the housekeeping genes evaluated was observed with the genes PHOS and EF1α. According to Castanera and coworkers19, the PHOS gene also showed high stability and is suitable for normalizing gene expression in P. ostreatus using the same algorithms. The EF1α gene was recently introduced as the secondary barcode region for molecular identification of fungi at the species level43, including basidiomycetes of the genus Pleurotus44,45. As such, this gene also adheres to the guidelines for ideal reference genes, with conservation in the target DNA sequence, species specificity, constant copy number, and without allelic variation36. Previously, the GAPDH gene was reported to be unsuitable for normalizing expression in the basidiomycete P. tuber-regium, according to data generated using the geNorm algorithm37. Here, the GAPDH gene was also shown to be unsuitable for normalizing gene expression in P. pulmonarius cultivated on jatropha cake, as was the ACTIN gene. Castanera and colleagues19 reported that the constitutive genes ACTIN, GAPDH1, and β-TUB, according to results generated using Genorm and NormFinder, were the least stable for normalizing gene expression in P. ostreatus cultivated on lignocellulosic residue. Clearly, the use of constitutive genes as reference genes without validation of their stability can lead to errors in gene expression analyses37,46. Given the previous reports across the three Pleurotus species, the constitutive gene GAPDH does not appear to be adequate for normalizing the expression of genes in basidiomycetes of the genus Pleurotus.
Pleurotus pulmonarius belongs to the class of saprotrophic basidiomycetes, fungi that have evolved the ability to degrade the complex polymers that make up plant cell walls47,48. Within the basidiomycetes, P. pulmonarius is classified as a White-rot fungus, capable of completely degrading lignin through the action of oxidative enzymes48,49,50,51. Laccase (LAC), manganese peroxidase (MnP), lignin peroxidase (LiP), and versatile peroxidase (VP) are important enzymes in lignin degradation and are assumed to be the first in the line of proteins expressed during fungal catabolism of lignin49. Members of the genus Pleurotus have been reported to possess genes encoding MNP and VP, but not LiP-encoding genes52. Given the importance of oxidative enzymes for saprotrophic basidiomycetes, genes encoding LAC, MnP, and VP proteins were also tested for here for gene stability. The literature is scarce regarding the selection of reference genes for P. pulmonarius, although the MYP gene has been described for QPCR quantification of P. pulmonarius mycelia53. In our results, the MYP gene showed stability only under toxic culture conditions. In addition, this gene showed the highest Cq value in both toxic and non-toxic conditions, suggesting that it may be less expressed in these conditions. The LAC and VP genes did not show stability during any of the tested conditions, with box-plot data indicating outliers within the analyses. In contrast to the other genes encoding oxidative enzymes, the MNP3 gene, encoding a distinct manganese peroxidase to that of the MYP gene, displayed high stability under the experimental conditions, as did the PHOS and EF1α genes.
According to Phengnuam and Suntornsuk54 and Nakao et al.55, esterases are the primary enzymes involved in the degradation of phorbol esters by bacteria of the genus Bacillus. Based on in-house RNA-seq data for P. pulmonarius on toxic and non-toxic jatropha cake (unpublished), the CHU_2040 gene encodes an esterase potentially involved in the complete degradation of phorbol esters. Through comparison with RNA-seq data, relative expression of this gene was used to validate the stability of the reference genes. When the most highly ranked stable gene combinations (PHOS-EF1α and EF1α-MNP3) were applied for normalizing relative expression of the CHU_2040 gene, differential expression results aligned with the in silico data. In contrast, when the least stable reference genes for normalization were employed, namely LAC and VP, the expression pattern of CHU_2040 exhibited a distinct profile that significantly diverged from the in silico expression data, revealing their unsuitability as normalization genes under the specific growth conditions employed for P. pulmonarius. These results clearly showed that different reference genes can influence the relative expression of target genes under specific growth conditions.
In summary, this first investigation into the development of reference genes for P. pulmonarius identified three stable genes suitable for normalizing gene expression during culture on jatropha cake with or without the toxic compound phorbol ester. In addition, paired variation analyses showed that gene normalization can be performed with two reference genes. Whilst P. pulmonarius offers considerable importance in biotechnology, data related to molecular investigation of this fungus is still relatively limited. Together with the recent reference genome for P. pulmonarius16, the set of reference genes developed here for gene expression normalization is appropriate for future transcriptomic analysis and validation of gene expression of this fungus on this lignocellulosic residue.
Methods
Jatropha cake material
Cakes were obtained by mechanically pressing seeds for genetic accessions of non-toxic (accession 169) and toxic J. curcas (accession 123), all provided by the germplasm bank at Embrapa Agroenergia. Prior to pressing, seeds were individually heated at 60 °C for two hours using a rotary mixer dryer (SCOLT TECH, ERT 60 II). The pressed seeds were then processed using an oil and grease extractor (SCOLT TECH, SMR 610-6) to obtain both crude oil and the cakes that were employed in this study. Methanolic extraction of phorbol esters and quantification by HPLC was conducted following the method described by Gomes et al.3. The concentration of phorbol esters in the toxic cake was measured at 2.17 mg/g, while the non-toxic cake did not contain detectable levels of phorbol esters using the employed technique.
Fungal material and bioassays
Pleurotus pulmonarius strain BRM 055674, originating from the basidiomycete collection at Embrapa Agroenergia, was preserved according to the Castellani method56. Subsequently, mycelial disks were transferred to Jatropha agar culture medium3 and incubated at 28 °C for 7 days. As described above, available naturally non-toxic varieties of J. curcas57 that do not synthesize or have minimal amounts of phorbol esters were employed in bioassays as controls for analysis of gene expression modulation after fungal growth on toxic jatropha cake. For solid-state cultivation, 100 g of each substrate of toxic and non-toxic jatropha cake were moistened to 70% humidity and transferred to 150 mm × 20 mm Petri dishes. Following sterilization, 7-day old P. pulmonarius mycelial plugs were inoculated onto either toxic jatropha cake (treatment T) or non-toxic jatropha cake (treatment NT), then incubated at 28 °C. Fungal mycelia colonizing the substrates were collected at three time points over a 3-, 7- and 11-day incubation period. All bioassays were performed in triplicate. As described above, the third condition analysed corresponded to a combination of T and NT treatment data sets and is referred to as the combined condition throughout (treatment C).
RNA extraction and cDNA synthesis
Mycelial samples of P. pulmonarius were collected from the surface of colonized jatropha cake for both treatments at 3, 7 and 11 days after inoculation (DAI). Mycelia was immediately flash frozen in liquid nitrogen then stored at – 80 °C. Extraction and purification of total RNA for each treatment was performed using an INVITRAP® SPIN PLANT RNA kit (Stratec Molecular GMBH, Berlin, Germany), following the manufacturer’s instructions. Total RNA was treated with DNase I (New England Biolabs, Ipswich, MA, USA) for removal of residual genomic DNA. Total RNA concentration and integrity were analysed via 1% agarose gel electrophoresis and Nanodrop ND-1000 spectrophotometry (Thermo Scientific, Waltham, MA, USA). For synthesis of cDNA, from biological triplicates for each experimental condition (3DAI_NT, 3DAI_T, 7DAI_NT, 7DAI_T, 11DAI_NT, 11DAI_T), a total of 1 μg of total RNA was reverse transcribed to cDNA using Super Script II RT and Oligo(dT) primers (Invitrogen, Carlsbad, CA, USA).
Primer design and selection
To select potential reference genes with stable expression in P. pulmonarius grown on jatropha cake waste, candidate genes were selected for screening that encode proteins likely involved in basal cell activities across diverse species of the genus Pleurotus (Supplementary Table S1). The software PrimerQuest Tool (Integrated DNA Technologies—IDT, Iowa, EUA) was employed for the design of specific primers for each candidate reference gene. Expected target amplicons varied from 90 to 120 bp, with each primer pair designed towards predicted exon-exon junctions, in order to avoid amplification from genomic DNA and to ensure amplification from cDNA. The specificity of primer pairs was initially tested by in-silico PCR against a local database of RNAseq data for P. pulmonarius. Primer specificity and efficiency was then tested against the cDNA originating from the biological triplicates of each experimental condition for P. pulmonarius cultivated on toxic and non-toxic jatropha cake.
Reverse transcription-quantitative PCR
A Platinum SYBR Green qPCR Super Mix-UDG w/ROX kit (Invitrogen, Carlsbad, CA, USA) was employed for RT-qPCR analysis of expression of potential reference genes. PCR amplifications were performed using an ABI StepOne® Real-Time PCR thermocycler (Applied Biosystems, Foster City, USA). PCR amplification was conducted on three experimental replicates and three technical replicates per treatment. Each PCR reaction comprised 2 μL of a 1:20 dilution of template cDNA, 0.2 μM of each primer and 5 μL Platinum® SYBR® Green qPCR Super Mix-UDG w/ROX kit (Invitrogen, Carlsbad, CA, USA), to a final volume of 10 μL. PCR thermocycling comprised 52 °C for 2 min, 95 °C for 10 min, followed by 40 cycles of denaturation at 95 °C for 15 s, followed by primer annealing and extension at 62 °C for 60 s. Analysis of the Tm (dissociation) of amplicons was performed using the software SDS 2.2.2 (Applied Biosystems, Foster City, USA) to verify specificity of primers. Raw ΔRn data was applied to determine RT-qPCR efficiency for each gene by using the program LinRegPCR, version 2017.1 (https://www.gear-genomics.com/rdml-tools/).
Analysis of expression stability
Stability analysis and validation of expression for each candidate reference gene were based on quantification cycle (Cq) values for each cDNA sample. This value indicates the total number of amplification cycles during the PCR exponential amplification phase that are needed to reach a default threshold value for amplification detection. The expression stability of each gene was determined using the analytical algorithms geNorm28, NormFinder29 and BestKeeper41. GeNorm was also employed to calculate expression stability based on an average M-value, representing the pairwise variation of a particular gene against all other genes. Greatest stability in expression is represented by the lowest M-values, with the most stable genes displaying M-values below a threshold of 0.5. GeNorm also calculates the pairwise variation V-value (Vn/n + 1) between each potential reference gene, for determination of the optimal number of reference genes for employment in gene expression normalization. Subsequently, the software qbase + (CellCarta, Montreal, Quebec) was used to calculate the average quantification cycles (Cqs) per gene and the software GraphPad Prism v7 (Dotmatics, Boston, EUA) for statistical analysis.
Expression analysis of CHU_2040
Combinations of the most stable genes (PHOS and EF1α; MNP3 and EF1α) and the least stable reference genes (LAC and VP) were compared for normalization of the relative expression of the CHU_2040 gene, which encodes an esterase enzyme, potentially involved in phorbol ester degradation. RNA extraction, cDNA synthesis and qPCR were all conducted as previously described. The software qbase + (Biogazelle) was employed to calculate the average quantification cycles (Cqs) per gene, with the software GraphPad Prism v7 employed for statistical analysis of the combined condition.
Data availability
All data generated and analysed during the study is included in the published article and its Supplementary Information files.
References
Giwa, A. et al. Techno-economic assessment of the sustainability of an integrated biorefinery from microalgae and Jatropha: A review and case study. Renew. Sustain. Energy Rev. 88, 239–257 (2018).
Piloto-Rodríguez, R. et al. An approach to the use of Jatropha curcas by-products as energy source in agroindustry. Energ. Sourc. Part A 2020, 1–21 (2020).
Gomes, T. G. et al. Phorbol ester biodegradation in Jatropha curcas cake and potential as a substrate for enzyme and Pleurotus pulmonarius edible mushroom production. Biocatal. Agric. Biotechnol. 45, 102498 (2022).
Makkar, H., Becker, K., Sporer, F. & Wink, M. Studies on nutritive potential and toxic constituents of different provenances of Jatropha curcas. J. Agric. Food Chem. 45, 3152–3157 (1997).
Makkar, H. P. & Becker, K. Jatropha curcas, a promising crop for the generation of biodiesel and value-added coproducts. Eur. J. Lipid Sci. Technol. 111, 773–787 (2009).
Devappa, R. K., Makkar, H. P. & Becker, K. Jatropha toxicity—a review. Part B Crit. Rev. J. Toxicol. 13, 476–507 (2010).
Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 9, 297–304 (2006).
Jia, Q. et al. Microbial-type terpene synthase genes occur widely in nonseed land plants, but not in seed plants. Proc. Natl. Acad. Sci. 113, 12328–12333 (2016).
Maurya, R., Ravi, M., Singh, S. & Yadav, P. P. A review on cassane and norcassane diterpenes and their pharmacological studies. Fitoterapia 83, 272–280 (2012).
Zerbe, P. et al. Diterpene synthases of the biosynthetic system of medicinally active diterpenoids in Marrubium vulgare. Plant J. 79, 914–927 (2014).
Vilanova, C., Marin, M., Baixeras, J., Latorre, A. & Porcar, M. Selecting microbial strains from pine tree resin: Biotechnological applications from a terpene world. PLoS ONE 9, e100740 (2014).
Gomes, T. G. et al. Current strategies for the detoxification of Jatropha curcas seed cake: A review. J. Agric. Food Chem. 66, 2510–2522 (2018).
Wang, Q. et al. The yield, nutritional value, umami components and mineral contents of the first-flush and second-flush Pleurotus pulmonarius mushrooms grown on three forestry wastes. Food Chem. 397, 133714 (2022).
Mahari, W. A. W. et al. A review on the valorization of oyster mushroom and waste generated in the mushroom cultivation industry. J. Hazard. Mater. 400, 123156 (2020).
Kaewlaoyoong, A. et al. White rot fungus Pleurotus pulmonarius enhanced bioremediation of highly PCDD/F-contaminated field soil via solid state fermentation. Sci. Total Environ. 738, 139670 (2020).
Vidal-Diez-de-Ulzurrun, G., Lee, Y.-Y., Stajich, J. E., Schwarz, E. M. & Hsueh, Y.-P. Genomic analyses of two Italian oyster mushroom Pleurotus pulmonarius strains. G3 11, jkaa007 (2021).
Yunan, N. A. M., Shin, T. Y. & Sabaratnam, V. Upcycling the spent mushroom substrate of the grey oyster mushroom Pleurotus pulmonarius as a source of lignocellulolytic enzymes for palm oil mill effluent hydrolysis. J. Microbiol. Biotechnol. 6, 823–832 (2021).
Chen, F., Xiong, S., Sundelin, J., Martín, C. & Hultberg, M. Potential for combined production of food and biofuel: Cultivation of Pleurotus pulmonarius on soft-and hardwood sawdusts. J. Clean. Prod. 266, 122011 (2020).
Castanera, R., López-Varas, L., Pisabarro, A. G. & Ramírez, L. Validation of reference genes for transcriptional analyses in Pleurotus ostreatus by using reverse transcription-quantitative PCR. Appl. Environ. Microbiol. 81, 4120–4129 (2015).
Devonshire, A. S. et al. Application of next generation qPCR and sequencing platforms to mRNA biomarker analysis. Methods 59, 89–100 (2013).
Galiveti, C. R., Rozhdestvensky, T. S., Brosius, J., Lehrach, H. & Konthur, Z. Application of housekeeping npcRNAs for quantitative expression analysis of human transcriptome by real-time PCR. RNA 16, 450–461 (2010).
VanGuilder, H. D., Vrana, K. E. & Freeman, W. M. Twenty-five years of quantitative PCR for gene expression analysis. Biotechniques 44, 619–626 (2008).
Albuquerque, G. M. et al. Stability analysis of reference genes for RT-qPCR assays involving compatible and incompatible Ralstonia solanacearum-tomato ‘Hawaii 7996’interactions. Sci. Rep. 11, 18719 (2021).
Rego, E. C. S. et al. Stable reference genes for RT-qPCR analysis of gene expression in the Musa acuminata-Pseudocercospora musae interaction. Sci. Rep. 9, 1–11 (2019).
Bustin, S. A. et al. MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622 (2009).
Bustin, S., Benes, V., Nolan, T. & Pfaffl, M. Quantitative real-time RT-PCR—a perspective. J. Mol. Endocrinol. 34, 597–601 (2005).
Gutierrez, L. et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 6, 609–618 (2008).
Vandesompele, J. et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 3, 1–12 (2002).
Andersen, C. L., Jensen, J. L. & Ørntoft, T. F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64, 5245–5250 (2004).
Wang, Q. et al. Stability of endogenous reference genes in postmortem human brains for normalization of quantitative real-time PCR data: Comprehensive evaluation using geNorm, NormFinder, and BestKeeper. Int. J. Legal Med. 126, 943–952 (2012).
Jacob, F. et al. Careful selection of reference genes is required for reliable performance of RT-qPCR in human normal and cancer cell lines. PLoS ONE 8, e59180 (2013).
De Spiegelaere, W. et al. Reference gene validation for RT-qPCR, a note on different available software packages. PLoS ONE 10, e0122515 (2015).
Zheng, S., Shan, L., Zhuang, Y. & Shang, Y. Identification of pyrG used as an endogenous reference gene in qualitative and real-time quantitative PCR detection of Pleurotus ostreatus. J. Food Sci. 83, 750–755 (2018).
Hou, Z. et al. Selection of reference genes for real-time quantitative PCR of Pleurotus ostreatus under heat stress. Acta Edulis Fung. 26, 11–18 (2019).
Garrido-Bazán, V. et al. Effect of textile dyes on activity and differential regulation of laccase genes from Pleurotus ostreatus grown in submerged fermentation. AMB Express 6, 1–9 (2016).
Wei, Y. et al. Identification of s9ap used as an endogenous reference gene in qualitative and real-time quantitative PCR detection of Pleurotus eryngii. Mol. Biol. Rep. 50, 621–629 (2023).
Sun, X., Wu, J., Mo, C. & Ma, A. Selection and validation of endogenous reference genes for RT-qPCR normalization in different stresses and tissues of the tiger milk mushroom, Pleurotus tuber-regium. Mycoscience 62, 281–288 (2021).
Guranowski, A. & Paszewski, A. Metabolism of 5′-methylthioadenosine in Aspergillus nidulans an alternative pathway for methionine synthesis via utilization of the nucleoside methylthio group. Acta Gen. Subj. 717, 289–294 (1982).
Negrutskii, B. & El-Skaya, A. Eukaryotic translation elongation factor 1α: Structure, expression, functions, and possible role in aminoacyl-tRNA channeling. Prog. Nucleic Acid Res. Mol. Biol. 60, 47–78 (1998).
Luo, Y., Li, W.-M. & Wang, W. Trehalose: Protector of antioxidant enzymes or reactive oxygen species scavenger under heat stress?. Environ. Exp. Bot. 63, 378–384 (2008).
Pfaffl, M. W., Tichopad, A., Prgomet, C. & Neuvians, T. P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Environ. Exp. Bot. 26, 509–515 (2004).
Hibbeler, S., Scharsack, J. P. & Becker, S. Housekeeping genes for quantitative expression studies in the three-spined stickleback Gasterosteus aculeatus. BMC Mol. Biol. 9, 1–10 (2008).
Hoang, M. T. V. et al. Dual DNA barcoding for the molecular identification of the agents of invasive fungal infections. Front. Microbiol. 10, 1647 (2019).
He, X.-L. et al. Intra-and inter-isolate variation of ribosomal and protein-coding genes in Pleurotus: Implications for molecular identification and phylogeny on fungal groups. BMC Microbiol. 17, 1–9 (2017).
He, X.-L. et al. Phylogenetic relationship of two popular edible Pleurotus in China, Bailinggu (P. eryngii var. tuoliensis) and Xingbaogu (P. eryngii), determined by ITS, RPB2 and EF1α sequences. Mol. Biol. Rep. 43, 573–582 (2016).
de Oliveira, L. A. et al. Reference genes for the normalization of gene expression in Eucalyptus species. Plant Cell Physiol. 53, 405–422 (2012).
Boddy, L. & Heilmann-Clausen, J. Chapter 12 Basidiomycete community development in temperate angiosperm wood. In British Mycological Society Symposia Series (ed. Boddy, L.) 211–237 (Elsevier, 2008).
Velázquez-Cedeño, M. A., Mata, G. & Savoie, J.-M. Waste-reducing cultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffee pulp: Changes in the production of some lignocellulolytic enzymes. World J. Microbiol. Biotechnol. 18, 201–207 (2002).
Janusz, G., Kucharzyk, K. H., Pawlik, A., Staszczak, M. & Paszczynski, A. J. Fungal laccase, manganese peroxidase and lignin peroxidase: Gene expression and regulation. Enzyme Microb. Technol. 52, 1–12 (2013).
Akinfemi, A., Adu, O. & Doherty, F. Conversion of sorghum stover into animal feed with white-rot fungi: Pleurotus ostreatus and Pleurotus pulmonarius. J. Biotechnol. 9, 875 (2010).
Wirasnita, R. & Hadibarata, T. Potential of the white-rot fungus Pleurotus pulmonarius F043 for degradation and transformation of fluoranthene. Pedosphere 26, 49–54 (2016).
Fernández-Fueyo, E. et al. Ligninolytic peroxidase genes in the oyster mushroom genome: Heterologous expression, molecular structure, catalytic and stability properties, and lignin-degrading ability. Biotechnol. Biofuels 7, 1–23 (2014).
Yamaguchi, M., Nakamura, M., Takano, M. & Sekiya, A. Quantification of the mycelial mass of the white-rot fungus Pleurotus pulmonarius by real-time PCR. Bull FFPRI 8, 133–141 (2009).
Phengnuam, T. & Suntornsuk, W. Detoxification and anti-nutrients reduction of Jatropha curcas seed cake by Bacillus fermentation. J. Biosci. Bioeng. 115, 168–172 (2013).
Nakao, M., Hasegawa, G., Yasuhara, T. & Ishihara, Y. Degradation of Jatropha curcas phorbol esters derived from Jatropha oil cake and their tumor-promoting activity. Ecotoxicol. Environ. Saf. 114, 357–364 (2015).
Castellani, O., Ribeiro, L. & Fernandes, J. Differentiation of Trypanosoma cruzi in culture. J. Protozool. Res. 14, 447–451 (1967).
Makkar, H., Aderibigbe, A. & Becker, K. Comparative evaluation of non-toxic and toxic varieties of Jatropha curcas for chemical composition, digestibility, protein degradability and toxic factors. Food Chem. 62, 207–215 (1998).
Acknowledgements
This work was funded by Brazilian funding bodies FAPDF (Grant 193.001195/2016), CNPq/Embrapa (Grant 404786/2013-8) and CAPES (Grants Capes-Embrapa 15/2014 and CAPES Finance Code 001). RNGM was supported by a fellowship from CNPq (Grant 308165/2021-7).
Author information
Authors and Affiliations
Contributions
R.N.G.M., F.C.A.F., G.S.C.A. and F.G.S. conceived and designed the study. T.G.G. conducted the bioassays and RT-qPCR analysis. F.C.A.F. conducted RT-qPCR analysis. T.G.G., F.C.A.F., G.S.C.A., F.G.S. and R.N.G.M. analysed the data, provided intellectual input, wrote and revised the manuscript. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gomes, T.G., de Assis Fonseca, F.C., Alves, G.S.C. et al. Development of reference genes for RT-qPCR analysis of gene expression in Pleurotus pulmonarius for biotechnological applications. Sci Rep 13, 12296 (2023). https://doi.org/10.1038/s41598-023-39115-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-39115-4
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
