
Abstract
Soil degradation resulting from deforestation contributes to a dramatic decline in soil quality whose restoration must go through reforestation with pioneer species. We investigated the effects of cypress and black locust, pioneer but exotic species, on soil chemical properties and microbial and enzymatic activities of two marginal soils. The sampling sites were Lama Giulia and Locone lake in the Murge plateau of the Apulia Region, Italy. The soils at Lama Giulia presented a silty loam texture, while at Locone Lake site were sandy, and most likely due to the different texture, the former exhibited higher organic C, N, P and micronutrients contents than Locone Lake under black locust reforestation, despite the latter was reforested earlier. In addition, the higher microbial entropy and turnover of Locone Lake’s soils suggested a less conservative soil state than Lama Giulia’s soils. The effects of black locust reforestation at Lama Giulia on almost all soil parameters considered did not differ from those of the corresponding pasture, confirming the more conservative soil state in that site and suggesting that the time of reforestation was not enough to get differences between the reforested and not reforested soil. The soils reforested with cypress showed the significantly highest SOC, N, dissolved organic C and microbial biomass C content. In addition, it presented also the numerically largest dehydrogenase, phosphatase and β-glucosidase activities, soluble carbohydrates, and phenolic compounds content. These results may be ascribed to the longer litter deposition occurred in cypress soils.
Similar content being viewed by others


Introduction
Land degradation is caused by natural and anthropogenic processes threatening the capacity of natural resources to carry out their ecological functions1. Soil degradation is related to a decline in the multiple ecosystem services relying on it, as a result of adverse changes in its biological, chemical, physical and hydrological properties2, affecting the soil capacity to perform a range of ecological functions3.
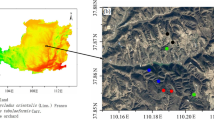
In the north-western sectors of Murge plateau, an inland part of Apulia region, southern Italy (Fig. 1), land degradation is mainly attributed to the intensive deforestation that started since the roman time due to the large amount of timber removed, which was necessary for the construction of warships4. The original floristic composition of the north-western sectors of Murge plateau was dominated by Quercus dalechampii Ten. and Quercus virgiliana Ten., with the contribution of species belonging to the Quercetalia pubescenti—petraeae order. The Stipo Bromoidis—Quercetum Dalechampii phytocoenosis was completed by Stipa bromoides L., Crataegus laevigata (Poir.) DC., Lonicera etrusca Santi, Carex hallerana Asso, and Iris collina N. Terracc. The ancient forest is today reduced to remnant strips because of the intense exploitation for coppicing, and sheep and goat pasturing5. The pastures not protected by trees were affected by intense erosion and “evolved” into degraded pastures, shallow soils or even outcropping rocks6.

Typical landscape of Murge plateau.
In order to solve such critical situation, several reforestations were realized over time utilizing pioneer species, such as Pinus halepensis L., Pinus pinea L. and Cupressus spp., and the nitrogen fixing black locust (Robinia pseudoacacia L.), due to their ability to grow up on shallow and eroded soils. Although the black locust does not enjoy a good reputation as a forest tree because of its invasiveness7,8, it has been utilized extensively for reforestation. These pioneer trees prevent erosion by covering the soil, reduce the impact of rainstorms erosion and produce, over time, litter and root exudates that increase the depth and the organic matter content of soils and can modify the biogeochemical cycles of N and C. These circumstances restore the soil ecological functions to reestablish native forestry species, such as several oaks, walnut, linden, etc., that need higher level of soil quality in comparison to pioneer forestry species.
Since the reforestations have not been conducted in a homogeneous way, in the Murge plateau one can find reforestations that differ in age of plantation and pioneer trees used. Therefore, we selected two sites that diverge for the aforementioned characteristics and investigated their effects on chemical, microbial, and enzymatic soil properties involved in the biogeochemical nutrients cycling, such as microbial biomass, organic matter, soil enzyme activities and soil microbial entropy that are some of the most well-known soil quality indicators9,10.
Materials and methods
Site description and samples collection
The Murgia plateau decreases its elevation from about 680 m a.s.l down to the Adriatic Sea, is characterized by an annual rainfall of about 650 mm11, concentrated mainly in fall and winter, and the rainwater reaches the sea through several incised karst valleys, locally called “lame”, that act as ephemeral streams only during the rainstorms12. From 2004 this area belongs to the “Alta Murgia National Park”.
The first sampling site was Lama Giulia (40°57′12.21′′ N; 16°20′14.75′′ E), belonging to the municipality of Gravina in Puglia, characterized by Calcic Haploxeralf fine loamy mixed thermic soils13. It was reforested with Cupressus arizonica Greene (CYG) during the years 1976–1980 (about 40 years of reforestation at the time of soil sampling), and with Robinia pseudoacacia L. (ROG), whose plantation dates to the end of 1990’s—beginning of 2000’s (about 17 years of reforestation at the time of soil sampling)14. Cupressus arizonica Greene is a conifer belonging to the family of Cupressaceae native to Arizona and New Mexico. It is also used as an ornamental tree, for timber and as a windbreak15. Robinia pseudoacacia L. is a broad-leaf tree belonging to the family of Leguminosae native to the North America. It is a promising fast-growing species for biomass production in short rotation coppice system16. It is also used as ornamental tree and windbreak, and for honey production8. The pasture near CYG and ROG, consisting in degraded but not reforested fields, was considered as control (COG) to check the effects of reforestation. It is mainly composed of shrubs and herbs belonging to four vegetation classes such as Artemisietea Vulgaris, Lygeo-Stipetea, Festuco-Brometea, and Stellarietea Mediae17. This site is close to an abandoned enclosure built with dry stone walls, commonly used in the past for sheltering sheeps and goats.
The other sampling site was located near the Locone Lake (41°04′52.88′′ N; 16°00′54.45′′ E), belonging to the municipality of Minervino Murge, characterized by Typic Calcixerept sandy mixed thermic soils13. It was mainly reforested with Robinia pseudoacacia L. (ROM) in 1985 (about 30 years of reforestation at the time of soil sampling)18. As per Lama Giulia, the pasture near ROM was considered as control (COM). It is currently characterized by arid mediterranean meadows comprising Brachipodium resutum (Pers.) P.Beauv., Brachipodium ramosum Roem. & Schult., Trachynia distachya (L.) P.Beauv., Bromus madritensis L., and Lagurus ovatus L.19. This site shows no signs of farming, even in the past, so grazing is presumably due only to wild herbivores.
Each site was sampled in spring 2017 using an auger, removing the litter and the herbaceous layer, and coring the soil until the abundant root apparatus depth (about 20–25 cm). In details, three composite samples (each made of five subsamples) were cored from CYG, ROG, COG, ROM, and COM, air dried, gently crushed and passed through a 2-mm sieve.
Physical and chemical soil analyses
Soil samples were analyzed according to standard methods20. The soil organic carbon (SOC) content was measured by the Walkley–Black method, and the SOC enrichment was calculated as the difference between the reforested SOC content and the SOC content of the corresponding control soil (pasture). The total nitrogen (TKN) was determined by the Kjeldahl method and the TKN enrichment was calculated as per SOC enrichment. The available phosphorus (Pava) was determined by UV–Vis spectrophotometry according to Olsen method. The exchangeable Ca and K were extracted with a BaCl2 and triethanolamine solution buffered at pH 8.2 and determined using the inductively coupled plasma optical emission spectrometer (ICP-OES) iCAP 6300 (Thermo Electron). The available fractions of Fe, Zn, Cu, Mn and Mo were extracted with diethylenetriaminepentaacetic acid (DTPA) and quantified using the ICP-OES.
Particle-size distribution was determined by the pipette method, and the soil texture was identified using the USDA soil textural classification system13.
Dissolved organic carbon (DOC), soluble carbohydrates and phenolic compounds of soil were analyzed on water extracts obtained at a soil to water ratio 1:10. DOC was determined with a Shimadzu TOC-VCSH analyzer (Kyoto, Japan). Soluble carbohydrates were determined spectrophotometrically by the anthrone method21 and expressed as glucose C equivalents (mg C kg-1). Phenolic compounds were determined by the method of Lowry et al.22 based on the Folin-Ciocalteu reagent and expressed as p-coumaric acid equivalents (mg kg-1).
Soil biochemical analyses
Dehydrogenase activity was measured as the reduction of 2-p-iodophenyl-3-p-nitrophenyl-5-phenyltetrazolium chloride (INT) to iodonitrophenyl-formazan (INTF)23 and expressed as micrograms of INTF produced per soil gram (dry mass) per h (μg INTF g−1 h−1). Microbial biomass C (MBC) content was determined by fumigation of the soil sample with ethanol-free CHCl3 and extraction with 0.5 M K2SO4, according to the method of Vance et al.24 modified by Gregorich et al.25, measured in a Shimadzu TOC-VCSH analyzer (Kyoto, Japan) and expressed in mg C per soil kg.
Alkaline phosphatase activity was determined following the method of Nannipieri et al.26, using p-nitrophenyl phosphate disodium (pNPP, 0.115 M) as substrate for this enzyme. This assay is based on the release and detection of p-nitrophenol (pNP) measured spectrophotometrically. β-glucosidase activity was analyzed according to the method of Tabatabai27, using p-nitrophenyl β-d-glucopyranoside (pNG, 0.05 M) as substrate for this enzyme. Phosphatase and β-glucosidase activities were expressed in micromoles of p-nitrophenol (pNP) produced per soil gram (dry weigh) per h (μmol pNP g−1 h−1).
Urease and protease activities were determined in 0.1 M phosphate buffer at pH 7; 1 M urea and 0.03 M N-α-benzoyl-argininamide (BAA) were used as substrates, respectively. Two mL of buffer and 0.5 mL of substrate were added to 0.5 g of the soil sample, which was incubated at 30 °C (urease) or 39 °C (protease) for 90 min. Both enzymatic activities were determined by the amount of NH4+ released26, expressed in units of micromoles of ammonium-N produced per soil gram (dry mass) per h (μmol NH4+-N g−1 h−1).
Statistical analysis
All experimental data were tested against the normal distribution of variables (Shapiro–Wilk’s test) and the homogeneity of variance (Bartlett’s test) using the RStudio software. The variables normally distributed and with homogeneity of variances were subjected to one-way ANOVA and HSD test, while results not normally distributed were subjected to Levene’s test to check the homogeneity of variance: data with heterogeneous variance were subjected to the nonparametric Friedman test.
The principal component analysis (PCA) was performed to compare the two sites that had different chemical, microbial, enzymatic and botanical characteristics. It was achieved using XLSTAT software and shown as biplots of scores (sites) and loadings (variables).
The regression analyses were performed using the RStudio software and the goodness of fit was evaluated through R2.
Results
All soils from Lama Giulia showed a silty loam texture, with an average composition of sand, silt and clay of 22.23, 54.04 and 23.73%, respectively, whereas all soils from Locone Lake were sandy (84% sand, 9.1% silt and 6.8% clay, on average).
Table 1 shows the average values of the main chemical parameters of the soil samples from Lama Giulia. Noteworthy, the SOC content was significantly (p = 0.0003) the highest in the soil samples of cypress with an average value of 94.3 g kg−1, followed by Robinia pseudoacacia L. (58.0 g kg−1) and pasture soils (45.1 g kg−1). The SOC enrichment of each reforested soil was linearly correlated with the age of each reforestation (Fig. 2). TKN resembled the SOC trend, with a significantly (p = 0.005) higher value recorded in soil samples from cypress (7.0 g kg−1) than black locust and pasture, that showed 4.9 and 3.9 g TKN kg−1, respectively. As per SOC, the linear correlation between the age of each reforestation and the TKN enrichment of each reforested soil showed that the differences of TKN content between the tree species was highly dependent by the time of planting (Fig. 2). In contrast, the Pava was the significantly highest (p = 0.002) in the soil samples from black locust (390.33 mg kg−1), while in cypress and pasture soils was 14.1 and 32.66 mg kg−1, respectively. Finally, the black locust soil showed numerically and significantly higher content of exchangeable Zn and Cu (p = 0.03), respectively, than the other two soils.

Linear regression between the age of each reforestation and the soil organic carbon enrichment. ROG black locust at Lama Giulia), ROM black locust at Locone Lake, CYG cypress at Lama Giulia.
The main chemical characteristics of soils from Locone Lake are reported in Table 2. The reforested soil had significantly higher SOC (p = 0.012) and TKN (p = 0.006) content than pasture, while all other parameters did not show any significant differences.
Table 3 reports the microbial and enzymatic properties of Lama Giulia soils. The cypress soil showed significantly higher MBC (p = 0.0001) and DOC (p = 0.003) content than black locust and control soil. In addition, also the MBC/SOC ratio was significantly higher (p = 0.011) in the cypress soil than the other two ones. Finally, the protease activity was significantly higher (p = 0.02) in reforested soils than pasture.
The microbial and enzymatic characteristics of Locone Lake soils are reported in Table 4. The soil from black locust showed significantly higher phosphatase (p = 0.016), protease (p = 0.048) and urease (p = 0.03) activities than pasture, together to significantly higher DOC (p = 0.0074) and phenolic compounds (p = 0.043) content. In contrast, the MBC/SOC ratio was significantly lower (p = 0.0061) in reforested soil than pasture.
The PCA analysis isolated five principal components, but only the first two were considered because they explained 74.2% of the variability (Fig. 3). Along the first principal component F1 there was a clear segregation between the points belonging to Lama Giulia (on the right) and those belonging to Locone Lake (on the left) (Fig. 3a). This result explained 50.15% of variability and it was mainly supported by SOC (load 0.940), TKN (load 0.952), soluble carbohydrates (load 0.813), phenolic compounds (load 0.783) and DOC (load 0.771) content, pH value (load −0.928 and −0.909 for pHH2O and pHKCl, respectively) and phosphatase activity (load 0.886) and, to a lesser extent, by Ca, Fe, Zn, Cu and Mn content (Fig. 3b; Table 5). Along the second principal component F2, the black locus from Lama Giulia (ROG) was clearly segregated from the black locust of the other site (ROM) and from the cypress of the same site (CYG) (Fig. 3a). This result explained 20.06% of variability and it was mainly supported by K content (load −0.740), and by glucosidase (load 0.722), urease (load 0.701) and dehydrogenase (load 0.768) activities (Table 5).

Principal component analysis shown as biplots of scores. (a) Sites, (b) variables loadings. ROM black locust at Locone Lake, COM degraded but not reforested soils at Locone Lake, CYG cypress at Lama Giulia, ROG black locust at Lama Giulia, COG degrade but not reforested soils at Lama Giulia.
Discussion
The two sites were segregated for their SOC content, with Lama Giulia soils showing a general higher SOC content than Locone Lake site. The larger SOC content of the cypress soil than black locust soils could be the result of about forty years of litter deposition occurred in the former soil with respect to the about thirty and seventeen years of black locust litter deposition at Locone Lake and Lama Giulia sites, respectively. In addition, the quality of the plant residues could have also played a role in C sequestration since the litter contribution is related to the type of plant28 and, in particular, black locust litter has a relatively higher input of aliphatic carbon, while the conifer litter composition is more aromatic and shows a slower decomposition29.
The Lama Giulia site differed from Locone Lake one for the TKN content too. Although a higher TKN content was expected in black locust soils because of the biological N fixation (BNF), the concentration of TKN was the highest in the cypress soil and mainly related to the litter deposition over time, since 90–95% of soil N is organic. Certainly, cypress also establishes relationships with beneficial microorganisms of the rhizosphere, while the BNF can be influenced by environmental conditions. Wei et al.30 reported that R. pseudoacacia L. forms nodules with many different bacterial species and this can explain the environmental success of black locust, but Schulze et al.31 and Zahran32 reported that the symbiotic association and the BNF are sensitive to soil water availability and drought stress. For example, Veste and Kriebitzsch33 demonstrated that, under long-term drought stress, the leaf N content of black locust reduced due to the negative influence of extreme drought on BNF. Even though Robinia pseudoacacia L. can grow in semi-arid environments, such as the Apulia region ones, it originated from climatic regions with annual rainfall of 1020 to 1830 mm34. Therefore, since in Apulia the balance between rainfall and evapotranspiration is frequently negative during late spring and summer, the drought may have affected the BNF.
The pH values of soils from Lama Giulia were lower than those from Locone Lake and inversely proportional to their SOC content since the soil organic matter offers a large source of acidic protons in the form of carboxyl groups that can determine a decrease of pH values35.
According to the PCA analysis, Locone Lake site showed a general lower phosphatase activity than Lama Giulia and, within each site, no significant difference was found in Lama Giulia (Table 3), while the black locust of Locone Lake showed significantly larger phosphatase activity than the corresponding pasture (Table 4). Since the phosphatase activity is mostly promoted by higher content of soil organic P and total N, and lower pH, the results are in accordance with the trend of those variables between the two sites and within each site. However, Pava has been demonstrated that did not correlate with phosphatase activity36. The highest content of Pava recorded in ROG almost certainly was related to the grazing cattle present in that area which enriched the site with their excreta, since black locust is also a food source for livestock and wild herbivores. This result is apparently confirmed also by the significantly higher content of Zn and Cu in the black locust soil within the Lama Giulia site (ROG), since those elements are often related to the animal droppings.
The two sites were segregated also by the soluble carbohydrates, phenolic compounds and DOC that represent labile fractions of the soil organic matter directly affected by the aboveground and belowground litter inputs. In general, the significantly largest DOC content and the numerically higher carbohydrates and phenolic compounds contents of the cypress soil suggest that the quality of the cypress litter has a better effect than black locust and pastures litters to maintain a pool of labile organic C that provide a source of energy for soil microbial activities. In fact, the chemical composition of the litter species such as lignin, polyphenols, and the C-to-N and lignin-to-N ratios control the litter decomposition and the environmental conditions for soil microbiota37. Indeed, the majority of enzymatic activities are correlated with SOC and the easily degradable organic compounds in soil38,39, which stimulate the growth of soil microbial community.
The PCA analysis indicated, even to a lesser extent, that CYG and ROM soils shared a slightly higher dehydrogenase activity than ROG. Dehydrogenase activity is an index of the global soil microbial metabolic activity and health status of the soil ecosystem40. Dehydrogenase is exclusively an intracellular enzyme that is only present in living microbial cells and therefore is a well-founded marker for monitoring changes in soil metabolism41. As the ecosystems develop with time, there is a reduction of metabolic activity and tendency to reach energy equilibrium in the system42,43, therefore the results pointed out that all soils showed different environmental conditions that modulated the changes in the overall soil metabolism. This result was confirmed also by the activities of the extracellular enzymes β-glucosidase and urease, involved in organic matter turnover and in soil N metabolism, respectively44. In fact, they showed lower activities in ROG soil than the other tree soils.
The protease showed a significantly tree-related stimulation of its activity over the pasture soils within each site (Tables 3 and 4), in response to the higher TKN content of such soils than corresponding pastures that stimulated the synthesis of this enzyme by soil microbial community45.
The MBC is a reliable indicator of changes in soil quality reflecting the long-term effects of soil-vegetation system and it was positively correlated to the SOC contents (R2 = 0.84). Even if the MBC did not segregate the two sites, within the Lama Giulia one it reached the significantly highest value in the cypress soil because of the longer inputs of litter that increased the food resources for microbes and the amount of soil microorganisms. In addition, De Marco et al.46 found that black locust soils had low MBC and respiratory response because of the presence of inhibitory substances, such as 4-hydroxyacetophenon, in black locust litter extracts, that negatively affects the abundance and activity of the soil microbial community and the microbial-derived organic matter formation.
Soil microbial entropy, defined as the ratio of soil MBC to SOC, is a sensitive indicator of the soil quality, assessing the SOC quality and its turnover rate47. It reflects the percentage of the active SOC in the soil ecosystem and the changes of soil environment48. MBC generally account for 1–4% of the total SOC49. Comparing the two pastures, the higher soil microbial entropy of COM with respect to COG was possibly due to the sandy texture of the former soil, that ensured a higher SOC turnover. For the same reason, ROM had higher MBC/SOC ratio than ROG. Within Lama Giulia site, CYG showed the slightly but significantly highest soil microbial entropy because of the amount of litter deposition and, to a lesser extent, the quality of the litter.
Conclusions
The effects of Robinia pseudocacia L. on Locone Lake were remarkable, with an increase of SOC and TKN content in comparison to the corresponding pasture. Comparing the two black locust plantations, the different soil texture between the two sites determined higher values of such parameters in Lama Giulia than Locone Lake, despite the latter was reforested earlier than the former site. In fact, the Locone Lake’s sandy soils showed a higher microbial entropy and turnover, especially in the not reforested soil, suggesting a less conservative soil state than Lama Giulia’s silty loam soil.
Within the Lama Giulia site, CYG soil showed the highest values of some chemical and microbial parameters due to the longest time of reforestation. In contrast, almost all soil parameters studied were not statistically different between the ROG and the corresponding pasture, suggesting that about 17 years of reforestation with black locust apparently are not sufficient to establish general better soil quality.
Data availability
The data presented in this study are available within the article.
References
Sacristán, D., Peñarroya, B., Recatalá, L. Increasing the Knowledge on the Management of Cu-Contaminated Agricultural Soils by Cropping Tomato (Solanum Lycopersicum L.). Land Degrad. Dev. 26, 587–595 (2015).
FAO. Land Degradation Assessment in Drylands. Manual for Local Level Assessment of Land Degradation and Sustainable Land Management. Part 1: Planning and Methodological Approach, Analysis and Reporting. https://www.fao.org/3/i6362e/i6362e.pdf (Food and Agriculture Organization of the United Nations, 2011).
Vlachodimos, K., Papatheodorou, E. M., Diamantopoulos, J. & Monokrousos, N. Assessment of Robinia pseudoacacia cultivations as a restoration strategy for reclaimed mine spoil heaps. Environ Monit. Assess. 185, 6921–6932 (2013).
Misano, G. & Di Pietro, R. Habitat 9250 “Quercus trojana woods” in Italy. Fitosociologia 44, 235–238 (2007).
Biondi, E. et al. A contribution towards the knowledge of semideciduous and evergreen woods of Apulia (south-eastern Italy). Fitosociologia 41(1), 3–28 (2004).
Brunetti, G. et al. Remediation of a heavy metals contaminated soil using mycorrhized and non-mycorrhized Helichrysum italicum (Roth) Don. Land Degrad. Dev. 29, 91–104 (2017).
Poblador, S. et al. The influence of the invasive alien nitrogen-fixing Robinia pseudoacacia L. on soil nitrogen availability in a mixed Mediterranean riparian forest. Eur. J. For. Res. 138, 1083–1093 (2019).
Vítková, M., Müllerová, J., Sádlo, J., Pergl, J. & Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 384, 287–302 (2017).
Doran, J.W., Parkin, T.B. Quantitative indicators of soil quality: a minimum data set. in Methods for Assessing Soil Quality (eds. Doran, J.W., Jones, A.J.). 25–37 (Soil Science Society of America, 1996).
Gil-Sotres, F., Trasar-Cepeda, C., Leirós, M. C. & Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 37, 877–887 (2005).
Andriani, G. F. & Walsh, N. An example of the effects of anthropogenic changes on natural environment in the Apulian karst (southern Italy). Environ. Geol. 58, 313–325 (2009).
Bisantino, T., Pizzo, V., Polemio, M. & Gentile, F. Analysis of the flooding event of October 22–23, 2005 in a small basin in the province of Bari (Southern Italy). J. Agric. Eng. 531, 197–204 (2016).
Soil Survey Staff. Keys to Soil Taxonomy 12th edn. (USDA-Natural Resources Conservation Service, 2014).
Tartarino, P. Inventario dei Boschi Spontanei e dei Rimboschimenti delle Provincie BAT e Bari e Stima del Loro Volume Legnoso e della sua Frazione Prelevabile nel Prossimo Ventennio. (Rapporto Tecnico Scientifico, 2011).
Ismail, A. et al. Chemical composition and biological activities of Tunisian Cupressus arizonica Greene essential oils. Chem. Biodivers. 11, 150–160 (2014).
Navarro, A. et al. Feasibility of SRC Species for growing in Mediterranean conditions. Bioenerg. Res. 9, 208–223 (2015).
Perrino, E. V., Brunetti, G. & Farrag, K. Plant communities in multi-metal contaminated soils: A case study in the National Park of Alta Murgia (Apulia Region-Southern Italy). Int. J. Phytoremediat. 16, 871–888 (2014).
VV AA Perizia Studi per il Riequilibrio Socio-Economico dell'area Interessata dall'invaso sul Torrente Locone. Consorzio Di Bonifica Apulo Lucano (1986).
Lavarra, P. et al. Il Sistema Carta della Natura della Regione Puglia. (ISPRA, Serie Rapporti 204, 2014).
Sparks, D. L. et al. Method of Soil Analysis: Part 3 (American Society of Agronomy Inc, 1996).
Brink, R. H. Jr., Dubach, P. & Lynch, D. L. Measurement of carbohydrates in soil hydrolyzates with anthrone. Soil Sci. 89, 157–166 (1960).
Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 193, 265–275 (1951).
García, C., Hernandez, T. & Costa, F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plant Anal. 28, 123–134 (1997).
Vance, E. D., Brookes, P. C. & Jenkinson, D. S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 19, 703–707 (1987).
Gregorich, E. G., Wen, G., Voroney, R. P. & Kachanoski, R. G. Calibration of a rapid direct chloroform extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 22, 1009–1011 (1990).
Nannipieri, P., Ceccanti, B., Cervelli, S. & Matarese, E. Extraction of phosphatase, urease, protease, organic carbon and nitrogen from soil. Soil Sci. Soc. Am. J. 44, 1011–1016 (1980).
Tabatabai, M.A. (1994) Soil enzymes. in Methods of Soil Analysis. Part 2. Microbiological and Biochemical Properties (eds. Weaver, R.W. et al.). 775–833 (Soil Science Society of America, Inc., 1996)
Traversa, A., Said-Pullicino, D., D’Orazio, V., Gigliotti, G., & Senesi, N. Properties of humic acids in Mediterranean forest soils (Southern Italy): Influence of different plant covering. Eur. J. For. Res. 130, 1045–1054 (2011)
De Marco, A. et al. Decomposition of black locust and black pine leaf litter in two coeval forest stands on Mount Vesuvius and dynamics of organic components assessed through proximate analysis and NMR spectroscopy. Soil Biol. Biochem. 51, 1–15 (2012).
Wei, G. et al. Invasive Robinia pseudoacacia in China is nodulated by Mesorhizobium and Sinorhizobium species that share similar nodulation genes with native American symbionts. FEMS Microbiol. Ecol. 68, 320–328 (2009).
Schulze, E. D., Gebauer, G., Ziegler, H. & Lange, O. L. Estimates of nitrogen fixation by trees on an aridity gradient in Namibia. Oecologia 88, 451–455 (1991).
Zahran, H. H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 63, 968–989 (1999).
Veste, M. & Kriebitzsch, W. U. Influence of drought stress on photosynthesis, transpiration, and growth of juvenile black locust (Robinia pseudoacacia L.). Forstarchiv 84, 35–42 (2013).
Nicolescu, V. N. et al. Ecology, growth and management of black locust (Robinia pseudoacacia L.), a non-native species integrated into European forests. J. For. Res. 31, 1081–1101 (2020).
Sposito, G. The Chemistry of Soil (Oxford University Press, 2008).
Margalef, O. et al. Global patterns of phosphatase activity in natural soils. Sci. Rep. 7, 1337. https://doi.org/10.1038/s41598-017-01418-8 (2017).
Prescott, C. E. & Grayston, S. J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 309, 19–27 (2013).
Frankenberger, W. T. & Dick, W. A. Relationships between enzyme, activities and microbial growth and activity indices in soil. Soil Sci. Soc. Am. J. 47, 945–951 (1983).
Frankenberger, W.T., Tabatabai, M.A. Amidase activity in soils III. Stability and distribution. Soil Sci. Soc. Am. J. 45, 333–338 (1981).
Nannipieri, P., Trasar-Cepeda, C. & Dick, R. P. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soils 54, 11–19 (2018).
Pascual, J. A., Garcia, C., Hernandez, T., Moreno, J. L. & Ros, M. Soil microbial activity as a biomarker of degradation and remediation processes. Soil Biol. Biochem. 32, 1877–1883 (2000).
García-Gil, J. C., Plaza, C., Solker-Rovira, P. & Polo, A. Long-term effects of municipal solid waste compost application on soil enzyme activities and microbial biomass. Soil Biol. Biochem. 32, 1907–1913 (2000).
Insam, H. & Domsch, K. H. Relationship between soil organic carbon and microbial biomass on chronosequences of reclamation sites. Microb. Ecol. 15, 177–188 (1988).
Acosta-Martinez, V. & Tabatabai, M. Enzyme activities in a limed agricultural soil. Biol. Fertil. Soils 31, 85–91 (2000).
Uselman, S. M., Qualls, R. G. & Thomas, R. B. A test of a potential short cut in the nitrogen cycle: the role of exudation of symbiotically fixed nitrogen from the roots of a N-fixing tree and the effects of increased atmospheric CO2 and temperature. Plant Soil 210, 21–32 (1999).
De Marco, A., Esposito, F., Berg, B., Zarrelli, A. & Virzo De Santo, A. Litter inhibitory effects on soil microbial biomass activity, and catabolic diversity in two paired stands of Robinia pseudoacacia L. and Pinus nigra Arn. Forest 9, 766. https://doi.org/10.3390/f9120766 (2018).
Haghverdi, K. & Kooch, Y. Effects of diversity of tree species on nutrient cycling and soil-related processes. CATENA 178, 335–344 (2019).
Anderson, H. T. Microbial eco-physiological indicators to assess soil quality. Agric. Ecosyst. Environ. 98, 285–293 (2003).
Jenkinson, D.S., Ladd, J.N. Microbial biomass in soil: Measurement and turnover. in Soil Biochemistry (eds. Paul, E.A., Ladd, J.N.). 415–471 (Marcel Dekker Inc., 1981)
Acknowledgements
Authors acknowledge Dr. Francesco Ceglie, CIHEAM Bari, for his support for PCA analysis.
Author information
Authors and Affiliations
Contributions
C.Co. and C.Ca. contributed to the conception of the work. C.Ca., J.C.G.G., F.D.M., A.T. and C.Co. contributed to the analyses. All authors contributed to the interpretation of data. C.Co., C.Ca., J.C.G.G. and A.T. have drafted the work. All authors have approved the submitted version. All authors have agreed both to be personally accountable for the author’s own contributions and to ensure that questions related to the accuracy or integrity of any part of the work, even ones in which the author was not personally involved, have appropriately investigated, resolved, and the resolution documented in the literature.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cacace, C., García-Gil, J.C., Cocozza, C. et al. Effects of different pioneer and exotic species on the changes of degraded soils. Sci Rep 12, 18548 (2022). https://doi.org/10.1038/s41598-022-23265-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-23265-y
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
