
Abstract
Juveniles of three cyprinids with various diets and habitat preferences were collected from the Szamos River (Hungary) during a period of pollution in November 2013: the herbivorous, benthic nase (Chondrostoma nasus), the benthivorous, benthic barbel (Barbus barbus), and the omnivorous, pelagic chub (Squalius cephalus). Our study aimed to assess the accumulation of these elements across species with varying diets and habitat preferences, as well as their potential role in biomonitoring efforts. The Ca, K, Mg, Na, Cd, Cr, Cu, Fe, Mn, Pb, Sr, and Zn concentration was analyzed in muscle, gills, and liver using MP-AES. The muscle and gill concentrations of Cr, Cu, Fe, and Zn increased with trophic level. At the same time, several differences were found among the trace element patterns related to habitat preferences. The trace elements, including Cd, Pb, and Zn, which exceeded threshold concentrations in the water, exhibited higher accumulations mainly in the muscle and gills of the pelagic chub. Furthermore, the elevated concentrations of trace elements in sediments (Cr, Cu, Mn) demonstrated higher accumulation in the benthic nase and barbel. Our findings show habitat preference as a key factor in juvenile bioindicator capability, advocating for the simultaneous use of pelagic and benthic juveniles to assess water and sediment pollution status.
Similar content being viewed by others


Introduction
Trace element pollution seriously threatens the biosphere, especially aquatic ecosystems1,2. These pollutants and their environmental importance have been widely known since the Minamata disease, one of the first documented human diseases caused by trace element pollution3,4. In aquatic ecosystems, trace elements have two primary sources: the natural geological background and anthropogenic activities, such as agriculture, industry, and mining5,6. Pollution of the aquatic environment with trace elements can be especially harmful due to their toxicity, non-biodegradable features, and long persistence7. When these elements enter aquatic ecosystems, they can accumulate in the water, sediments, and aquatic organisms8.
The chemical analysis of water and sediments is a traditional tool for monitoring trace element pollution9. Due to their bioaccumulation and biomagnification susceptibility, the trace element concentrations in aquatic organisms could be relatively higher than those in water and sediments10,11,12. Among these organisms, fish have attracted considerable interest in monitoring aquatic contamination 13,14,15. Fish can provide a longer-term, complex sight of the pollution status of their environment because all environmental factors (such as trace element contamination) integrate into their organs during their lifespan8,16. Furthermore, fish is one of the most important food sources for humans; thus, monitoring their trace elements patterns is essential to ensure the safety of fish consumption17,18,19.
Fish can take up trace elements by several routes, e.g., through the skin and gills or the digestive system20,21. Therefore, trace element accumulation in fish depends on several factors, such as habitat preference and the diet of different species22. For example, several studies suggest that species with benthic habitat preferences and piscivorous or benthivorous diets may accumulate higher concentrations of trace elements22,23,24, while others found that pelagic and omnivorous fish accumulate more trace elements25. At the same time, the trace element patterns are different in the different tissues of a given species because of their various physiological features20,26. In general, the trace element concentrations are higher in the liver and gills compared to those in muscle tissue; hence, they have been widely used as bioindicator organs for trace element pollution of aquatic ecosystems in monitoring and risk assessment programs27,28. As a secondary target of bioaccumulation of trace elements, the muscle generally contains lower concentrations of trace elements than the liver or gills29. However, the muscle is the main part of the fish consumed by humans; therefore, monitoring its trace element pattern is equally important19,30.
Traditionally, most of the field studies focused on the trace element patterns of adult fish6,31,32. However, the bioaccumulation of trace elements is influenced not only by habitat preference and diet but also by age and size18,29,33,34,35,36,37,38. During the last decade, a considerable amount of research highlighted the bioindicator capability of juveniles to assess the trace element pollution of aquatic ecosystems18,29,39. The trace element concentrations of juveniles can be higher than those of adult fish due to various mechanisms, e.g., increased sensitivity, inadequately developed detoxification system, and higher relative metabolic rate18,29,36,38. In addition, juveniles integrate the environmental effects of the current year only; therefore, as a result of the phenomena mentioned above, they can be used as bioindicators of the recent trace element pollution18. However, there is a scarcity of data regarding the functional traits, such as diet and habitat preference, that influence the bioindicator characteristics of juveniles inhabiting a polluted aquatic ecosystem.
Nase [Chondrostoma nasus (Linnaeus, 1758)] is a widespread cyprinid species in Western and Central Europe, with a benthic habitat preference40. Its primary habitat is moderate to fast-flowing large to medium-sized rivers with rock or gravel bottoms40. The larvae feed mainly on zooplankton, while the juveniles and adults feed almost exclusively on benthic algae cleaned up from rock or stone surfaces using their specific lower lip with thick cornified sheath35,40.
Barbel [Barbus barbus (Linnaeus, 1758)] is one of the most frequent cyprinid species in Europe, living in medium-sized and large rivers with moderate or fast-flowing water and gravel bottom40. It has a benthic habitat preference and a benthivorous diet, feeding chiefly on benthic invertebrates and detritus40.
Chub [Squalius cephalus (Linnaeus, 1758) previously Leuciscus cephalus] is also one of the most common and widespread cyprinid species in Europe40. Chub is most abundant in small rivers and large streams, even in slow-flowing lowland rivers and mountain streams40. It has a pelagic habitat preference and omnivorous diet, feeding on various aquatic organisms40.
At the beginning of their developmental stages, larvae and early juveniles of these species primarily rely on zooplankton as their main food source, and they typically inhabit shallow shoreline habitats40. However, after they advance through early development, significant ontogenetic shifts in both feeding behavior and habitat preference become apparent41,42,43,44, i.e., older juveniles gradually shift towards consuming species-specific prey items, adopting similar habitat preferences to adults several months after hatching45,46,47.These three cyprinid species are popular game fish and are commercially exploited in several European countries22,40,48. Furthermore, they are considered good bioindicators of the health of aquatic ecosystems22,35,49,50,51,52,53,54,55,56,57,58,59,60.
The Szamos River is one of the most polluted rivers in Central Europe, mainly due to the intensive mining activities in Romania11,61. In 2000, 100,000 m3 of cyanide and trace element-rich liquid waste was released into the Water System of the Szamos River62. The pollution of the Szamos River and its recipient, the Tisza River, has been named the worst environmental disaster in Europe since the Chernobyl nuclear leak in 198662,63. After this ecological catastrophe, the trace element concentrations rapidly decreased in the Szamos River61,64,65. Nonetheless, a significant level of pollution with Cd, Cr, Cu, Mn, Pb, Sr, and Zn was observed in 2013 based on earlier studies11,18,66. Subsequently, according to water chemistry data from the National Environmental Information System of Hungary, there was no notable pollution in the Szamos River.
This study aimed to investigate the trace element accumulation pattern in different tissues of nase, barbel, and chub juveniles from the polluted aquatic ecosystem of the Szamos River. Previous research highlighted the potential of juveniles as effective bioindicators of recent trace element pollution18,29,39, prompting their selection for this investigation. However, further exploration is needed to elucidate how habitat preference and diet may influence the bioindicator capabilities of juveniles. Different trace element patterns of juveniles of nase, barbel, and chub were hypothesized. It was also predicted that the trace element concentrations would be the highest in juveniles with benthic habitat preference and higher trophic levels due to higher exposure from their microhabitat and diet. Hence, diverse patterns of trace element accumulation in juveniles of three cyprinids with various habitat preferences and diets were analyzed to test these hypotheses. At the same time, the potential risks of fish consumption on human health were also evaluated. Moreover, the bioindicator potential of juveniles was evaluated to investigate the suitability of the studied species as indicators of trace element pollution specifically during the polluted period in 2013, which was selected as the focal year due to its heightened contamination levels.
Results
Biological features of fish
A total of 15 cyprinids were investigated. The mean standard length and standard deviation of nase, barbel, and chub were 72.3 ± 3.8 mm, 52.1 ± 2.4 mm, and 60.6 ± 5.1 mm, respectively. The mean body weight and standard deviation of nase, barbel, and chub were 5.57 ± 1.10 g, 2.56 ± 0.32 g, and 4.06 ± 1.23 g, respectively. According to the length and weight data, the studied fish were categorized as the 0 + age class (juveniles)67,68. The trophic level of nase, barbel, and chub were 2.0, 3.1, and 2.7, respectively69.
Characterization of the study area
The descriptive statistics of the concentrations of trace elements in water, obtained from the database of the National Environmental Information System of Hungary (OKIR in Hungarian), are presented in Table 1. The mean concentrations of Cu, Pb, and Zn were above the criterion chronic concentrations (CCCs) for the freshwater of the National Recommended Water Quality Criteria prescribed by the USEPA70 (Table 1). The annual mean concentration of Cd remained below the threshold; however, the monthly mean concentrations surpassed the CCCs multiple times in 2013 (Table 1).
The descriptive statistics of the concentrations of trace elements in the sediments obtained from previous studies11,66 are presented in Table 1. The mean concentration of Cr represented the sediments as moderately polluted. In contrast, the mean concentrations of Cu, Mn, and Zn classified the sediment status as heavily polluted, according to the toxicity classification of sediments71.
Macroelements
In this study, the macroelements (Ca, K, Mg, Na) and trace elements (Cd, Cr, Cu, Fe, Mn, Pb, Sr, Zn) were separately evaluated due to their various physiological and bioaccumulation features. The mean concentrations of macroelements in muscle, gills, and liver are summarized in Table 2.
The Kruskal–Wallis test revealed significant differences among the three species concerning the concentrations of K, Mg, and Na (p < 0.05) in muscle, Ca and Mg (p < 0.05) in gills, and K, Mg, and Na (p < 0.05) in the liver (Table 2).
In muscle, the lowest concentrations of K and Mg for barbel, while the highest concentrations of Na for chub were recorded (p < 0.05). In gills, the highest concentration of Ca for chub and lowest concentrations of Mg for barbel were measured (p < 0.05). In the liver, the concentrations of K, Mg, and Na were significantly higher in nase compared to chub (p < 0.05) (Table 2).
There was a significant (p < 0.05) decrease in the concentrations of some macroelements with trophic levels (Table 3). K concentrations in muscle, Na concentrations in the liver, and Mg concentrations in muscle and gills were negatively correlated with trophic levels (Table 3).
Essential and non-essential trace elements
The mean concentrations of the trace elements in muscle, gills, and liver are presented in Table 2. The concentrations of Cd in the muscle, gills, and liver of nase and the muscle and gills of barbel were below the detection limit. In general, the highest concentrations of Cd, Cr, and Pb were measured in the liver, while Fe, Mn, and Sr reached the highest concentrations in the gills. The concentrations of Cu and Zn were highest in the liver of nase and barbel, respectively. However, Zn concentrations were most elevated in the gills of chub, and Cu concentrations in the gills of nase were lower than those of chub (Table 2). The trace element concentrations were generally lowest in the muscle in all the studied fish species (Table 2).
The Kruskal–Wallis test detected significant differences among the species in the trace element concentrations of Cd, Cr, Cu, Fe, Mn, Sr, and Zn (p < 0.05) in the muscle, Cd, Cr, Cu, Mn, Sr, and Zn (p < 0.05) in the gills, and Cd, Cr, Cu, Fe, and Pb (p < 0.05), in the liver.
In the case of muscle, the highest concentrations of Cr and Cu were found in barbel, the highest concentrations of Sr in nase, while concentrations of Cd were highest in chub (Mann–Whitney U test, p < 0.05) (Table 2). The lowest concentrations of Mn were found in chub, and Zn concentrations were the lowest in nase (Mann–Whitney U test, p < 0.05) (Table 2). In the case of gills, the highest concentrations of Cd and Zn were measured in chub (Mann–Whitney U test, p < 0.05) (Table 2). The lowest concentrations of Cr were found in chub, those of Cu and Zn were measured in nase, while the concentrations of Sr were lowest in barbel (Mann–Whitney U test, p < 0.05) (Table 2). In the case of the liver, the highest concentrations of Cu were found in nase (Mann–Whitney U test, p < 0.05) (Table 2). The lowest concentrations of Cd were measured in nase, while Cr and Pb accumulated in the lowest concentrations in chub (Mann–Whitney U test, p < 0.05) (Table 2).
Furthermore, the concentrations of several trace elements showed a significant increase in terms of the trophic level of fish (p < 0.05) (Table 3). The concentrations of Cr, Cu, Fe, and Zn in muscle and concentrations of Cu and Zn in gills showed a significant (p < 0.05) positive correlation with the trophic level (Table 3). The concentrations of Sr in muscle and gills and those of Cu in the liver showed a significant (p < 0.05) decrease with trophic levels (Table 3).
The PCA showed clear separations among the different species based on trace element patterns in muscle (Fig. 1A) and the liver (Fig. 1C). In the case of muscle, the first component (PCA 1) contributed 58.85% of the total variance, while the second (PCA 2) contributed 20.75% of the total variance (Fig. 1A). In the case of liver, the first component contributed 44.92% of the total variance, the second (PCA 2) contributed 29.58% of the total variance (Fig. 1C). The trace element patterns in gills were also well separated among the different species. However, nase showed a minimal overlap with barbel and chub (Fig. 1B). In the case of gills, the first component (PCA 1) explained 53.72% of the total variance, while the second component (PCA 2) described the 20.61% of the total variance (Fig. 1B). The elements exhibiting significant correlations with the PCA1 and PCA2 axes, along with their respective Spearman's rho values, are indicated on Fig. 1.

The principal component analysis of trace element concentrations (mg kg−1, wet weight) in the muscle (A), gills (B), and liver (C) of the three cyprinid species from the Szamos River. The elements exhibiting significant correlations with the PCA1 and PCA2 axes and their respective Spearman’s rho values are indicated.
Bioconcentration factor and metal pollution index
The bioconcentration factors for water and sediments are summarized in Table 4. In general, the highest BCF values for water for Cd, Cr, Cu, and Pb were observed in the liver. At the same time, the highest BCF values for water for Fe and Mn were found in the gills (Mann–Whitney U test, p < 0.05).
In the case of muscle, the highest BCF values for water for Cd were found in chub, while those for Cr and Cu were measured in barbel. The lowest BCF values for water for Mn were in chub and those for Zn were found in nase (Mann–Whitney U test, p < 0.05). In gills, the highest BCF values for water for Cd and Zn were in chub, while those for Cr were in barbel. At the same time, the lowest BCF values for water for Cu and Zn were in nase (Mann–Whitney U test, p < 0.05). In the liver, the highest BCF values for water for Cu were in Nase, while the lowest for Cd were found in nase, and those for Cr were in chub (Mann–Whitney U test, p < 0.05) (Table 4A).
Similar tendencies were also observed in the BCF values for the sediments (Mann–Whitney U test, p < 0.05) (Table 4B). In addition, the BCF values for the sediments for Sr were highest generally in the gills (Mann–Whitney U test, p < 0.05).
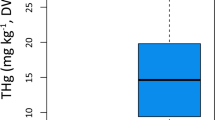
The metal pollution index (MPI) values of muscle and gills were highest in barbel, while the MPI of the liver was highest in nase (Mann–Whitney U test, p < 0.05) (Fig. 2.). The lowest MPI values were observed in chub in all examined organs (Mann–Whitney U test, p < 0.05) (Fig. 2.).

Metal pollution index (MPI) in the different organs of cyprinid juveniles from the Szamos River.
Human health implications
The mean concentrations of Cd and Pb in the chub muscle were over the EU’s limit: Cd (0.05 mg kg−1 ww) and Pb (0.30 mg kg−1 ww)72. Additionally, concentrations of the other analyzed trace elements in the muscle of the studied species were found to be below the Maximum Allowable Concentrations (MACs in mg kg−1 ww): Cd (0.05), Pb (0.30)72, Cr (1.0), Cu (30.0), Fe (43.0), Mn (1.0), and Zn (40.0)73.
Discussion
Both our study and earlier research have emphasized that the Szamos River experienced substantial trace element pollution in 2013, originating from industrial, mining, and agricultural activities11,18,66. The trace element pattern of sediments is a stable indicator of long-term pollution of rivers, while that of water reflects recent pollution74. The sediments of the Szamos River were moderately polluted with Cr and heavily polluted with Cu, Mn, and Zn, indicating that this aquatic ecosystem has been continuously polluted by these elements11,66. At the same time, the concentrations of Cu, Cd, Pb, and Zn in the river’s water exceeded the CCCs for the freshwater of EPA70 in 2013. According to the water chemistry data of the National Environmental Information System of Hungary (OKIR in Hungarian), the concentrations of these elements were approximately 2–10 times higher than those in previous years.
The distribution of macroelements (Ca, K, Mg, Na) in fish were found according to their biological functions. The highest concentrations of Ca and Mg were found in the gills because these elements play an essential role in forming and maintaining bony structures (i.e., arches and rakers in gills)75. Elevated concentrations of Na were measured in the liver of nase and barbel, while similarly high concentrations were detected in the gills of chub. This phenomenon can be attributed to the essential role of Na, an electrolyte, in liver functioning. Additionally, the gills serve as the primary site for Na uptake from water. 18,76. Similar results were observed in field studies25,60,77 and laboratory experiments with juveniles of common carp (Cyprinus carpio Linnaeus, 1758)13.
Among the investigated species, the herbivorous nase had the lowest trophic level, followed by the omnivorous chub, while the benthivorous barbel had the highest trophic level. The Cr, Cu, Fe, and Zn concentrations in muscle and Cu and Zn concentrations in gills increased with the trophic level. These results partially confirmed our first hypothesis that the concentration of trace elements would be highest in species with higher trophic levels. A central part of the positive correlation between trace element concentration and the trophic level was observed in the case of muscle, independently from their pollution status in water and sediment because the concentrations of Cu and Zn were over the limit of polluted levels in the water and sediment too70,71. At the same time, the Fe concentrations did not exceed these limits, and Cr was detected only in levels describing a moderately polluted status in the sediments70,71. According to these results, many trace elements in the muscle increased with trophic level, which indicated the significant influence of diet on the trace element patterns of the muscle of juveniles18,36,38. During the last decade, several authors also highlighted the importance of the muscle tissue in bioaccumulation and biomagnification of trace elements in the case of juveniles18,29,34,38,39.
In addition, some trace elements did not show a correlation between their concentrations and the trophic level, despite significant differences, especially in the gills and liver. Moreover, recent studies also revealed that habitat preference has a more substantial influence on the trace element pattern of juveniles than the trophic level itself33,78,79. This phenomenon may further support our results since the concentrations of the central part of trace elements did not differ between the benthic, herbivorous nase, and benthivorous barbel (except for Cu). In contrast, they differed between these benthic species and the pelagic, omnivorous chub. Previous studies conducted in Serbian water bodies, including the Danube River, Kačer River, Zaovine, and Medjuvršje reservoirs, also proved elevated Cu concentrations in the liver of nase22,35,60. Nase feeds exclusively on benthic algae, and according to published data, high amounts of Cu can accumulate in green algae and diatoms80,81. The significant negative correlation between Cu concentrations in the liver and trophic level among the investigated cyprinids can potentially be attributed to the specific diet of nase. This is because, among the species studied, nase exhibits the lowest trophic level.
The non-essential (or toxic) trace elements negatively affect health even at low concentrations, e.g., Cd and Pb82. Cd was detected in elevated concentrations in the tissues of pelagic chub, while its concentrations were below the detection limit in the gills of nase and barbel and the liver of nase. The Cd concentration of the water of the Szamos River was approximately ten times higher in 2013 than in previous years. It exceeded the CCCs during several months in 2013 based on the water chemistry data from the National Environmental Information System of Hungary (OKIR in Hungarian). The elevated concentration of Cd in water was reflected in the trace element pattern of pelagic chub, whereas its impact on benthic juveniles was less pronounced. Similar patterns were observed in multispecies investigations, wherein elevated Cd concentrations in the water primarily accumulated in pelagic species52,79. At the same time, Pb concentrations were highest in the liver of benthic juveniles. In general, habitat preference accounts for the trace element pattern of gills6,18. However, both Cd and Pb concentrations were elevated in the water, but the primary source of Pb accumulation in fish is the food consumed, while fish take up Cd directly from the water through their gills29,83. According to this phenomenon, toxic trace elements that primarily enter fish through their gills from water have a more significant effect on the trace element pattern of pelagic species than on that of benthic species6,24,52. These results confirmed our second hypothesis that habitat preference influences trace element patterns.
Several mechanisms make fish juveniles effective bioindicators of trace element pollution of aquatic ecosystems, e.g., relative higher growth and metabolic rate, relative food intake and quantity of respiratory water passing through the gills, and inadequately developed detoxification system18,29,38,78. The primary objective of this study was to examine the bioindicator potential of various juveniles and evaluate the influence of their diet and habitat preferences on it within a polluted aquatic ecosystem. Cu and Zn polluted both the water and the sediment. In this case, when the microhabitats of species were relatively equally contaminated, the diet significantly affected the bioaccumulation of these elements. However, in the case of pelagic chub, the Zn concentrations in the gills were higher than in the liver. The highest Zn concentrations can generally be found in the liver35,84,85. This revealed that relatively higher Zn concentrations in the gills of a pelagic species could reflect a recent Zn pollution15,86,87. Cr and Mn were in moderately and heavily polluted concentrations in the sediments, respectively, while their concentrations did not exceed the CCCs in the water. These elements bioaccumulated in higher concentrations in the benthic species (nase, barbel) than in the pelagic chub. Cd concentrations stayed over the CCCs of water for several months, while Cd did not accumulate in the sediments. As a result of this phenomenon, Cd was found at higher concentrations in tissues of the pelagic chub than in benthic nase and barbel.
These results highlighted that habitat preference influenced the bioindicator capability of juveniles more than the diet29,78. However, the diet and trophic level of nase, barbel, and chub differed significantly, but in larvae and early juvenile stages, they fed on the same prey40. Fish larvae onsets the exogenous feeding after the depletion of yolk reserves41. Most larvae and early juveniles feed exclusively on zooplankton, showing an ontogenic shift in feeding later45. Juveniles commence feeding on their species-specific prey items several months after hatching45. Therefore, in juveniles, due to the similar feeding habits during the early stages of the lifespan, the diet may not affect the trace element patterns as much as in the case of adults18,35,36. However, the investigated juveniles were caught from the same section of the Szamos River; different environmental trace element exposure impacted them. Because fish with benthic habitat preference frequently cause bioturbation, which can increase trace element release from the sediment88. Consequently, the trace element pattern of the microenvironment of a benthic and a pelagic fish is different, even in the same water body. Trace elements could also interact with each other during their uptake by fish through biological barriers, such as the gills or digestive system. Furthermore, habitat preference developed earlier than the final species-specific diet in the case of juveniles. Based on the results, we consider that the habitat preference of juveniles can seriously impact the bioaccumulation of trace elements. According to the phenomena mentioned above, the bioaccumulation patterns of trace elements in juveniles with various habitat preferences can be utilized as effective bioindicators of trace element pollution of aquatic ecosystems. Thus, the simultaneous use of pelagic and benthic juveniles can be very helpful also in applied monitoring programs for determining water and sediment pollution status.
In the case of muscle, the mean concentrations of Cd and Pb in the chub were over the prescribed MACs. Interestingly, the concentrations of these toxic elements were also over the CCCs in the water, while their concentrations were not in a polluted status in the sediments. This also followed our finding that the pelagic chub was more sensitive to recent water pollution than the benthic nase and barbel. Hungary has a size limit for both investigated cyprinids, and anglers can harvest only adults. Nevertheless, higher concentrations of Cd and Pb in the muscle of chub juveniles can be dangerous to the ecosystem (due to biomagnification in the food web) and human health (due to illegal poaching). Therefore, further detailed examinations of trace element patterns of adult fish from the Szamos River are required to assess human health risks.
Conclusions
This study provides new information on the ecotoxicology of water, sediments, and juveniles of nase, barbel, and chub from the Szamos River. Overall, the trace element concentrations observed in both water and sediments were notably elevated, with some exceeding threshold concentration values. This highlights the significant pollution burden of the Szamos River in 2013. The accumulated trace element patterns of the juveniles showed significant differentiation. The various habitat preferences and diets could explain these differences. However, only a few trace element concentrations showed correlations with the trophic levels, especially in the case of muscle. At the same time, several differences were found among the species related to habitat preference. The pelagic chub had a higher affinity for bioaccumulation of trace elements, which exceeded the threshold concentrations of water. In contrast, the benthic nase and barbel had a higher affinity for trace elements that exceeded the sediment threshold concentrations. These may reflect different exposure of the juveniles with various habitat preferences due to their different microhabitat and several mechanisms, e.g., bioturbation. As a result of recent trace element pollution of water, the concentrations of toxic Cd and Pb in the muscle tissue of pelagic chub exceeded the prescribed MACs by the EU. Therefore, it is advisable to monitor the health risks associated with the consumption of fish living in the Szamos River, particularly by sampling adult individuals. Nase, barbel, and chub have an important role in the food web, angling, and commercial fishing, in addition to their widespread status in the rivers of Europe. According to our results, future monitoring studies should simultaneously focus on juveniles or adults with various habitat preferences as adequate bioindicators of trace element pollution of water and sediments. However, further studies are needed to identify such patterns in other habitats.
Material and methods
Ethical approval
The Workplace Animal Experiments Committee of Debrecen University approved the experimental protocol and the end-points of the experiments. All methods were carried out following relevant national and international guidelines and regulations (permission number: HBH/01/00971-2/2013). The study complies with the Animal Research: Reporting of in Vivo Experiments (ARRIVE) guidelines.
Study area and sample collection
The Szamos River is one of the largest tributaries of the Tisza River. Its watershed collects water over a 15,882 km2 area of Romania and Hungary. Its basin is strongly affected by anthropogenic activities, such as mining and industrial emissions, and thus, it is continuously polluted by trace elements11,18. Fish were collected at the settlement, Csenger, near the Hungarian-Romanian border. The geocoordinates of the sampling site are N47.838303 and E22.693744.
For this study, a total of 15 juveniles of nase, barbel, and chub (5 individuals per fish species) were collected in November 2013 by electrofishing (Hans Grassl IG200/2b, PDC, 75–100 Hz, 350–650 V, max. 10 kW, Hans Grassl GmbH, Germany). The sample size for each group was designed similarly to the study by35. Additionally, the fisheries society authorized the collection of up to 5 specimens per species. The fish were transported in containers with aerated river water to the laboratory. The standard length (SL) and the total weight (W) of each specimen were measured to the nearest 0.1 mm and 0.01 g, respectively. After the measurements, the fish were sacrificed immediately by spinal severance and stored at -18 °C until sample processing.
Sample processing and element analysis
Muscle samples were dissected from the dorsolateral muscle of the left side. The second gill arch from the left side was collected as a gill sample. After the incision of the abdominal wall, the whole liver was sampled. During the dissection, sterile plastic tools were used to avoid any trace element contamination. The dissected samples were rinsed with double deionized water (Milli-Q), and their wet weight (WW) was measured into glass beakers using an analytical balance (Precisa 240A, Switzerland). The samples were dried overnight at 105 °C. The samples were afterward digested on an electric hot plate using 4.0 ml 65% (m/m) nitric acid (reagent grade, Merck, USA) and 1.0 ml 30% (m/m) hydrogen-peroxide (reagent grade, Merck, USA) in the same container at 80 °C for 4 h. The digested samples were then diluted with 1% (m/m) nitric acid (reagent grade, Merck, USA) and Milli-Q water up to a final volume of 10 ml89,90.
The Ca, K, Mg, Na, Cd, Cr, Cu, Fe, Mn, Pb, Sr, and Zn concentrations were measured by a microwave plasma-atomic emission spectrometer (MP-AES 4200, Agilent Technologies, USA) system. The values of the limits of detection (LOD) in μg l−1 were: Ca, 0.070; K, 0.270; Mg, 0.080; Na, 0.484; Cd, 1.863; Cr, 0.489; Cu, 0.414; Fe, 0.856; Mn 0.057; Pb, 1.235; Sr, 0.051; and Zn, 3.125, respectively. Each sample’s detection limit was verified based on the instrumental detection limit, sample mass, and volume to which it had been diluted. The average detection limits for each of the assessed elements were (mg kg−1 wet weight): Ca, < 0.001; K, 0.002; Mg, 0.001; Na, 0.004; Cd, 0.009; Cr, 0.004; Cu, 0.003; Fe, 0.006; Mn < 0.001; Pb, 0.009; Sr, < 0.001; and Zn, 0.023, respectively. The following wavelength lines of the MP-AES analysis were used: Ca 422.673 nm, K 766.491 nm, Mg 285.213 nm, Na 588.995 nm, Cd 228.802 nm, Cr 425.433 nm, Cu 324.754 nm, Fe 371.993 nm, Mn 403.076 nm, Pb 405.781 nm, Sr 407.771 nm, and Zn 481.053 nm.
We applied a five-point calibration procedure prepared from the multi-element standard solution (Merck ICP multi-element standard solution IV). An autosampler (Agilent SPS4, USA), a Meinhard-type nebulizer, and a double-pass spray chamber were used. The measurement used a certified reference material (ERM-BB422, fish muscle, LGC Standards, UK). The recoveries were within 10% of the certified values for the metals. The wavelengths and measuring parameters were chosen based on suggestions provided by the instrument’s software (MP Expert). The concentrations of all elements were expressed as mg kg−1 wet weight (ww).
Bioconcentration factors and metal pollution index
The bioconcentration factor (BCF) of a trace element is defined as the proportion of the concentration of that trace element in an organism (or in a specific tissue of the organism) to the concentration of the trace element in the water or the sediment91:
where Cfish means the concentration of the trace element in the whole body or tissue of the organism, expressed as mg kg−1 wet weight, Cwater is the concentration of the trace element in the water, described as mg l−1., and Csediment is the concentration of the trace element in the sediment, expressed as mg kg−1 dry weight.
The trace element concentration data for water were sourced from the database of the National Environmental Information System of Hungary (OKIR in Hungarian). For this study, we utilized the average concentrations derived from monthly measurements conducted in 2013. Specifically, data from April to November were used because of this period is important regarding the lifespan of the studied juveniles. The trace element concentrations of sediment from the Szamos River from 2013 were used based on earlier studies 11,66. References11,66 collected and analyzed the physical and chemical parameters from the same sites and the same period than in the present study. The water data were compared with criterion chronic concentrations (CCCs) for the freshwater of the National Recommended Water Quality Criteria70. The trace element concentrations in the sediments were evaluated according to the toxicity classification of sediments by Baudo and Muntau (2020).
Metal pollution index (MPI) was assessed to compare the total content of trace elements (Cd, Cr, Cu, Fe, Mn, Pb, Sr, Zn), excluding the macroelements for the different tissues of the different species. The MPI formula is the following92,93:
where Cn is the mean concentration of trace element n in the analyzed tissue (mg kg−1 wet weight).
Statistical analysis
The data were presented as mean values accompanied by standard deviation. The statistical analysis was performed with IBM SPSS Statistics for Windows (Version 20.0)94 and Past 4.095. Before analyses, the normal distribution and homogeneity of variances were tested with Shapiro–Wilk and Levene’s tests, respectively. The non-parametric Kruskal–Wallis test was used to evaluate the differences in the trace element concentrations and BCF values of trace elements in the tissues of nase, barbel, and chub. As a post hoc test, a Mann–Whitney U test was used to explore the significant differences. Spearman’s rank correlation test was used to study the relationship between element concentrations and trophic levels of fish in order to explore possible biomagnification features of elements in juveniles96,97. The trophic levels were obtained from the database of FishBase69. A principal component analysis (PCA) was used to assess the differentiation of species based on the concentration of trace elements (Cd, Cr, Cu, Fe, Mn, Pb, Sr, Zn) of the studied tissues, except for the macroelements. Spearman’s rank correlation test was conducted between the concentration of trace elements and the component scores of the first two PC axes to explore variables significantly affecting the differentiation of species.
To assess the risk for human consumption of nase, barbel, and chub from the Szamos River, trace element concentrations of muscle (filet) were compared with the maximum acceptable concentrations (MACs) established by the European Union72 and the Food and Agriculture Organization of the United Nations73.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
References
Nriagu, J. O. Global metal pollution poisoning the biosphere?. Environment 32, 6 (1990).
Verma, P. & Ratan, J. K. Assessment of the negative effects of various inorganic water pollutants on the biosphere—an overview. Inorg. Pollut. Water https://doi.org/10.1016/b978-0-12-818965-8.00005-6 (2020).
Harada, M. Minamata disease: Methylmercury poisoning in Japan caused by environmental pollution. Crit. Rev. Toxicol. 25, 1–24 (1995).
Semionov, A. Minamata disease—Review. World J. Neurosci. 8, 178–184 (2018).
Cajaraville, M. P. et al. The use of biomarkers to assess the impact of pollution in coastal environments of the Iberian Peninsula: A practical approach. Sci. Total Environ. 247, 295–311 (2000).
Subotić, S. et al. Heavy metal and trace element bioaccumulation in target tissues of four edible fish species from the Danube River (Serbia). Ecotoxicol. Environ. Saf. 98, 196–202 (2013).
Sary, A. A. & Mohammadi, M. Lead bioaccumulation and toxicity in tissues of economically fish species from river and marine water. Bull. Environ. Contam. Toxicol. 89, 82–85 (2012).
Jordanova, M. et al. Accumulation of heavy metals in some organs in Barbel and Chub from Crn Drim river in the Republic of Macedonia. Bull. Environ. Contam. Toxicol. 101, 392–397 (2018).
Jovanovic, B., Mihaljev, Ž, Maletin, S. & Palić, D. Assessment of heavy metal load in chub liver (Cyprinidae—Leuciscus cephalus) from the Nišava River (Serbia). Biol. Nyssana 2(1), 51–58 (2011).
Pandiyan, J. et al. An assessment of level of heavy metals pollution in the water, sediment and aquatic organisms: A perspective of tackling environmental threats for food security. Saudi J. Biol. Sci. 28, 1218–1225 (2021).
Simon, E. et al. Assessment of contamination based on trace element concentrations in Gomphus flavipes (Odonata: Insect) larvae of the Upper Tisza Region. Ecotoxicol. Environ. Saf. 136, 55–61 (2017).
Simon, E. et al. Environmental-friendly contamination assessment of habitats based on the trace element content of dragonfly exuviae. Water (Switzerland) 11, 2200 (2019).
Harangi, S. et al. Accumulation of metals in Juvenile carp (Cyprinus carpio) exposed to sublethal levels of iron and manganese: Survival, body weight and tissue. Biol. Trace Elem. Res. 177, 187–195 (2017).
Jia, Y., Wang, L., Qu, Z. & Yang, Z. Distribution, contamination and accumulation of heavy metals in water, sediments, and freshwater shellfish from Liuyang River, Southern China. Environ. Sci. Pollut. Res. 25, 7012–7020 (2018).
Subotić, S. et al. Distribution and accumulation of elements (As, Cu, Fe, Hg, Mn, and Zn) in tissues of fish species from different trophic levels in the Danube River at the confluence with the Sava River (Serbia). Environ. Sci. Pollut. Res. 20, 5309–5317 (2013).
Plessl, C. et al. Mercury, silver, selenium and other trace elements in three cyprinid fish species from the Vaal Dam, South Africa, including implications for fish consumers. Sci. Total Environ. 659, 1158–1167 (2019).
Anandkumar, A., Nagarajan, R., Prabakaran, K., Bing, C. H. & Rajaram, R. Human health risk assessment and bioaccumulation of trace metals in fish species collected from the Miri coast, Sarawak, Borneo. Mar. Pollut. Bull. 133, 655–663 (2018).
Nyeste, K. et al. Age and diet-specific trace element accumulation patterns in different tissues of chub (Squalius cephalus): Juveniles are useful bioindicators of recent pollution. Ecol. Indic. 101, 1–10 (2019).
Traina, A. et al. Heavy metals concentrations in some commercially key species from Sicilian coasts (Mediterranean Sea): Potential human health risk estimation. Ecotoxicol. Environ. Saf. 168, 466–478 (2019).
Yancheva, V., Velcheva, I., Stoyanova, S. & Georgieva, E. Fish in ecotoxicological studies. Ecol. Balk. 7, 149–169 (2015).
Yilmaz, F., Özdemir, N., Demirak, A. & Tuna, A. L. Heavy metal levels in two fish species Leuciscus cephalus and Lepomis gibbosus. Food Chem. 100, 830–835 (2007).
Subotić, S. et al. Metal accumulation in muscle and liver of the common nase (Chondrostoma nasus) and vimba bream (Vimba vimba) from the Danube River, Serbia: Bioindicative aspects. Bull. Environ. Contam. Toxicol. 103, 261–266 (2019).
Colombo, J. C. et al. Detritivorous fish contamination in the Rio de la Plata estuary: A critical accumulation pathway in the cycle of anthropogenic compounds. Can. J. Fish. Aquat. Sci. 57, 1139–1150 (2000).
Jiang, Z. et al. Metal concentrations and risk assessment in water, sediment and economic fish species with various habitat preferences and trophic guilds from Lake Caizi, Southeast China. Ecotoxicol. Environ. Saf. 157, 1–8 (2018).
Nikolić, D. et al. Accumulation of 25 elements in gills, liver, gonads, and muscle of European chub (Squalius cephalus), Cactus roach (Rutilus virgo), and pikeperch (Sander lucioperca) from Zlatar reservoir (Serbia). Environ. Sci. Pollut. Res. 29, 50271–50280 (2022).
Jovičić, K. et al. Mapping differential elemental accumulation in fish tissues: Assessment of metal and trace element concentrations in wels catfish (Silurus glanis) from the Danube River by ICP-MS. Environ. Sci. Pollut. Res. 22, 3820–3827 (2015).
Bachouche, S., Houma, F., Gomiero, A. & Rabah, B. Distribution and environmental risk assessment of heavy metal in surface sediments and red mullet (Mullus barbatus) from Algiers and BouIsmail Bay (Algeria). Environ. Model. Assess. 22, 473–490 (2017).
Dikanović, V., Skorić, S. & Gačić, Z. Concentrations of metals and trace elements in different tissues of nine fish species from the Meduvršje Reservoir (West Morava River Basin, Serbia). Arch. Biol. Sci. 68, 811–819 (2016).
Jia, Y., Wang, L., Qu, Z., Wang, C. & Yang, Z. Effects on heavy metal accumulation in freshwater fishes: Species, tissues, and sizes. Environ. Sci. Pollut. Res. 24, 9379–9386 (2017).
Karadede, H., Oymak, S. A. & Ünlü, E. Heavy metals in mullet, Liza abu, and catfish, Silurus triostegus, from the Atatürk Dam Lake (Euphrates). Turkey. Environ. Int. 30, 183–188 (2004).
Hermenean, A. et al. Histopatological alterations and oxidative stress in liver and kidney of Leuciscus cephalus following exposure to heavy metals in the Tur River, North Western Romania. Ecotoxicol. Environ. Saf. 119, 198–205 (2015).
Köhler, H. R. et al. Monitoring pollution in river Mureş Romania, part III: Biochemical effect markers in fish and integrative reflection. Environ. Monit. Assess. 127, 47–54 (2007).
Jiang, X. et al. Assessment of heavy metal accumulation in freshwater fish of Dongting Lake, China: Effects of feeding habits, habitat preferences and body size. J. Environ. Sci. (China) 112, 355–365 (2021).
Ndimele, P. E., Pedro, M. O., Agboola, J. I., Chukwuka, K. S. & Ekwu, A. O. Heavy metal accumulation in organs of Oreochromis niloticus (Linnaeus, 1758) from industrial effluent-polluted aquatic ecosystem in Lagos, Nigeria. Environ. Monit. Assess. 189, 1–15 (2017).
Djikanović, V., Skorić, S., Jarić, I. & Lenhardt, M. Age-specific metal and accumulation patterns in different tissues of nase (Chodrostoma nasus) from the Medjuvršje Reservoir. Sci. Total Environ. 566–567, 185–190 (2016).
Farkas, A., Salánki, J. & Specziár, A. Age- and size-specific patterns of heavy metals in the organs of freshwater fish Abramis brama L. populating a low-contaminated site. Water Res. 37, 959–964 (2003).
Has-Schön, E. et al. Distribution and age-related bioaccumulation of lead (Pb), mercury (Hg), cadmium (Cd), and arsenic (As) in tissues of common carp (Cyprinus carpio) and European catfish (Silurus glanis) from the Buško Blato reservoir (Bosnia and Herzegovina). Chemosphere 135, 289–296 (2015).
Nikolić, D., Skorić, S., Janković, S., Hegediš, A. & Djikanović, V. Age-specific accumulation of toxic metal(loid)s in northern pike (Esox lucius) juveniles. Environ. Monit. Assess. 193, 229 (2021).
Merciai, R., Guasch, H., Kumar, A., Sabater, S. & García-Berthou, E. Trace metal concentration and fish size: Variation among fish species in a Mediterranean river. Ecotoxicol. Environ. Saf. 107, 154–161 (2014).
Kottelat, M. & Freyhof, J. Handbook of European Freshwater Fishes. Kottelat, Cornol, Switzerland and Freyhof, Berlin (2007).
Heming, T. A. & Buddington, R. K. 6 Yolk absorption in embryonic and larval fishes. Fish Physiol. https://doi.org/10.1016/S1546-5098(08)60203-4 (1988).
Reckendorfer, W. et al. Diet shifts in 0+ nase, Chondrostoma nasus: Size-specific differences and the effect of food availability. River Syst. 12, 425–440 (2001).
Granado-Lorencio, C. & Garcia-Novo, F. Feeding habits of the fish community in a eutrophic reservoir in Spain. Ekol. Pol. 34, 95–110 (1987).
Mann, R. H. K. Observations on the age, growth, reproduction and food of the chub Squalius cephalus (L.) in the River Stour. Dorset. J. Fish Biol. 8, 265–288 (1976).
Yúfera, M. & Darias, M. J. The onset of exogenous feeding in marine fish larvae. Aquaculture 268, 53–63 (2007).
Baras, E. & Nindaba, J. Seasonal and diel utilisation of inshore microhabitats by larvae and juveniles of Leuciscus cephalus and Leuciscus leuciscus. In When do Fishes Become Juveniles? (eds Copp, Gordon H. et al.) 183–197 (Springer Netherlands, 1998). https://doi.org/10.1007/978-94-017-3678-7_13.
Hofer, K. & Kirchhofer, A. Drift, habitat choice and growth of the nase (Chondrostoma nasus, Cyprinidae) during early life stages. In Conservation of Endangered Freshwater Fish in Europe (eds Kirchhofer, A. & Hefti, D.) 269–278 (Birkhäuser Basel, 1996). https://doi.org/10.1007/978-3-0348-9014-4_26.
Lyach, R. Harvest rates of Rheophilic fish Vimba vimba, Chondrostoma nasus, and Barbus barbus have a strong relationship with restocking rates and harvest rates of their predator Silurus glanis in lowland mesotrophic rivers in central Europe. Sustainability 13, 11379 (2021).
Rašković, B. et al. Use of histopathology and elemental accumulation in different organs of two benthophagous fish species as indicators of river pollution. Environ. Toxicol. 30, 1153–1161 (2015).
Morina, A. et al. Common barbel (Barbus barbus) as a bioindicator of surface river sediment pollution with Cu and Zn in three rivers of the Danube River Basin in Serbia. Environ. Sci. Pollut. Res. 23, 6723–6734 (2016).
Sunjog, K. et al. Heavy metal accumulation and the genotoxicity in barbel (Barbus barbus) as indicators of the Danube river pollution. Sci. World J. https://doi.org/10.1100/2012/351074 (2012).
Milošković, A. et al. Spatial monitoring of heavy metals in the inland waters of Serbia: A multispecies approach based on commercial fish. Environ. Sci. Pollut. Res. 23, 9918–9933 (2016).
Demirak, A., Yilmaz, F., Levent Tuna, A. & Ozdemir, N. Heavy metals in water, sediment and tissues of Leuciscus cephalus from a stream in southwestern Turkey. Chemosphere 63, 1451–1458 (2006).
Zuliani, T. et al. Potentially toxic elements in muscle tissue of different fish species from the Sava River and risk assessment for consumers. Sci. Total Environ. 650, 958–969 (2019).
Randak, T. et al. Effects of pollution on chub in the River Elbe, Czech Republic. Ecotoxicol. Environ. Saf. 72, 737–746 (2009).
Winter, M. J. et al. Tissue levels and biomarkers of organic contaminants in feral and caged chub (Leuciscus cephalus) from rivers in the West Midlands, UK. Aquat. Toxicol. 73, 394–405 (2005).
Sunjog, K. et al. Assessment of status of three water bodies in Serbia based on tissue metal and metalloid concentration (ICP-OES) and genotoxicity (comet assay). Environ. Pollut. 213, 600–607 (2016).
Rašković, B. et al. Effects of mine tailing and mixed contamination on metals, trace elements accumulation and histopathology of the chub (Squalius cephalus) tissues: Evidence from three differently contaminated sites in Serbia. Ecotoxicol. Environ. Saf. 153, 238–247 (2018).
Sunjog, K. et al. Seasonal variation in metal concentration in various tissues of the European chub (Squalius cephalus L.). Environ. Sci. Pollut. Res. 26, 9232–9243 (2019).
Nikolić, D., Subotić, S. & Skorić, S. The common nase (Chondrostoma nasus) as an indicator of aquatic pollution and human health risk assessment associated with its consumption. Environ. Sci. Pollut. Res. Int. 31, 1050–1063 (2024).
Óvári, M. et al. Total reflection X-ray fluorescence spectrometric determination of element inlets from mining activities at the upper Tisza catchment area, Hungary. Spectrochim. Acta - Part B At. Spectrosc. 59, 1173–1181 (2004).
Bridges, O. & Bridges, J. Taking risks: Cyanide and heavy metal spill in Baia Mare, Romania. Environ. Prot. Bull. 72, 16–19 (2001).
Soldán, P., Pavonič, M., Bouček, J. & Kokeš, J. Baia mare accident—Brief ecotoxicological report of Czech experts. Ecotoxicol. Environ. Saf. 49, 255–261 (2001).
Mages, M., Woelfl, S., Óvári, M., Tümpling, W. V. & Encina, F. The use of a portable total reflection X-ray fluorescence spectrometer for trace element determination in freshwater microcrustaceans (Daphnia). Spectrochim. Acta - Part B At. Spectrosc. 59, 1265–1272 (2004).
Woelfl, S., Mages, M., Óvári, M. & Geller, W. Determination of heavy metals in macrozoobenthos from the rivers Tisza and Szamos by total reflection X-ray fluorescence spectrometry. Spectrochim. Acta - Part B At. Spectrosc. 61, 1153–1157 (2006).
Málnás, K. et al. Nehézfém analitikai vizsgálatok a Felső-Tisza és a Szamos folyó hazai szakaszán [Toxic element analysis on the Upper-Tisza and the River Szamos] [in Hungarian]. Hidrológiai Közlöny 5–6, 62–64 (2014).
Epler, P., Nowak, M. & Popek, W. Growth rate of the chub (Squalius cephalus) and the nase (Chondrostoma nasus) from Raba, Dunajec, and Poprad River. AACL Bioflux 2, 1–8 (2009).
Prokeš, M. et al. Growth of barbel, Barbus barbus, in the River Jihlava following major habitat alteration and estimated by two methods. Folia Zool. 55, 86 (2006).
Froese, R. & Pauly, D. Fishbase. World Wide Web electronic publication. Version (01/2023). https://www.fishbase.de (2023).
EPA. National recommended water quality criteria - aquatic life criteria table. https://www.epa.gov/wqc/national-recommended-water-quality-criteria-aquatic-life-criteria-table#table. (2018).
Baudo, R. & Muntau, H. Lesser known in-place pollutants and diffuse source problems. In Sediments (eds Baudo, R. et al.) 1–14 (CRC Press, 2020). https://doi.org/10.1201/9780367810894-1.
European Commission. Commission regulation (EC) no 629/2008. Off. J. Eur. Union (2008).
FAO. Compilation of legal limits for hazardous substances in fish and fishery product. Library (Lond). (1983).
Balogh, Z. et al. Effects of anthropogenic activities on the elemental concentration in surface sediment of oxbows. Water. Air. Soil Pollut. 227, 1–8 (2016).
Playle, R. C. Modelling metal interactions at fish gills. Sci. Total Environ. 219, 147–163 (1998).
Maetz, J. Transport of ions and water across the epithelium of fish gills. In Ciba Foundation Symposium 38 ‐ Lung Liquids (eds Porter, R. & O’Connor, M.) 133–159 (Wiley, 1976). https://doi.org/10.1002/9780470720202.ch9.
Subotić, S. et al. Seasonal variations of macro-, micro-, and toxic elements in tissues of vimba bream (Vimba vimba) from the Danube River near Belgrade. Serbia. Environ. Sci. Pollut. Res. 28, 63087–63101 (2021).
Llamazares Vegh, S. et al. Bioaccumulation of 10 trace elements in juvenile fishes of the Lower Paraná River, Argentina: Implications associated with essential fish growing habitat. Environ. Sci. Pollut. Res. 28, 365–378 (2021).
Llamazares Vegh, S., Biolé, F., Bavio, M., Tripodi, P. & Volpedo, A. V. Distribution and accumulation of trace elements in organs of juvenile fishes from a freshwater system (Paraná River, South America). Biol. Trace Elem. Res. 200, 2416–2431 (2022).
González-Dávila, M., Santana-Casiano, J. M. & Laglera, L. M. Copper adsorption in diatom cultures. Mar. Chem. 70, 161–170 (2000).
Novák, Z. et al. Effects of metal quantity and quality to the removal of zinc and copper by two common green microalgae (Chlorophyceae) species. Phycol. Res. 68, 227–235 (2020).
Singh, R., Gautam, N., Mishra, A. & Gupta, R. Heavy metals and living systems: An overview. Indian J. Pharmacol. 43, 246–253. https://doi.org/10.4103/0253-7613.81505 (2011).
Subotić, S. et al. Concentrations of 18 Elements in Muscle, Liver, Gills, and Gonads of Sichel (Pelecus cultratus), Ruffe (Gymnocephalus cernua), and European Perch (Perca fluviatilis) in the Danube River near Belgrade (Serbia). Water. Air. Soil Pollut. 226, 1–11 (2015).
Machala, M. et al. Determination and multivariate statistical analysis of biochemical responses to environmental contaminants in feral freshwater fish Leuciscus cephalus L.. Environ. Toxicol. Chem. 20, 1141 (2001).
Podrug, M. & Raspor, B. Seasonal variation of the metal (Zn, Fe, Mn) and metallothionein concentrations in the liver cytosol of the European chub (Squalius cephalus L.). Environ. Monit. Assess. 157, 1–10 (2009).
Harkabusová, V., Čelechovská, O., Lavičková, A. & Svobodová, Z. Monitoring of risk metals in chub (Leuciscus cephalus L.) from the Svitava and Svratka rivers in the urban area of Brno, Czech Republic. Acta Vet. Brno 81, 69–73 (2012).
Nikolić, D. et al. The European Chub (Squalius cephalus) as an indicator of reservoirs pollution and human health risk assessment associated with its consumption. Environ. Pollut. 310, 119871 (2022).
Adámek, Z. & Maršálek, B. Bioturbation of sediments by benthic macroinvertebrates and fish and its implication for pond ecosystems: A review. Aquac. Int. 21, 1–17 (2013).
Braun, M., Simon, E., Fábián, I. & Tóthmérész, B. The effects of ethylene glycol and ethanol on the body mass and elemental composition of insects collected with pitfall traps. Chemosphere 77, 1447–1452 (2009).
Braun, M., Simon, E., Fábián, I. & Tóthmérész, B. Elemental analysis of pitfall-trapped insect samples: Effects of ethylene glycol grades. Entomol. Exp. Appl. 143, 89–94 (2012).
Ivanciuc, T., Ivanciuc, O. & Klein, D. J. Modeling the bioconcentration factors and bioaccumulation factors of polychlorinated biphenyls with posetic quantitative super-structure/activity relationships (QSSAR). Mol. Divers. 10, 133–145 (2006).
Ju, Y. R., Chen, C. W., Chen, C. F., Chuang, X. Y. & Dong, C. D. Assessment of heavy metals in aquaculture fishes collected from southwest coast of Taiwan and human consumption risk. Int. Biodeterior. Biodegrad. 124, 314–325 (2017).
Usero, J., González-Regalado, E. & Gracia, I. Trace metals in the bivalve molluscs Ruditapes decussatus and Ruditapes philippinarum from the Atlantic Coast of Southern Spain. Environ. Int. 23, 291–298 (1997).
IBM Corp. Released. IBM SPSS Statistics for Windows, Version 20.0. 2011 (2011).
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4, 9 (2001).
Vetsis, E., Kalantzi, I., Pergantis, S. A., Kokokiris, L. & Karakassis, I. Metals in tissues of marine fish from the Thermaikos Gulf, Eastern Mediterranean Sea: Detection of changes with trophic level. Mar. Pollut. Bull. 173, 113024 (2021).
Annabi, A., Bardelli, R., Vizzini, S. & Mancinelli, G. Baseline assessment of heavy metals content and trophic position of the invasive blue swimming crab Portunus segnis (Forskål, 1775) in the Gulf of Gabès (Tunisia). Mar. Pollut. Bull. 136, 454–463 (2018).
Acknowledgements
Project no. TKP2021-NKTA-32 was implemented with the support provided from the National Research, Development and Innovation Fund of Hungary, financed under the TKP2021-NKTA funding scheme. The research presented in the article was carried out within the framework of the Széchenyi Plan Plus program with the support of the RRF 2.3.1-21-2022-00008 and GINOP_PLUSZ-2.1.1-21-2022-00245 projects. This work was supported by the European Union-NextGenerationEU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, project № BG-RRP-2.004-0001-C01. László Antal was supported by the János Bolyai Research Scholarship of the Hungarian Academy of Sciences. Researchers were supported by the ÚNKP-23-3-II (Dóra Somogyi) and ÚNKP-23-5 (László Antal) New National Excellence Program of the Ministry for Culture and Innovation from the source of the National Research, Development and Innovation Fund. Dóra Somogyi. was supported by the PhD Excellence Scholarship from the Count István Tisza Foundation for the University of Debrecen. István Czeglédi was supported by the OTKA PD 138296 grant (National Research, Development and Innovation Office – NKFIH). The elemental analysis was carried out in the Agilent Atomic Spectroscopy Partner Laboratory at the Department of Inorganic and Analytical Chemistry, University of Debrecen. Novo-Lab Ltd provided the MP-AES 4100 instrument. We want to express our thanks to Csilla Noémi Tóth, Judit Vári, and Patrik Dobrocsi for assistance in the laboratory and the Rákóczi Fisheries Cooperative (Rákóczi Halászati Szövetkezet) for the permission to collect fish.
Funding
Open access funding provided by University of Debrecen.
Author information
Authors and Affiliations
Contributions
K.N.: Conceptualization, Methodology, Formal analysis, Writing—Original Draft, Investigation. N.Z.: Formal analysis, Writing—Original Draft. I.E.U.: Formal analysis, Writing—Review and Editing. D.S.: Investigation, Writing—Review and Editing. L.N.: Investigation, Writing—Review and Editing. I.C.: Formal analysis, Writing—Review and Editing. S.H.: Conceptualization, Methodology, Validation. E.B.: Conceptualization, Methodology, Validation. E.S.: Conceptualization, Methodology, Supervision, Writing—Review and Editing. S.A.N.: Supervision, Formal analysis, Funding acquisition. I.V.: Formal analysis, Validation, Funding acquisition. V.Y.: Formal analysis, Writing—Review and Editing, Funding acquisition. L.A.: Conceptualization, Methodology, Resources, Writing—Review and Editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nyeste, K., Zulkipli, N., Uzochukwu, I.E. et al. Assessment of trace and macroelement accumulation in cyprinid juveniles as bioindicators of aquatic pollution: effects of diets and habitat preferences. Sci Rep 14, 11288 (2024). https://doi.org/10.1038/s41598-024-61986-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-61986-4
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
