
Abstract
Exposure to stress has profound, but complex, actions on motivated behavior and decision-making. These effects are central to core symptoms of a number of psychiatric disorders that are precipitated or augmented by stress, such as depressive disorders and substance use disorders. Studying the neural substrates of stress's effects on motivation has revealed that stress affects multiple targets on circuits throughout the brain using diverse molecular signaling processes. Moreover, stress does not have unitary effects on motivated behavior, but differences in the intensity, duration, intermittency, controllability and nature of the stressor produce qualitatively and quantitatively different behavioral endpoints. Unsurprisingly, the results of neuroscientific investigations into stress and motivation often open more questions than they resolve. Here we discuss contemporary results pertaining to the neural mechanisms by which stress alters motivation, identify points of contention and highlight integrative areas for continuing research into these multifaceted complexities.
Similar content being viewed by others

Main
Stressors engage an array of neural and endocrine systems, causing the mobilization of physiological and psychological resources that allow an organism to respond to the present challenge to its homeostasis and overall well-being. Although the subjectively aversive qualities of stress are often emphasized, responses to acute stressors are generally adaptive, beneficial to the individual and potentially critical for survival. Moreover, stress itself is not universally aversive, as we even actively seek out stress under the right circumstances. Many individuals find the stimulation of thrill rides, scary movies and rough-and-tumble play to be invigorating. Notably, such stressors tend to be relatively mild and transient and typically do not entail a complete loss of control. It is now widely recognized that severe or chronic stress, particularly when coupled with a lack of predictability or perceived lack of control, can cause a variety of long-lasting physiological changes that wreak havoc on multiple organ systems as well as contributing to cognitive and affective deficits characteristic of many neuropsychiatric disorders. Although much research has focused on stress effects on hippocampal-dependent memory functions1 and emotional processing related to fear and anxiety2, reward processing and decision-making has become an important domain for stress-related research. Chronic stress induction procedures, including, but not limited to, chronic mild stress3,4 and chronic social defeat stress5,6,7, are commonly used as rodent models for the induction of depressive-like symptoms such as anhedonia, social withdrawal and behavioral despair, as indicated by decreases in sucrose preference and social interaction and increased immobility in forced swim or tail suspension assays, respectively. Aside from the inherent limitations and challenges of fully modeling in rodents any psychiatric disorders that themselves are heterogeneous and imprecisely defined in humans8,9,10,11,12, these stress-induction procedures nevertheless robustly affect several forms of motivated behavior that are amenable to precise circuit manipulation with contemporary genetic techniques13,14,15.
In the current Review, we highlight recent studies examining how stress affects decision-making and general motivational processes, as understanding how stress leads to reprioritization in these basic behavioral domains is a prerequisite to gaining greater insight into the contributions of stress to various forms of psychopathology.
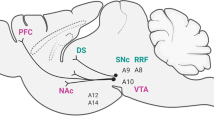
First, we focus on the contributions of dopamine-releasing neurons in the ventral tegmental area (VTA) as well as consequences immediately downstream in the nucleus accumbens (NAc). Mesolimbic dopamine is widely considered to be critical for learning the incentive value of stimuli or actions such that these learned values can be used by the organism to guide its future behavior16,17,18,19,20,21,22. This system thereby contributes to basic motivational processes through facilitating reward seeking, particularly that requiring animals to overcome behavioral challenges such as effortful response costs23,24,25,26,27,28,29. Moreover, mesolimbic dopamine also has been implicated in social aspects of motivated behavior30,31,32,33. Thus, much of our review focuses on studies investigating how a variety of stress-induction procedures influence multiple measures of motivated behavior through alterations in VTA dopamine neurons and the NAc. Recent work on this topic has not only described the nature of structural and physiological changes engendered by stress in this mesolimbic circuitry, but has also begun to reveal the specific neurotransmitter receptors, channels and intracellular signaling molecules underlying these stress-induced alterations.
Second, we discuss work investigating how stress affects decision-making by influencing which of multiple valuation systems organisms use to guide their selection of actions. Although this literature currently lacks the same level of molecular detail regarding the stress-induced systems-level alterations that have been identified in corticostriatal circuitry implicated in action selection, these investigations have spanned both human and nonhuman animal work and utilize sophisticated behavioral designs that are nonetheless amenable to more mechanistic future investigations. Across both sections we discuss current evidence pertaining to the complex interactions between multiple neural systems underlying motivated behavior and stress responsivity (Fig. 1), and we identify apparent inconsistencies in the literature and underexplored areas that are ripe for future investigations.

Kim Caesar/Nature Publishing Group
Select works discussed in this review include stress effects on motivated behaviors mediated by dopamine neurons in the VTA34,35,36,37, their downstream targets in the NAc55,56,57,58,59,60,61, and interactions with neuropeptides such as CRF46,52,57 and BDNF7,38,57 in each region. The second section includes discussion of stress effects on different forms of instrumental behavior and corresponding structural and functional alterations in corticostriatal circuitry supporting these behaviors81,82,83,84,85,86,87,88,89, including regions such as the mPFC, OFC, dorsomedial striatum (DMS) and dorsolateral striatum (DLS).
Stress effects on general motivational processes
The mesolimbic dopamine system has a central role in the control of motivated behavior. Recent studies examining different forms of motivated behavior following protracted stress manipulations have revealed that midbrain dopamine neurons have complex and perhaps contradictory roles in the behavioral consequences of different types of stressors34,35,36. Recent work from Tye and colleagues demonstrates how chronic stress can alter behavior via modulation of mesolimbic dopamine system activity34. Specifically, 8–12 weeks of chronic mild stress caused a reduction in sucrose preference during a two-bottle choice test and a reduction in time spent struggling during a tail suspension test, both of which were restored to the levels of non-stressed control mice by acute phasic (30 Hz) stimulation of dopamine neurons in the ventral tegmental area (VTA) every 5 s throughout periods of these behavioral assays34. Likewise, inhibiting VTA dopamine neurons in stress-naive control animals reduced sucrose preference and increased immobility. These causal manipulations are consistent with the observation that chronic mild stress causes a 50% reduction in spontaneously active VTA dopamine neurons, mediated by a pathway involving the basolateral amygdala and ventral pallidum37. Collectively these findings support the notion that stress-induced reductions in mesolimbic dopamine activity disrupt these projections' normal contributions to motivation and appetitive behavior16,27,29.
In contrast, a series of studies from Han and colleagues revealed a distinct and counterintuitive role for mesolimbic dopamine in mediating animals' susceptibility to the behavioral alterations resulting from social defeat stress. Following 10 d of repeated social defeat stress, a subset of mice exhibited avoidance in a social interaction assay and were designated as 'susceptible' to this defeat stress38. These susceptible mice also demonstrated reduced sucrose preference compared with unsusceptible mice and non-stressed controls38. Rather than showing reduced population activity, however, VTA dopamine neurons of susceptible mice actually exhibited an increased firing rate36,38,39. In previously unsusceptible mice, phasic (20 Hz) stimulation of VTA neurons during a second social interaction test caused social avoidance and reduced subsequent sucrose preference35. Likewise, in a 2-d defeat procedure that is subthreshold for inducing the susceptible phenotype, phasic stimulation of VTA dopamine neurons either during the defeat bouts or during the social interaction test caused social avoidance and reduced subsequent sucrose preference, and phasic stimulation during the latter time point caused a lasting increase in the intrinsic excitability of these neurons35. Following the full 10-d defeat procedure, VTA dopamine neurons of both susceptible and unsusceptible mice exhibited increased excitatory hyperpolarization-activated cation channel current (Ih) relative to non-stressed control mice36,39, consistent with the previous finding that the stress-related peptide corticotropin-releasing factor (CRF) increases VTA dopamine neuron firing rates by enhancing Ih current40. However, unsusceptible mice also showed upregulation of several voltage-gated potassium channels in the VTA38 and corresponding increases in potassium channel currents36 that may promote resilience by normalizing the excitability of these dopamine neurons following repeated defeat stress. Indeed, viral overexpression of the inwardly rectifying potassium channel Kir2.1 in the VTA of susceptible mice reduced the firing rate of VTA dopamine neurons and eliminated behavioral avoidance in a subsequent social interaction test38, as did administration of an Ih inhibitor into the VTA39.
Notably, although potentiating Ih currents pharmacologically might have been expected to exacerbate the behavioral alterations exhibited by susceptible mice, this manipulation actually reversed their social avoidance and increased sucrose preference36. Similar restorative effects were observed following phasic stimulation of VTA dopamine neurons for 20-min periods administered outside of any task context for 5 d. Although stimulation of VTA dopamine neurons in this instance did promote subsequent social approach and appetitive behavior, overall this finding is consistent with the larger body of work suggesting that hyperexcitability of mesolimbic dopamine neurons contributes to social and motivational deficits following defeat stress. In particular, both the pharmacological and stimulation treatments seemed to engage mechanisms of homeostatic plasticity, increasing potassium channel currents and thereby reducing the firing rates of VTA dopamine neurons in these previously susceptible mice. Thus, this series of studies demonstrated remarkable internal consistency and mechanistic detail even though the notion that increased mesolimbic dopamine neuron activity promotes susceptibility to stress may be surprising.
These studies raise an apparent conundrum regarding the contributions of mesolimbic dopamine to stress-induced effects on motivated behavior. In the one common behavioral measure between these studies, both chronic mild stress and repeated social defeat stress (for a subset of animals) caused reductions in sucrose preference. But these distinct stressors caused opposing changes in VTA dopamine neuron activity, and causal manipulations of these neurons either alleviated or exacerbated the behavioral consequences of each stressor. On the one hand, chronic mild stress reduced the number of spontaneously active VTA dopamine neurons37 and phasic stimulation during the two-bottle choice test restored sucrose preference to control levels34. On the other hand, susceptibility to social defeat stress was associated with increased VTA dopamine neuron firing rates36,38,39 and phasic stimulation during subthreshold defeat or during the post-defeat interaction test in unsusceptible mice was sufficient to cause subsequent reductions in sucrose preference35. These divergent contributions of dopamine to changes in sucrose preference following each type of stressor are particularly intriguing given that others have reported that congenitally dopamine-deficient animals retain the ability to form and express preferences for sucrose41 and pharmacological depletion or antagonism of mesolimbic dopamine does not alter sucrose preference following extensive training in stress-naive animals42. Nevertheless, both series of stress studies34,35,36 provided causal evidence that acute manipulations of mesolimbic dopamine altered performance in this simple assay and mediated motivational changes following the different forms of stress.
Moreover, the physiological consequences of social defeat stress were specific to VTA neurons projecting to the NAc; indeed, VTA neurons projecting to the medial prefrontal cortex (mPFC) exhibited opposite physiological changes following social defeat stress35,36. Although the projection targets of dopamine neurons affected by chronic mild stress have not been fully examined, pharmacological blockade of dopamine receptors in the NAc increased immobility in the tail suspension test and prevented any stimulation-induced increase in struggling34, implicating mesolimbic dopamine, but leaving the question of whether mesocortical dopamine has a role in the consequences of chronic mild stress open. Further direct comparison across studies is challenging given the numerous methodological differences—from the timing of dopamine neuron stimulation relative to the stress induction and behavioral tests to the pathway- and cell type–specificity of each study's approaches, and a variety of other subtle, but potentially important, differences (for an extensive discussion, see ref. 43)—but the severity and type of stressors may have contributed to these divergent findings. Indeed, Grace and colleagues have shown that, in addition to chronic mild stress reducing VTA dopamine neuron population activity37, acute stressors such as restraint or repeated foot shock increased this population activity44,45. These stressor-specific changes in population activity, tonic firing rate or excitability of VTA dopamine neurons in anesthetized animals or ex vivo slice preparations permit inferences and the generation of new hypotheses regarding the discrepant behavioral effects observed, but there is not always a straightforward relationship between these measures of basal activity and the effect of specific behavioral events on dopamine transmission. For example, although CRF in the VTA increased dopamine neuron firing rates40, intra-VTA CRF attenuated the phasic dopamine response to reward delivery without affecting the response to presentation of reward-predictive cues in behaving animals46. Indeed, CRF in the VTA differentially affected mesolimbic dopamine release evoked by stimulation of distinct inputs to the VTA. Thus, in conjunction with the growing emphasis on efferent projection–specific heterogeneity of midbrain DA neurons35,36,47,48,49, further characterization of the specific roles of the many diverse inputs to dopamine neurons49,50,51 will be critical for gaining a more complete understanding of how different types of stressors affect this circuitry and influence motivated behavior.
In addition to the regulation of firing of midbrain dopamine neurons, stress also can alter mesolimbic dopamine transmission at the terminal level in the NAc. CRF receptors are found at dopamine neuron terminals in the NAc, and their activation by CRF augments evoked dopamine release in a concentration-dependent manner in stress-naive animals52. Moreover, CRF infusion into the NAc of behaving animals induces a preference for the CRF-paired location52, enhances the ability of appetitive Pavlovian-conditioned stimuli to invigorate instrumental reward-seeking behavior53 and accelerates social bond formation54. Thus, this stress-related peptide acts in the NAc to positively mediate appetitive behavior in stress-naive animals. However, after repeated forced swim stress, CRF in the NAc no longer increases dopamine release and now induces a conditioned place aversion rather than preference. This attenuation of CRF-potentiated dopamine release lasts at least 90 d after the stress induction and is mediated by glucocorticoid receptors, as CRF retains the ability to increase dopamine release when mice are pretreated with the glucocorticoid receptor antagonist RU486 before each swim stress session. Although the site of action and mechanism through which glucocorticoids regulate this CRF-dopamine interaction awaits further investigation, this work has begun to reveal a neural substrate through which repeated stress induces a switch in the affective significance of this stress-related signal's ability to regulate a critical modulator of motivated behavior. Other stress effects in the NAc include dendritic hypertrophy and increased spine density in medium spiny neurons (MSNs) following chronic mild stress as well as increased expression of several genes, including brain-derived neurotropic factor (Bdnf)55. Increased dendritic spine density56 and BDNF levels38 have been observed in the NAc of susceptible mice following repeated social defeat, exemplifying a point of convergence between these stress induction procedures. Notably, increased BDNF levels were also found in postmortem NAc tissue from depressed human patients38. CRF signaling in the NAc is necessary for this social defeat stress–induced BDNF increase in susceptible mice, as both social avoidance and the increase in BDNF were blocked by intra-NAc administration of the CRF receptor antagonist alpha-helical CRF57.
Although the combination of intra-NAc CRF and phasic stimulation of VTA neurons was sufficient to increase NAc BDNF levels in stress-naive mice, their social interaction behavior remained unaffected, indicating that increased BDNF alone is not sufficient to induce social avoidance in the absence of actual stress experience57. Moreover, the source of the BDNF necessary for defeat stress–induced social avoidance seems to be the VTA rather than in the NAc itself, as Bdnf mRNA levels in the NAc were unchanged in susceptible mice relative to unsusceptible and control mice, and BDNF knockdown in the VTA, but not NAc, reduced social avoidance and increased sucrose preference following repeated defeat stress38.
Although these studies reporting increased MSN spine density did not specify which subpopulations exhibited these stress-induced alterations, additional work has demonstrated distinct morphological and physiological consequences in different MSN populations containing predominantly D1 versus D2 dopamine receptors. Specifically, D1-MSNs of mice susceptible to social defeat stress exhibited increased intrinsic excitability and a decreased frequency of miniature excitatory postsynaptic currents (mEPSCs), whereas D2-MSNs from susceptible mice had unchanged intrinsic excitability and an increase in mEPSC frequency58. In contrast, unsusceptible, but not susceptible, mice had increased unitary EPSC amplitude specifically onto mushroom spines of D1-MSNs and decreased EPSC amplitude onto mushroom spines of D2-MSNs59. Accordingly, specific manipulation of each pathway can differentially exacerbate or alleviate stress-induced behavioral alterations58,60. Adding even greater complexity to the roles of these circuits in general motivational processes, these pathway-specific manipulations resulted in dissociable effects on distinct behavioral measures. For example, high-frequency stimulation of D2-MSNs for several days before and immediately after subthreshold defeat stress increased subsequent social avoidance, but had no effect on sucrose preference58. Likewise, overexpression of β-catenin in the NAc prevented social avoidance and reduced forced-swim immobility following repeated social defeat stress, with the pro-social effect of β-catenin being mediated specifically by D2-MSNs and not by D1-MSNs, but this manipulation again did not affect sucrose preference60. In contrast, repeated (but not acute) high-frequency stimulation of D1-MSNs after repeated defeat stress increased subsequent social interaction time and sucrose preference in previously susceptible mice, and inhibition of D1-MSNs induced social avoidance and reduced sucrose preference in previously unsusceptible mice, whereas neither manipulation of D2-MSNs had any effect58. Notably, D1-MSN stimulation also increased subsequent sucrose preference in stress-naive mice.
Although the long-term physiological consequences of these circuit manipulations remain unknown, a recent study provided mechanistic insight into a feeding-related peptide whose activity at D1-MSNs in the NAc contributes to stress-induced reductions selectively in sucrose preference without affecting escape-related behavior in tail suspension or forced swim tests61. Specifically, 8 d of restraint stress caused reductions in sucrose preference and increases in immobility along with changes in synaptic physiology at D1-MSNs, but not D2-MSNs, that included NMDA receptor–dependent long-term depression, reduced AMPA/NMDA ratios and endocytosis of GluA2-containing AMPA receptors. Activation of melanocortin 4 receptor (MC4R) in the NAc by 2–3-h incubation with the endogenous MC4R ligand α-melanocyte–stimulating hormone recapitulated these stress-induced physiological changes in D1-MSNs without affecting D2-MSNs. Knocking down MC4Rs or blocking AMPA receptor endocytosis in the NAc selectively prevented the stress-induced reduction in sucrose preference without affecting stress-induced increases in immobility during tail suspension or forced swim tests. Notably, this repeated restraint stress also caused a reduction in food consumption and consequent cessation of normal weight gain, which were also prevented by these manipulations of NAc MC4R signaling. Other feeding-related signaling molecules potentially interact with stress signals and may affect motivated behaviors beyond food intake and sucrose preference. For example, the orexigenic hormone ghrelin is increased following repeated social defeat stress62,63, and not only does it mediate the stress-induced preference for high-fat food63, but it also reduces forced-swim immobility62. Given the recent resurgence of studies characterizing the detailed circuitry involved in appetite regulation using precise molecular genetic techniques to dissect the multitude of excitatory, inhibitory, neuropeptidergic and other modulatory interactions between a variety of hypothalamic and other limbic and brainstem nuclei64,65,66,67,68,69,70,71,72, further examination of the reciprocal interactions between stress and feeding-related circuits represents an important area for ongoing research73.
Collectively, these studies demonstrate that stress effects on general motivational processes remain a complex topic, but they also point toward promising avenues for future research that will continue to provide mechanistic insight into the molecular substrates contributing to these processes. In particular, the studies focusing on mesolimbic dopamine discussed above have highlighted the importance of future investigations of different types of stressors in controlled studies using otherwise equivalent neural perturbations and behavioral measures. Although the conclusions regarding the role of dopamine in mediating stress effects on motivated behavior differed across these previous studies, the investigators in each study demonstrated internally consistent effects of dopamine manipulations across multiple assays of motivated behaviors affected by stress. In contrast with these relatively coherent behavioral clusters or syndromes affected by mesolimbic dopamine manipulations, the subsequent examinations of pathway-specific manipulations immediately downstream in the NAc have revealed dissociable effects on these same behavioral measures. To the extent that certain behavioral assays or stress-induction procedures may be more relevant to or valid for modeling neuropsychiatric disorders in nonhuman animals, the molecular detail provided by these studies may contribute to the development of improved treatments that more selectively target specific symptoms and, ideally, the underlying etiological mechanisms causing a particular individual's pathology. As these findings also highlight that motivated behavior is of course by no means a unitary process, there remains the critical challenge of refining the existing behavioral assays to better understand what specific psychological processes underlie any particular behavioral measure and how they are affected by a given molecular or circuit-level manipulation.
A promising complementary way forward entails using these mechanistic approaches in conjunction with more sophisticated behavioral procedures examining more specific aspects of decision-making processes. There is a growing body of literature on stress effects on economic decision-making, much of which comes from a top-down perspective that has not yet been able to capitalize on the molecular studies of stress effects on general motivated behaviors as building blocks. This discontinuity provides a major research opportunity to consolidate and reconcile the literature on the specific molecular processes underlying general motivated behavior and the decision-making literature, the latter of which has a less detailed analysis of the neural substrates but more precision in the cognitive functions affected by stress. Nonetheless, there are a few examples of studies that have begun to transcend the levels of analysis from molecular biology to cognition. Following acute restraint stress, animals exhibit decremented responding for food rewards, a change in motivation that is mediated by the action of endogenous CRF in the VTA46. This effect can be recapitulated by administration of exogenous CRF into the VTA, which decreases the amplitude of phasic dopamine release in the NAc in response to food delivery46. Indeed, direct manipulations of dopamine transmission in the NAc, such as local application of dopamine receptor antagonists or dopamine neurotoxins, produce a similar reduction in responding for food rewards74,75, even though dopamine depletion does not alter hedonic responses to reward delivery76. This action is argued to be a selective effect on effort-based decision-making, as, when tested in concurrent decision-making tasks, these manipulations of NAc dopamine bias animals' preferences away from rewards that are available for a high response requirement toward otherwise less desirable rewards (smaller quantity or less palatable food) available for a minimal response requirement25,26,27,28. Thus, these data collectively suggest that following acute restraint stress, CRF acts in the VTA to reduce the excitability of dopamine neurons to reward delivery, decreasing dopamine release in the NAc, which selectively alters decision-making policies so that animals prefer rewards that are available at lower response costs (Fig. 2), that is, they exhibit anergia25. When animals were tested in an effort-based decision-making task following acute restraint stress, this prediction held true77. Thus, these studies demonstrate transparency to how some of the molecular changes associated with stress effects on general motivation processes could have direct effect on decision-making.

Kim Caesar/Nature Publishing Group
Acute restraint stress biases decision-making away from high-effort, high-reward options when a low-reward option is concurrently available for lower effort77. The proposed mechanism for this effect is by reduction of reward-evoked dopamine release in the NAc as a result of release of CRF into the VTA during acute stress46.
Stress and multiple valuation processes
To this point, we have treated reward valuation as a solitary process to understand the effects of stress on motivation. However, it is commonly accepted that dual, or even multiple, sets of valuation processes exist, differing in their cognitive demands and the resultant flexibility of behaviors controlled by different underlying associative structures. One such example of dual valuation processes is identified in instrumental behavior based on the representation of current goal information as opposed to purely stimulus-elicited responses, designated as goal-directed versus habitual, respectively. Across a series of behavioral, pharmacological and neuroimaging studies using human participants subjected to an acute stressor that combined both physical and social aspects (the socially evaluated cold pressor test), Schwabe, Wolf and colleagues repeatedly demonstrated that this acute stress biased participants toward habitual behavior rather than goal-directed instrumental behavior. These researchers used an instrumental conditioning task developed for use with humans undergoing functional magnetic resonance imaging78 (fMRI) and adapted from similar behavioral tasks employed with rodents79,80. Briefly, participants first learned to associate different actions with specific reinforcer outcomes (for example, chocolate milk or orange juice) and then were given the opportunity to consume one reinforcer to satiety to selectively devalue that outcome. In a subsequent post-devaluation test session in which neither outcome was delivered, participants' choices revealed whether their behavior was under goal-directed or habitual control: a preference for the action associated with the non-devalued option was indicative of goal-directed behavior, whereas an equivalent preference for the actions associated with the devalued and non-devalued options was indicative of habitual behavior. Schwabe et al. demonstrated that, although non-stressed controls exhibited goal-directed behavior in this task, the acute physical and social stress manipulation biased participants toward habitual control, as their behavior was insensitive to this change in action-outcome value in the first block of trials in the post-devaluation test session81,82. This stress-induced insensitivity to outcome-selective devaluation was observed whether the stress induction occurred before initial acquisition of the instrumental responses81 or just before the post-devaluation extinction test session82. The latter finding indicated that stress affected performance rather than solely disrupting acquisition of goal-directed behavior, although the bias toward habitual behavior was longer lasting when the stressor occurred before acquisition. Notably, when the acute stressor occurred before initial acquisition, significantly fewer stressed participants than non-stressed controls were able to report explicit knowledge of the action-outcome contingencies81, but this explicit task knowledge was unaffected when the stressor occurred after acquisition82.
Following the observation that stress-induced changes in cortisol levels significantly correlated with individuals' probability of choosing the devalued option82, the authors subsequently investigated the contribution of specific stress-related neuroendocrine systems to this bias toward habitual behavior83,84,85. In addition to this association between glucocorticoids and stress-induced habitual behavior82, the noradrenergic system also has been implicated in this bias toward habitual behavior, as administration of the β-adrenergic receptor antagonist propranolol before the stress manipulation prevented the shift toward habitual behavior induced by acute stress84. Moreover, exogenous activation of both systems concurrently was sufficient to bias participants' decisions toward habitual behavior in the absence of any overt stressor83,85. Specifically, in a double-blind, placebo-controlled design, participants were administered either the synthetic glucocorticoid hydrocortisone, the anxiogenic α2-adrenoceptor antagonist yohimbine, both hydrocortisone and yohimbine, or placebo only, each in the absence of any stress manipulation. The combined administration of hydrocortisone and yohimbine rendered participants' behavior habitual. However, neither hydrocortisone nor yohimbine treatment on its own caused any difference from the placebo controls, as these three groups' behavior each remained goal-directed. A previous fMRI study using this behavioral task highlighted the recruitment of orbitofrontal (OFC) and ventromedial prefrontal cortex in the goal-directed selection of non-devalued actions over devalued actions78. Although neither hydrocortisone nor yohimbine alone affected the activation observed in these ventral prefrontal regions, the combined treatment significantly disrupted this activity pattern85. These neuroimaging findings were consistent with the requirement of concurrent activation of glucocorticoid and noradrenergic systems to disrupt goal-directed behavior and induce a shift toward less-flexible, habitual behavior that was insensitive to changes in outcome value.
These effects of acute stress and exogenous activation of stress-related neuroendocrine systems were partially replicated in a recent rodent study86. Following either an acute 60-min restraint stress or combined treatment with corticosterone and yohimbine, rats remained sensitive to outcome-selective devaluation, indicating that these manipulations were not sufficient to render the animals' behavior habitual, in contrast with the previous human studies. However, a more severe stressor (a 60-min sequence of 20-min restraint, 20-min on an elevated platform and another 20-min restraint plus five tail shocks, all with loud music playing in a brightly lit novel room during the rats' dark cycle) did disrupt animals' sensitivity to devaluation. Although these findings in rats were only partially consistent with those from the human studies described above, it is noteworthy that these manipulations in rats were all conducted just before the post-devaluation choice tests, well after initial acquisition of the action-outcome contingencies. Although the stress manipulation in these human studies did bias participants toward habitual behavior regardless of whether the stress induction occurred before initial acquisition or only just before the post-devaluation test, the effects were stronger and longer lasting when the stress manipulation occurred before acquisition81,82. Moreover, these studies collectively highlight that differences in stressor severity or other parameters potentially could account for the inconsistencies observed across studies. In the studies with human subjects, stress-induced increases in habitual behavior have consistently been observed when social and physical stressors were administered in combination, but how these stressors might affect behavior when given in isolation has not been tested. In the rodent study, only the combined stressor, but not the single restraint stress, significantly disrupted goal-directed behavior86. Surprisingly, however, both stressor types caused comparable increases in the rats' plasma corticosterone levels, perhaps indicating that this physiological metric is not sufficiently sensitive to detect the differences in psychological severity between these two stress manipulations. Although further studies are required to more systematically investigate the effects of stressor type and severity on goal-directed versus habitual control of behavior, these studies collectively provide initial evidence for certain acute stressors biasing both humans and rodents toward habitual behavior.
Studies examining the effects of chronic stress on instrumental behavior have yielded more consistent results across species, indicating that chronic stress biases animals toward habitual behavior. In one study, rats were subjected to 21 d of chronic unpredictable stress that included randomly interleaved bouts of social defeat, forced swim and restraint stress87. Following 12 d of instrumental training, chronically stressed rats' behavior became insensitive to reinforcer devaluation and changes in action-outcome contingencies, indicative of habitual behavior, whereas non-stressed control rats remained goal-directed. These stressed rats exhibited dendritic atrophy in the medial prefrontal cortex and dorsomedial striatum, coupled with hypertrophy in the dorsolateral striatum. Insensitivity to outcome devaluation and parallel corticostriatal alterations were also observed in human participants following prolonged stress88. This fMRI study examined instrumental behavior in medical students who had recently completed their medical residency selection exam (that is, after a long and stressful preparation period) versus a control group of medical students who had not recently prepared for this exam. Not only did chronic stress bias the former group toward habitual behavior, but it was also associated with changes in neural structure and function: morphological hypertrophy and increased activity were observed in the putamen, whereas atrophy and reduced activity were observed in the caudate of the stressed group relative to controls, and atrophy was also observed in the ventromedial prefrontal cortex of the stressed group. Another rodent study demonstrated that chronic delivery of corticosterone via drinking water recapitulated this bias toward habitual behavior, as corticosterone-treated animals became insensitive to both reinforcer devaluation and contingency degradation89. This study also demonstrated that glucocorticoid receptors and prefrontal BDNF have dissociable roles in habit formation and general motivation, respectively. Thus, these experiments lay the foundation for future investigations of molecular mechanisms underlying stress-induced biases toward habitual behavior. Moreover, such investigations represent a promising avenue for integration with additional recent human studies suggesting that stress preferentially disrupts prefrontal-dependent cognitive computations contributing to goal-directed decision-making90, as well as potentially aligning more broadly with the extensive body of cross-species work investigating the mechanisms through which stress alters prefrontal circuitry supporting working memory and other forms of cognitive flexibility91,92,93,94.
Summary and conclusions
In this Review, we have not comprehensively cataloged the extensive literature on the effects of stress on motivation and decision-making, but instead highlighted contemporary areas of research and the key parameters that may affect their interpretation. As we have noted, the effects of stress on neural substrates related to motivated behavior are numerous and complex, and it is likely that the existing knowledge is just the tip of the iceberg. As such, the existing data at times seem to be confusing and even contradictory, no doubt because they are incomplete. We have emphasized that the different stress-induction protocols may have important bearing on the results, perhaps most importantly differences in the intensity, duration, intermittency and controllability of stressor exposure and differences between social and physical stress. Adding to the complexity is the concept of multiple valuation systems and the effects of stress thereon. Because stress biases the arbitration between these competing valuation systems, the cognitive substrates used by stress-naive and stress-exposed individuals may be fundamentally different. In this regard, it is also noteworthy that other forms of behavioral and cognitive flexibility that are relevant to, but not directly subsumed in, motivation per se are also affected by stress, including those identified in spatial, working memory and set-shifting tasks91,92,93,94,95,96,97,98,99. Despite these impediments, research into the neural substrates underlying stress effects on motivated behavior has been fruitful and continues to grow as a topic of research. It is clear that future studies will need to use different types of stressor exposure under otherwise identical conditions and to compare these effects using common metrics and analyses. Understanding this relationship better will no doubt prove to be increasingly complex, but it is a critically important endeavor, as the link between stress and motivation is instrumental in the most prevalent psychiatric disorders, including substance use disorders and depressive disorders.
References
McEwen, B.S. Stress and hippocampal plasticity. Annu. Rev. Neurosci. 22, 105–122 (1999).
Rodrigues, S.M., Ledoux, J.E. & Sapolsky, R.M. The influence of stress hormones on fear circuitry. Annu. Rev. Neurosci. 32, 289–313 (2009).
Willner, P., Towell, A., Sampson, D., Sophokleous, S. & Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology (Berl.) 93, 358–364 (1987).
Willner, P. Chronic mild stress (CMS) revisited: consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology 52, 90–110 (2005).
Kudryavtseva, N.N., Bakshtanovskaya, I.V. & Koryakina, L.A. Social model of depression in mice of C57BL/6J strain. Pharmacol. Biochem. Behav. 38, 315–320 (1991).
Golden, S.A., Covington, H.E., Berton, O. & Russo, S.J. A standardized protocol for repeated social defeat stress in mice. Nat. Protoc. 6, 1183–1191 (2011).
Berton, O. et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 311, 864–868 (2006).
Insel, T.R. & Landis, S.C. Twenty-five years of progress: the view from NIMH and NINDS. Neuron 80, 561–567 (2013).
Anisman, H. & Matheson, K. Stress, depression, and anhedonia: caveats concerning animal models. Neurosci. Biobehav. Rev. 29, 525–546 (2005).
Nestler, E.J. & Hyman, S.E. Animal models of neuropsychiatric disorders. Nat. Neurosci. 13, 1161–1169 (2010).
Berton, O., Hahn, C.-G. & Thase, M.E. Are we getting closer to valid translational models for major depression? Science 338, 75–79 (2012).
Monteggia, L.M., Malenka, R.C. & Deisseroth, K. Depression: the best way forward. Nature 515, 200–201 (2014).
Deisseroth, K. Circuit dynamics of adaptive and maladaptive behavior. Nature 505, 309–317 (2014).
Huang, Z.J. & Zeng, H. Genetic approaches to neural circuits in the mouse. Annu. Rev. Neurosci. 36, 183–215 (2013).
Luo, L., Callaway, E.M. & Svoboda, K. Genetic dissection of neural circuits. Neuron 57, 634–660 (2008).
Fields, H.L., Hjelmstad, G.O., Margolis, E.B. & Nicola, S.M. Ventral tegmental area neurons in learned appetitive behavior and positive reinforcement. Annu. Rev. Neurosci. 30, 289–316 (2007).
Berridge, K.C. From prediction error to incentive salience: mesolimbic computation of reward motivation. Eur. J. Neurosci. 35, 1124–1143 (2012).
Cohen, J.Y., Haesler, S., Vong, L., Lowell, B.B. & Uchida, N. Neuron-type-specific signals for reward and punishment in the ventral tegmental area. Nature 482, 85–88 (2012). In this work, the authors recorded the firing patterns of neurons in the VTA during reward and punishment procedures. The neurons were neurochemically identified by optogenetics with cell type–specific expression of channelrhodopsin-2.
Nicola, S.M. The nucleus accumbens as part of a basal ganglia action selection circuit. Psychopharmacology (Berl.) 191, 521–550 (2007).
du Hoffmann, J. & Nicola, S.M. Dopamine invigorates reward seeking by promoting cue-evoked excitation in the nucleus accumbens. J. Neurosci. 34, 14349–14364 (2014).
Flagel, S.B. et al. A selective role for dopamine in stimulus-reward learning. Nature 469, 53–57 (2011). This paper demonstrated that multiple valuation systems have different dependencies on dopamine transmission.
Phillips, P.E.M., Stuber, G.D., Heien, M.L.A.V., Wightman, R.M. & Carelli, R.M. Subsecond dopamine release promotes cocaine seeking. Nature 422, 614–618 (2003).
Floresco, S.B., Tse, M.T.L. & Ghods-Sharifi, S. Dopaminergic and glutamatergic regulation of effort- and delay-based decision making. Neuropsychopharmacology 33, 1966–1979 (2008).
Nicola, S.M. The flexible approach hypothesis: unification of effort and cue-responding hypotheses for the role of nucleus accumbens dopamine in the activation of reward-seeking behavior. J. Neurosci. 30, 16585–16600 (2010).
Salamone, J.D., Correa, M., Farrar, A. & Mingote, S.M. Effort-related functions of nucleus accumbens dopamine and associated forebrain circuits. Psychopharmacology (Berl.) 191, 461–482 (2007).
Salamone, J.D., Correa, M., Farrar, A.M., Nunes, E.J. & Pardo, M. Dopamine, behavioral economics, and effort. Front. Behav. Neurosci. 3, 13 (2009).
Salamone, J.D. & Correa, M. The mysterious motivational functions of mesolimbic dopamine. Neuron 76, 470–485 (2012).
Treadway, M.T. et al. Dopaminergic mechanisms of individual differences in human effort-based decision-making. J. Neurosci. 32, 6170–6176 (2012).
Ikemoto, S. & Panksepp, J. The role of nucleus accumbens dopamine in motivated behavior: a unifying interpretation with special reference to reward-seeking. Brain Res. Brain Res. Rev. 31, 6–41 (1999).
Gunaydin, L.A. et al. Natural neural projection dynamics underlying social behavior. Cell 157, 1535–1551 (2014).
Aragona, B.J. et al. Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds. Nat. Neurosci. 9, 133–139 (2006).
Robinson, D.L., Heien, M.L.A.V. & Wightman, R.M. Frequency of dopamine concentration transients increases in dorsal and ventral striatum of male rats during introduction of conspecifics. J. Neurosci. 22, 10477–10486 (2002).
Willuhn, I. et al. Phasic dopamine release in the nucleus accumbens in response to pro-social 50 kHz ultrasonic vocalizations in rats. J. Neurosci. 34, 10616–10623 (2014).
Tye, K.M. et al. Dopamine neurons modulate neural encoding and expression of depression-related behavior. Nature 493, 537–541 (2013). This study demonstrated reduced dopamine function following chronic mild stress and showed that restoring dopamine function reversed depression-like behaviors.
Chaudhury, D. et al. Rapid regulation of depression-related behaviours by control of midbrain dopamine neurons. Nature 493, 532–536 (2013). This paper showed that phasic activity of VTA dopamine neurons increased the susceptibility for social-defeat stress to induce a depressive-like phenotype in mice.
Friedman, A.K. et al. Enhancing depression mechanisms in midbrain dopamine neurons achieves homeostatic resilience. Science 344, 313–319 (2014).This paper showed that enhancing VTA dopamine neuron activity in mice susceptible to social-defeat stress reversed depression-like behaviors. This effect was considered to be somewhat paradoxical, as the susceptible mice already exhibited hyperactivity in dopamine neurons.
Chang, C.-H. & Grace, A.A. Amygdala-ventral pallidum pathway decreases dopamine activity after chronic mild stress in rats. Biol. Psychiatry 76, 223–230 (2014). This paper demonstrated that chronic mild stress reduces the number of spontaneously firing midbrain dopamine neurons.
Krishnan, V. et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 131, 391–404 (2007). This work identified molecular differences in the VTA-NAc pathway between individual mice that were susceptible or resilient to social-defeat stress and in postmortem tissue from depressed human patients and controls.
Cao, J.-L. et al. Mesolimbic dopamine neurons in the brain reward circuit mediate susceptibility to social defeat and antidepressant action. J. Neurosci. 30, 16453–16458 (2010).
Wanat, M.J., Hopf, F.W., Stuber, G.D., Phillips, P.E.M. & Bonci, A. Corticotropin-releasing factor increases mouse ventral tegmental area dopamine neuron firing through a protein kinase C-dependent enhancement of Ih. J. Physiol. (Lond.) 586, 2157–2170 (2008). This study identified the cellular mechanism by which CRF increases the basal firing rate of VTA dopamine neurons.
Cannon, C.M. & Palmiter, R.D. Reward without dopamine. J. Neurosci. 23, 10827–10831 (2003). This work demonstrated that reward preferences can be exhibited in animals that do not have intact dopamine transmission.
Pardo, M., López-Cruz, L., Miguel, N.S., Salamone, J.D. & Correa, M. Selection of sucrose concentration depends on the effort required to obtain it: studies using tetrabenazine, D1, D2, and D3 receptor antagonists. Psychopharmacology (Berl.) 232, 2377–2391 (2015).
Lammel, S., Tye, K.M. & Warden, M.R. Progress in understanding mood disorders: optogenetic dissection of neural circuits. Genes Brain Behav. 13, 38–51 (2014).
Valenti, O., Lodge, D.J. & Grace, A.A. Aversive stimuli alter ventral tegmental area dopamine neuron activity via a common action in the ventral hippocampus. J. Neurosci. 31, 4280–4289 (2011).
Valenti, O., Gill, K.M. & Grace, A.A. Different stressors produce excitation or inhibition of mesolimbic dopamine neuron activity: response alteration by stress pre-exposure. Eur. J. Neurosci. 35, 1312–1321 (2012).
Wanat, M.J., Bonci, A. & Phillips, P.E.M. CRF acts in the midbrain to attenuate accumbens dopamine release to rewards but not their predictors. Nat. Neurosci. 16, 383–385 (2013). This paper described afferent-selective regulation of dopamine transmission by CRF in the VTA and demonstrated its role in mediating alterations of motivation following acute restraint stress.
Lammel, S. et al. Unique properties of mesoprefrontal neurons within a dual mesocorticolimbic dopamine system. Neuron 57, 760–773 (2008).
Lammel, S., Ion, D.I., Roeper, J. & Malenka, R.C. Projection-specific modulation of dopamine neuron synapses by aversive and rewarding stimuli. Neuron 70, 855–862 (2011).
Lammel, S. et al. Input-specific control of reward and aversion in the ventral tegmental area. Nature 491, 212–217 (2012). This paper reported that VTA dopamine neurons with distinct projection targets encode reward and aversion.
Watabe-Uchida, M., Zhu, L., Ogawa, S.K., Vamanrao, A. & Uchida, N. Whole-brain mapping of direct inputs to midbrain dopamine neurons. Neuron 74, 858–873 (2012). This work provided a comprehensive brain-wide survey of neurons that synapse onto midbrain dopamine neurons.
Matsui, A., Jarvie, B.C., Robinson, B.G., Hentges, S.T. & Williams, J.T. Separate GABA afferents to dopamine neurons mediate acute action of opioids, development of tolerance, and expression of withdrawal. Neuron 82, 1346–1356 (2014).
Lemos, J.C. et al. Severe stress switches CRF action in the nucleus accumbens from appetitive to aversive. Nature 490, 402–406 (2012). This paper identified a mechanism by which CRF regulates dopamine that is ablated by repeated swim stress.
Peciña, S., Schulkin, J. & Berridge, K.C. Nucleus accumbens corticotropin-releasing factor increases cue-triggered motivation for sucrose reward: paradoxical positive incentive effects in stress? BMC Biol. 4, 8 (2006). This work demonstrated that CRF acts in the NAc to enhance the effects of Pavlovian stimuli on instrumental behavior.
Lim, M.M. et al. CRF receptors in the nucleus accumbens modulate partner preference in prairie voles. Horm. Behav. 51, 508–515 (2007).
Bessa, J.M. et al. Stress-induced anhedonia is associated with hypertrophy of medium spiny neurons of the nucleus accumbens. Transl. Psychiatry 3, e266 (2013). This paper reported that chronic mild stress produced hypertrophy and increased expression of the genes encoding neurotrophins, cell adhesion molecules and synaptic proteins in NAc medium spiny neurons.
Christoffel, D.J. et al. I,B kinase regulates social defeat stress-induced synaptic and behavioral plasticity. J. Neurosci. 31, 314–321 (2011). This study reported that repeated social defeat stress caused alterations in dendritic structure and plasticity in NAc medium spiny neurons, and it identified a molecular signaling pathway involved in these effects.
Walsh, J.J. et al. Stress and CRF gate neural activation of BDNF in the mesolimbic reward pathway. Nat. Neurosci. 17, 27–29 (2014). This study investigated the interactions between mesolimbic dopamine, neurotrophic and stress-related factors, and actual stress exposure in mediating stress-induced social withdrawal.
Francis, T.C. et al. Nucleus accumbens medium spiny neuron subtypes mediate depression-related outcomes to social defeat stress. Biol. Psychiatry 77, 212–222 (2015). This study examined stress-induced changes in electrophysiological properties of specific populations of NAc medium spiny neurons, as well as the dissociable contributions of each subpopulation to distinct forms of motivated behavior disrupted by social defeat stress.
Khibnik, L.A. et al. Stress and cocaine trigger divergent and cell type–specific regulation of synaptic transmission at single spines in nucleus accumbens. Biol. Psychiatry (2015).
Dias, C. et al. β-catenin mediates stress resilience through Dicer1/microRNA regulation. Nature 516, 51–55 (2014). This paper characterized the involvement of a molecular signaling pathway in a specific subpopulation of NAc medium spiny neurons contributing to stress-induced effects on select forms of motivated behavior.
Lim, B.K., Huang, K.W., Grueter, B.A., Rothwell, P.E. & Malenka, R.C. Anhedonia requires MC4R-mediated synaptic adaptations in nucleus accumbens. Nature 487, 183–189 (2012). This paper revealed the mechanisms through which a feeding-related peptide affects stress-induced changes in sucrose preference by altering physiological properties of a specific subpopulation of medium spiny neurons in the NAc.
Lutter, M. et al. The orexigenic hormone ghrelin defends against depressive symptoms of chronic stress. Nat. Neurosci. 11, 752–753 (2008).
Chuang, J.-C. et al. Ghrelin mediates stress-induced food-reward behavior in mice. J. Clin. Invest. 121, 2684–2692 (2011).
Wu, Q., Clark, M.S. & Palmiter, R.D. Deciphering a neuronal circuit that mediates appetite. Nature 483, 594–597 (2012).
Carter, M.E., Soden, M.E., Zweifel, L.S. & Palmiter, R.D. Genetic identification of a neural circuit that suppresses appetite. Nature 503, 111–114 (2013).
Atasoy, D., Betley, J.N., Su, H.H. & Sternson, S.M. Deconstruction of a neural circuit for hunger. Nature 488, 172–177 (2012).
Betley, J.N., Cao, Z.F.H., Ritola, K.D. & Sternson, S.M. Parallel, redundant circuit organization for homeostatic control of feeding behavior. Cell 155, 1337–1350 (2013).
Jennings, J.H., Rizzi, G., Stamatakis, A.M., Ung, R.L. & Stuber, G.D. The inhibitory circuit architecture of the lateral hypothalamus orchestrates feeding. Science 341, 1517–1521 (2013).
Jennings, J.H. et al. Visualizing hypothalamic network dynamics for appetitive and consummatory behaviors. Cell 160, 516–527 (2015).
Krashes, M.J. et al. An excitatory paraventricular nucleus to AgRP neuron circuit that drives hunger. Nature 507, 238–242 (2014).
Shah, B.P. et al. MC4R-expressing glutamatergic neurons in the paraventricular hypothalamus regulate feeding and are synaptically connected to the parabrachial nucleus. Proc. Natl. Acad. Sci. USA 111, 13193–13198 (2014).
Nieh, E.H. et al. Decoding neural circuits that control compulsive sucrose seeking. Cell 160, 528–541 (2015).
Hardaway, J.A., Crowley, N.A., Bulik, C.M. & Kash, T.L. Integrated circuits and molecular components for stress and feeding: implications for eating disorders. Genes Brain Behav. 14, 85–97 (2015).
Aberman, J.E., Ward, S.J. & Salamone, J.D. Effects of dopamine antagonists and accumbens dopamine depletions on time-constrained progressive-ratio performance. Pharmacol. Biochem. Behav. 61, 341–348 (1998).
Hamill, S., Trevitt, J.T., Nowend, K.L., Carlson, B.B. & Salamone, J.D. Nucleus accumbens dopamine depletions and time-constrained progressive ratio performance: effects of different ratio requirements. Pharmacol. Biochem. Behav. 64, 21–27 (1999).
Berridge, K.C., Venier, I.L. & Robinson, T.E. Taste reactivity analysis of 6-hydroxydopamine-induced aphagia: implications for arousal and anhedonia hypotheses of dopamine function. Behav. Neurosci. 103, 36–45 (1989). This study demonstrated that the hedonic reactivity to sucrose is preserved following dopamine depletion.
Shafiei, N., Gray, M., Viau, V. & Floresco, S.B. Acute stress induces selective alterations in cost/benefit decision-making. Neuropsychopharmacology 37, 2194–2209 (2012).
Valentin, V.V., Dickinson, A. & O'Doherty, J.P. Determining the neural substrates of goal-directed learning in the human brain. J. Neurosci. 27, 4019–4026 (2007). This study adapted tasks traditionally used in rodents for human imaging studies of valuation systems.
Dickinson, A. Action and habits: the development of behavioral autonomy. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366, 67–78 (1985). This paper proposed that goal-directed actions could become habits depending on the nature and extent of training. This is now the predominant view of habit formation.
Balleine, B.W. & Dickinson, A. The role of incentive learning in instrumental outcome revaluation by sensory-specific satiety. Anim. Learn. Behav. 26, 46–59 (1998).
Schwabe, L. & Wolf, O.T. Stress prompts habit behavior in humans. J. Neurosci. 29, 7191–7198 (2009).
Schwabe, L. & Wolf, O.T. Socially evaluated cold pressor stress after instrumental learning favors habits over goal-directed action. Psychoneuroendocrinology 35, 977–986 (2010).
Schwabe, L., Tegenthoff, M., Höffken, O. & Wolf, O.T. Concurrent glucocorticoid and noradrenergic activity shifts instrumental behavior from goal-directed to habitual control. J. Neurosci. 30, 8190–8196 (2010).
Schwabe, L., Höffken, O., Tegenthoff, M. & Wolf, O.T. Preventing the stress-induced shift from goal-directed to habit action with a β-adrenergic antagonist. J. Neurosci. 31, 17317–17325 (2011).
Schwabe, L., Tegenthoff, M., Höffken, O. & Wolf, O.T. Simultaneous glucocorticoid and noradrenergic activity disrupts the neural basis of goal-directed action in the human brain. J. Neurosci. 32, 10146–10155 (2012). This paper demonstrated that concurrent activation of glucocorticoid and noradrenergic systems promotes habit formation and disrupts neural activity patterns associated with goal-directed decisions.
Braun, S. & Hauber, W. Acute stressor effects on goal-directed action in rats. Learn. Mem. 20, 700–709 (2013).
Dias-Ferreira, E. et al. Chronic stress causes frontostriatal reorganization and affects decision-making. Science 325, 621–625 (2009). This study found that chronic unpredictable stress causes habit formation in rats and concomitant anatomical alterations in corticostriatal circuits associated with goal-directed versus habitual behavior.
Soares, J.M. et al. Stress-induced changes in human decision-making are reversible. Transl. Psychiatry 2, e131 (2012). This study replicated in humans the findings observed in ref. 87, observing stress-associated habitual behavior and changes in corticostriatal structure and functional activity.
Gourley, S.L. et al. Action control is mediated by prefrontal BDNF and glucocorticoid receptor binding. Proc. Natl. Acad. Sci. USA 109, 20714–20719 (2012). This paper showed that exogenous elevation of stress-related signals promotes habitual behavior, whereas medial prefrontal neurotrophins mediate motivation to seek reward.
Otto, A.R., Raio, C.M., Chiang, A., Phelps, E.A. & Daw, N.D. Working-memory capacity protects model-based learning from stress. Proc. Natl. Acad. Sci. USA 110, 20941–20946 (2013). This paper reported that acute cold-pressor stress in humans selectively disrupted model-based valuation processes that underlie cognitive and deliberative decision-making.
Arnsten, A.F.T. Stress signalling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 10, 410–422 (2009).
Arnsten, A.F.T., Wang, M.J. & Paspalas, C.D. Neuromodulation of thought: flexibilities and vulnerabilities in prefrontal cortical network synapses. Neuron 76, 223–239 (2012).
McEwen, B.S. & Morrison, J.H. The brain on stress: vulnerability and plasticity of the prefrontal cortex over the life course. Neuron 79, 16–29 (2013).
Gamo, N.J. et al. Stress impairs prefrontal cortical function via D1 dopamine receptor interactions with hyperpolarization-activated cyclic nucleotide-gated channels. Biol. Psychiatry (2015).
Kim, J.J., Song, E.Y. & Kosten, T.A. Stress effects in the hippocampus: synaptic plasticity and memory. Stress 9, 1–11 (2006).
Schwabe, L. & Wolf, O.T. Stress and multiple memory systems: from ′thinking′ to ′doing′. Trends Cogn. Sci. (Regul Ed) 17, 60–68 (2013).
Guenzel, F.M., Wolf, O.T. & Schwabe, L. Glucocorticoids boost stimulus-response memory formation in humans. Psychoneuroendocrinology 45, 21–30 (2014).
Thai, C.A., Zhang, Y. & Howland, J.G. Effects of acute restraint stress on set-shifting and reversal learning in male rats. Cogn. Affect. Behav. Neurosci. 13, 164–173 (2013).
Butts, K.A., Floresco, S.B. & Phillips, A.G. Acute stress impairs set-shifting but not reversal learning. Behav. Brain Res. 252, 222–229 (2013).
Acknowledgements
This work was supported by US National Institutes of Health grants P50-MH106428 and F31-DA036278.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article

Cite this article
Hollon, N., Burgeno, L. & Phillips, P. Stress effects on the neural substrates of motivated behavior. Nat Neurosci 18, 1405–1412 (2015). https://doi.org/10.1038/nn.4114
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nn.4114
This article is cited by
-
Chemogenetic activation of corticotropin-releasing factor-expressing neurons in the anterior bed nucleus of the stria terminalis reduces effortful motivation behaviors
Neuropsychopharmacology (2024)
-
Neural sensitivity following stress predicts anhedonia symptoms: a 2-year multi-wave, longitudinal study
Translational Psychiatry (2024)
-
Chronic unpredictable stress induces depression-related behaviors by suppressing AgRP neuron activity
Molecular Psychiatry (2021)
-
Housing conditions during self-administration determine motivation for cocaine in mice following chronic social defeat stress
Psychopharmacology (2021)
-
Responsibility for forgetting
Philosophical Studies (2019)
