
Abstract
Biological competition is widely believed to result in the evolution of selfish preferences. The related concept of the ‘homo economicus’ is at the core of mainstream economics. However, there is also experimental and empirical evidence for other-regarding preferences. Here we present a theory that explains both, self-regarding and other-regarding preferences. Assuming conditions promoting non-cooperative behaviour, we demonstrate that intergenerational migration determines whether evolutionary competition results in a ‘homo economicus’ (showing self-regarding preferences) or a ‘homo socialis’ (having other-regarding preferences). Our model assumes spatially interacting agents playing prisoner's dilemmas, who inherit a trait determining ‘friendliness’, but mutations tend to undermine it. Reproduction is ruled by fitness-based selection without a cultural modification of reproduction rates. Our model calls for a complementary economic theory for ‘networked minds’ (the ‘homo socialis’) and lays the foundations for an evolutionarily grounded theory of other-regarding agents, explaining individually different utility functions as well as conditional cooperation.
Similar content being viewed by others

Introduction
Many societal problems, such as pollution, global warming, overfishing, or tax evasion, result from social dilemmas. In these dilemmas, uniform cooperation would be good for everybody, but each individual can benefit from free-riding1. Although societies found ways to cope with socalled ‘tragedies of the commons’2, the evolution of other-regarding preferences under competitive selection pressure is still a challenging and topical scientific puzzle.
In social dilemma situations, caring about others can reduce individual success. While profit maximisation in single interactions would always demand non-cooperative behaviour, repeated interactions may sometimes support reciprocal altruism and result in human sociality3,4. But even in one-shot interactions, humans are not as selfish as theory suggests. A large body of experimental and field evidence indicates that people genuinely care about each other5,6,7,8. They tend to be not only concerned about individual success, but also about that of others6,8,9.
Also in dictator and ultimatum games, the tendency to share is often attributed to other-regarding preferences10,11. But how did such other-regarding preferences evolve and spread? It was suggested that group selection would solve the puzzle12,13,14 but it only works when groups do not mix15. Therefore, mechanisms not requiring kin or group selection have been looked for16. Some authors argue that humans intrinsically favour fairness10,17. But why do other-regarding preferences then vary geographically6?
To explain an innate sense of fairness, Gintis proposed a modification of reproduction rates by culture18. Other models studying the evolution of fairness preferences typically assume mechanisms in favour of pro-sociality, such as a ‘shadow of the future’4, costly punishment19,20, reputation21,22, genetic favouritism23,24, genetic drift25, or local interactions with an imitation of more successful neighbours26,27,28,29,30. However, the ‘best response’ rule is not favourable for the spreading of cooperation in social dilemma situations, where non-cooperative behaviour creates a higher payoff, no matter what the behavioural strategy of the interaction partner(s) is31.
Results
Model
Our model does not require any of the previously mentioned social mechanisms and it even works when the best response rule is applied. For simplicity, we assume a spatial square lattice with periodic boundary conditions and L × L sites; 60% of the sites are occupied by one agent each, the other 40% are empty. Agents simultaneously interact with all other agents in their Moore neighbourhood—the eight sites surrounding their own site. In each time period, agents can choose to cooperate (C) or to defect (D). For all interactions with neighbours, agents get a payoff. If two interacting agents cooperate, each obtains the amount R (‘Reward’); if both defect, each gets P (‘Punishment’); and if one cooperates and the other one defects, the former one gets S (‘Sucker's Payoff’), while the latter gets T (‘Temptation’). The agents' reproductive fitness in time period t is given by the sum of all payoffs from interactions with neighbours (minus a value of 8|S| to ensure non-negative payoffs and avoid the reproduction of agents who are exploited by all their neighbours).
At the end of each period, individuals die with probability β. To keep population size constant, all agents who die are replaced by an offspring of one of the surviving agents. The likelihood of parents to create an offspring is strictly proportional to their actual payoff, i.e. their reproductive fitness. The offspring is born in one of the empty sites closest to the parent with probability ν (‘local reproduction’), while it occupies a random empty site irrespective of the distance to the parent with probability 1 − ν (‘random reproduction’).
We assume a strict prisoner's dilemma with T > R > P > S. Although collective success is highest when everybody cooperates, defection is the payoff-maximising individual strategy in each single interaction, independently of the neighbours' strategies. In our model, individuals update their strategy (cooperation or defection) based on the myopic best response rule at the end of each period. However, rather than maximising their payoff Pi, we assume here that an individual i chooses the strategy that maximises the utility

where  denotes the average payoff of the interaction partners j. We do this because of previous studies and empirical evidence10,29,32 suggesting that the utility is not just given by the own payoff Pi, but the payoff Pj of interaction partners j is also given a certain weight ρi. The variable ρi ∈ [0, 1]—the ‘friendliness’—represents the degree of other-regarding preferences of agent i. A purely self-regarding individual with ρi = 0 only cares about the own payoff when choosing a strategy. An other-regarding individual gives the own payoff a weight of 1 − ρi and the payoff of interaction partners a weight of ρi. Hence, strategy updates are assumed to be ‘empathic’, but reproduction is exclusively driven by individual payoff.
denotes the average payoff of the interaction partners j. We do this because of previous studies and empirical evidence10,29,32 suggesting that the utility is not just given by the own payoff Pi, but the payoff Pj of interaction partners j is also given a certain weight ρi. The variable ρi ∈ [0, 1]—the ‘friendliness’—represents the degree of other-regarding preferences of agent i. A purely self-regarding individual with ρi = 0 only cares about the own payoff when choosing a strategy. An other-regarding individual gives the own payoff a weight of 1 − ρi and the payoff of interaction partners a weight of ρi. Hence, strategy updates are assumed to be ‘empathic’, but reproduction is exclusively driven by individual payoff.
When selfishness is fixed (ρi = 0), best response behaviour promotes a ‘tragedy of the commons’1. Instead, however, we assume a (genetic or cultural) transmission of friendliness ρi from parent i to offspring j, which is subject to random mutation. In our model, mutation occurs with a constant probability μ that is independent of the strategies pursued in the neighbourhood. To avoid ‘genetic drift’, which would eventually promote friendliness scores of 0.5, the mutation of ρi is specified such that the offspring tends to be more self-regarding than the parent (if ρi > 0.2): With probability 0.8, ρj is set to a uniformly distributed random value between 0 and ρi and with probability 0.2 it is set to a uniformly distributed value between ρi and 1.
Simulation results
Our computer simulations start in the most adverse condition for friendliness and cooperation. At time t = 0, all agents defect and nobody cares about the payoff of others (ρi = 0). However, mutations will eventually create higher levels of friendliness. According to the best response rule, an agent will cooperate, if the utility Ui(C) of cooperation is larger than the utility Ui(D) of defection. The utility of cooperation is

when surrounded by c co-operators and d defectors and the utility of defection is

Therefore, cooperation is expected to occur for

That is, cooperativeness depends on the number of cooperative and defective neighbours, but it also depends on the level of friendliness ρi. We find that, for ρi = 0, agents never cooperate, while above a critical threshold of friendliness, namely for

they cooperate unconditionally. For values ρi of friendliness between (T − R)/(T − S) and (P − S)/(T − S), we find conditional cooperation9, when enough neighbours cooperated in the previous round (note that, in our simulations, S < 0).
Hence, ‘idealists’ with a level of friendliness ρi > (P − S)/(T − S) happen to cooperate even when they are surrounded and exploited by defectors. However, such idealists will normally get miserable payoffs and have very small reproduction rates. They tend to die without reproducing themselves. In fact, other-regarding preferences do not spread and selfishness thrives when offspring occupy randomly selected empty cells.
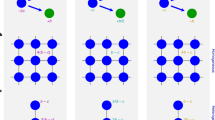
In contrast, when agents reproduce locally, other-regarding preferences suddenly emerge after some time (see Fig. 1A). How does this surprising, sudden transition from the ‘homo economicus’ to the ‘homo socialis’ take place? In principle, mutations could create a random co-location of mutation-borne ‘idealists’ by coincidence after a long time28. This would lead to the formation of a cluster of cooperators of ‘supercritical’ size. Such clustering would dramatically increase the relative fitness of other-regarding agents in the cluster and create sufficiently high reproduction rates to spread friendliness.

A random spatial coincidence of friendly agents can lead to the sudden spreading of other-regarding preferences and a transition from a ‘homo economics’ to a ‘homo socialis’.
The graphs show representative simulation runs on a 30 × 30 spatial grid with periodic boundary conditions. 60% of all sites are occupied with agents who can either cooperate or defect. The payoff of interacting agents is determined as sum of payoffs from prisoner dilemma games with all Moore neighbours. The payoff parameters are: ‘Temptation’ T = 1.1, ‘Reward’ R = 1, ‘Punishment’ P = 0 and ‘Sucker's Payoff’ S = −1. The strategies (cooperation or defection) are updated simultaneously for all agents, applying the myopic best response rule to the utility function of each agent. It weights the payoffs of neighbours with the friendliness ρi and the own payoff with (1 − ρi). Agents die at random with probability β = 0.05. To keep population size constant, surviving agents produce offsprings proportionally to their payoff in the previous round. Offspring move to the closest empty site (ν = 1) and inherit attributes from the parent, here: the friendliness ρi. However, with probability μ = 0.05, the friendliness of offsprings mutates. With probability 0.8 it is ‘reset’ to a uniformly distributed random value between 0 and the friendliness ρi of the parent and with probability 0.2 it takes on a uniformly distributed value between ρi and 1. (A) Average of friendliness and share of cooperating agents as a function of time (one generation is 1/β periods). (B) Average payoffs of cooperators and defectors as a function of time. Initially, defectors are more successful than cooperators. However, when the sudden transition from a ‘homo economics’ to a ‘homo socialis’ occurs, the payoffs for defectors increases, but the payoffs for cooperators increases even more, which implies higher production rates of agents with other-regarding preferences.
However, why does this transition happen in just a few generations (see Fig. 1B), i.e. much faster than expected? This relates to our distinction of preferences and behaviour. When an ‘idealist’ is born in a neighbourhood with friendliness levels supporting conditional cooperation, this can trigger off a cascade of changes from defective to cooperative behaviour. Under such conditions, a single ‘idealist’ may quickly turn a defective neighbourhood into a largely cooperative one. This implies higher payoffs and higher reproduction rates for both, idealists and conditional co-operators.
The intriguing phase transition from self-regarding to other-regarding preferences critically depends on the local reproduction rate (see Fig. 2). The clustering of friendly agents, which promotes other-regarding preferences, is not supported when offspring move away. Then, offspring are more likely to encounter defectors elsewhere and parents are not ‘shielded’ by their own friendly offspring anymore. In contrast, with local reproduction, offspring settle nearby and a clustering of friendly agents is reinforced. Under such conditions, friendliness is evolutionary advantageous over selfishness.

Local reproduction is crucial for the transition from a ‘homo economicus’ to a ‘homo socialis’.
The rate ν of local reproduction determines the probability of an offspring to occupy the closest empty site to the parent. With probability (1 − ν), the offspring moves to an empty site that is randomly selected. All other parameters are specified as in Fig. 1. The circle size indicates average friendliness, while the circle colour represents the share of cooperators. The values are averages over 100 simulation runs between generation 100 and 500. Even at temptation levels around T = 1.3, the above phase diagram shows a sudden transition from self-regarding preferences (small dots) to other-regarding preferences (large circles), when the degree ν of local reproduction is high enough.
Discussion
In conclusion, we offer an over-arching theoretical perspective that could help to overcome the historical controversy in the behavioural sciences between largely incompatible views about human nature. Both, self-regarding and other-regarding types of humans may result from the same evolutionary process. Whereas high levels of intergenerational migration promote the evolution of a ‘homo economicus’, low levels of intergenerational migration promote a ‘homo socialis’, even under ‘Darwinian’ conditions of a survival of the fittest and random mutations. The significance of local reproduction for the evolution of other-regarding preferences is striking and may explain why such preferences are more common in some parts of the world than in others6.
Our modelling approach distinguishes between the evolution of individual preferences and behaviours. This makes cooperation conditional on the level of cooperation in the respective neigh-bourhood. Hence, when a few ‘idealists’ are born, who cooperate unconditionally, this can trigger off cooperation cascades, which can largely accelerate the spreading of cooperation33. Our model can also serve as a basis to develop an economic theory of other-regarding agents. The advantage is that it does not need to assume certain properties of boundedly rational agents—these properties rather result from an evolutionary process. In fact, our model naturally explains the evolution of individually different utility functions, as they are experimentally observed (see Figs. 3 + 4) and also the evolution of conditional cooperators9,34.

Evolution of the distribution of friendliness in the course of time for the parameter values used in Fig. 1.
The plot shows an average over 100 runs, smoothed with MATLAB's local regression using weighted linear least squares and a 1st degree polynomial model. It is clearly visible that a broad distribution of individual utility functions results, even though everybody starts off with a purely self-regarding behaviour, for which the utility function agrees exactly with the payoff function.

Empirical distribution of other-regarding preferences (from R. Murphy et al. 201132, reproduction with kind permission of Ryan Murphy).
(A) This figure shows the primary items from a ‘Slider Measure’ to determine Social Value Orientation. (B) Distribution of Social Value Orientation scores from the experimentally determined Slider Measure, as represented by angles. The dark line is a smoothed kernel density estimation.
A great share of economic literature is based on the assumption of the ‘homo economicus’, who takes decisions without considering the payoff or utility of others. In contrast to this traditional view, the ‘homo socialis’ never takes independent decisions, if the behaviour has external effects35,36. We might characterise this as a situation of ‘networked minds’, where everybody is trying to put himself or herself into other people's shoes, to take into account their utilities in the decision-making process. As a consequence, besides paying attention to networks of companies37, economics should also consider networks of individual minds, i.e. social aspects. This is of particular relevance for information societies, in which individuals are increasingly connected via information and communication systems, such as social media38,39. A theory of networked minds could make a significant contribution to the convergence of the behavioural sciences41 and it might also shed new light on social capital, power, reputation and value and create a fundamentally new understanding of these40. We believe that this view can stimulate a huge and exciting field of research and lead to a complementary theory to the one based on the ‘homo economicus’. Due to the simplicity and fundamental character of the model proposed by us, we expect that it might serve as a starting point and basis for this new field.
References
Hardin, G. The tragedy of the commons. Science 162, 1243–1248 (1968).
Ostrom, E. Governing the Commons: The Evolution of Institutions for Collective Action (Cambridge University Press, Cambridge, 1990).
Trivers, R. L. The evolution of reciprocal altruism. Q. Rev. Biol. 46, 35–57 (1971).
Axelrod, R. & Hamilton, W. D. The evolution of cooperation. Science 211, 1390–1396 (1981).
Kahneman, D., Slovic, P. & Tversky, A. Judgement Under Uncertainty-Heuristics and Biases (Cambridge University Press, Cambridge, 1982).
Henrich, J. et al. In search of homo economicus: behavioral experiments in 15 small-scale societies. Am. Econom. Rev. 91, 73–78. (2001).
Camerer, C. Behavioral Game Theory—Experiments in Strategic Interaction (Princeton University Press, Princeton, 2003).
Hoffman, E., McCabe, K. & Smith, V. Social distance and other-regarding behavior in dictator games. Am. Econom. Rev. 86, 653–660 (1996).
Fehr, E. & Fischbacher, U. The nature of human altruism. Nature 425, 785–791 (2003).
Tricomi, E., Rangel, A., Camerer, C. & O'Doherty, J. Neural evidence for inequality-averse social preferences. Nature 463, 1089–1091 (2010).
Dufwenberg, M., Heidhues, P., Kirchsteiger, G., Riedel, F. & Sobel, J. Other-regarding preferences in general equilibrium. Review of Economic Studies 78, 613–639 (2011).
Bowles, S. Group competition, reproductive levelling and the evolution of human altruism. Science 314, 1569–1572 (2006).
Traulsen, A. & Nowak, M. A. Evolution of cooperation by multilevel selection. PNAS 103, 10952–10955 (2006).
Perc, M., Gómes-Gardeñes, J., Szolnoki, A., Floría, L. M. & Moreno, Y. Evolutionary dynamics of group interactions on structured populations: A review. J. R. Soc. Interface 10, 20120997 (2013).
Sober, E. & Wilson, D. S. Unto Others: The Evolution and Psychology of Unselfish Behavior (Cambridge University Press, Cambridge, 1998).
Akay, E., Van Cleve, J., Feldman, M. W. & Roughgarden, J. A theory for the evolution of other-regard integrating proximate and ultimate perspectives. PNAS 106, 19061–19066 (2009).
Fehr, E. & Schmidt, K. A theory of fairness, competition and cooperation. Q. J. Econ. 114, 817–868 (1999).
Gintis, H. Strong reciprocity and human sociality. J. Theor. Biol. 206, 169–179 (2000).
Fehr, E. & Gächter, S. Altruistic punishment in humans. Nature 415, 137–140 (2002).
Helbing, D., Szolnoki, A., Perc, M. & Szabó, G. Evolutionary establishment of moral and double moral standards through spatial interactions. PLoS Comput. Biol. 6 (4), e1000758 (2010).
Nowak, M. A. & Sigmund, K. Evolution of indirect reciprocity by image scoring. Nature 393, 573–577 (1998).
Milinski, M., Semmann, D. & Krambeck, H. J. Reputation helps solve the tragedy of the commons. Nature 415, 424–426 (2002).
Simon, H. A. Altruism and economics. Am. Econom. Rev. 83, 156–161 (1993).
Wilson, D. S. & Dugatkin, L. A. Group selection and assortative interactions. Am. Nat. 149, 336–351 (1997).
Leimar, O. & Hammerstein, P. Evolution of cooperation through indirect reciprocity. Proc. R. Soc. Lond. B 268, 745–753 (2001).
Nowak, M. A. & May, R. M. Evolutionary games and spatial chaos. Nature 359, 826–829 (1992).
Epstein, J. M. Zones of cooperation in demographic prisoner's dilemma. Complexity 4 (2), 36–48 (1998).
Helbing, D. & Yu, W. The outbreak of cooperation among success-driven individuals under noisy conditions. PNAS 106 (8), 3680–3685 (2009).
Szabó, G. & Szolnoki, A. Selfishness, fraternity and other-regarding preference in spatial evolutionary games. J. Theor. Biol. 299, 81–87 (2011).
Perc, M. & Szolnoki, A. Coevolutionary games – A mini review. BioSystems 99, 109–125 (2010).
Roca, C., Cuesta, J. A. & Sanchez, A. Evolutionary game theory: temporal and spatial effects beyond replicator dynamics. Phys. Life Rev. 6 (4), 208–249 (2009).
Murphy, R. O., Ackermann, K. A. & Handgraaf, M. J. J. Measuring social value orientation. Judgment and Decision Making 6 (8), 771–781 (2011).
Fowler, J. H. & Christakis, N. Cooperative behavior cascades in human social networks. PNAS 107, 5334–5338 (2010).
Gracia-Lazaro, C. et al. Heterogeneous networks do not promote cooperation when humans play a Prisoner's Dilemma. PNAS 109, 12922–12926 (2012).
Christakis, N. A. & Fowler, J. H. Connected (Back Bay Books, Norwell, MA, 2011).
Bolton, G. E. & Ockenfels, A. ERC: A theory of equity, reciprocity and competition. The American Economic Review 90 (1), 166–193 (2000).
Schweitzer, F. et al. Economic networks: The new challenge. Science 325, 422–425 (2009).
Helbing, D. & Carbone, A. (eds.) Participatory science and computing for our complex world. EPJ Special Topics 214, 1–666 (2012).
Helbing, D. Globally networked risks and how to respond. Nature, in print (2013).
Helbing, D. & Balietti, S. Fundamental and real-world challenges in economics. Science and Culture 76 (9–10), 399–417 (2010).
Gintis, H. The Bounds of Reason: Game Theory and the Unification of the Behavioral Sciences (Princeton University Press, Princeton, 2009).
Acknowledgements
We would like to thank Thomas Chadefaux, Michael Mäs and Ryan Murphy for valuable comments. DH acknowledges support by the Future and Emerging Technologies program FP7-COSI-ICT of the European Commission through the project QLectives (Grant No. 231200) and by the ERC Advanced Investigator Grant ‘Momentum’ (Grant No. 324247). DH is grateful to Stefan Rustler for preparing Fig. 3 and to Ryan Murphy for providing Fig. 4.
Author information
Authors and Affiliations
Contributions
D.H. and T.G. developed the model and wrote the paper. T.G. and C.W. performed the computer simulations and the data analysis.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Grund, T., Waloszek, C. & Helbing, D. How Natural Selection Can Create Both Self- and Other-Regarding Preferences and Networked Minds. Sci Rep 3, 1480 (2013). https://doi.org/10.1038/srep01480
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep01480
This article is cited by
-
On the role of collective sensing and evolution in group formation
Swarm Intelligence (2018)
-
Multiple effect of social influence on cooperation in interdependent network games
Scientific Reports (2015)
-
Globally networked risks and how to respond
Nature (2013)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
