
Abstract
Much of what we know about terrestrial life during the Carboniferous Period comes from Middle Pennsylvanian (~315–307 Mya) Coal Measures deposited in low-lying wetland environments1,2,3,4,5. We know relatively little about terrestrial ecosystems from the Early Pennsylvanian, which was a critical interval for the diversification of insects, arachnids, tetrapods, and seed plants6,7,8,9,10. Here we report a diverse Early Pennsylvanian trace and body fossil Lagerstätte (~320–318 Mya) from the Wamsutta Formation of eastern North America, distinct from coal-bearing deposits, preserved in clastic substrates within basin margin conglomerates. The exceptionally preserved trace fossils and body fossils document a range of vertebrates, invertebrates and plant taxa (n = 131), with 83 distinct foliage morphotypes. Plant-insect interactions include what may be the earliest evidence of insect oviposition. This site expands our knowledge of early terrestrial ecosystems and organismal interactions and provides ground truth for future phylogenetic reconstructions of key plant, arthropod, and vertebrate groups.
Similar content being viewed by others

Introduction
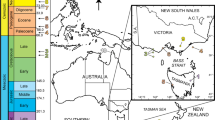
Modern terrestrial ecosystems are defined not only by the composition of their communities but also by functional interactions among them. These relationships and their diversity play important roles in the assembly and maintenance of terrestrial ecosystems today, but their development through time is challenging to infer from the fossil record (e.g.,11,12) without instances of exceptional preservation (i.e., Lagerstätten). The Carboniferous Coal Measures of Europe and North America host a comprehensive record of Middle Pennsylvanian (~315–307 Mya) life on land1,2. Lagerstätten from these deposits occur as body fossils in shales and in nodules embedded among economically valuable coal seams that have been mined for centuries. Most of these Lagerstätten, such as Mazon Creek (Illinois, USA), Union Chapel Mine (Alabama, USA), Hamilton Quarry (Kansas, USA), Linton Mines (Ohio, USA), and Montceau-les-Mines (France), are dated to the Moscovian–Kasimovian stages (= Bolsovian–Stephanian; ca. 315–304 Ma)3,4,5 and represent marginal marine and lowland tropical continental settings (Fig. 1).

Palaeogeography of the Wamsutta Formation and Comparisons of Carboniferous Terrestrial Lagerstätten (A) Modern geographical location of the Wamsutta Formation. Base map obtained from www.freeworldmaps.net. B Palaeogeographic map (~317 Mya) of important Pennsylvanian terrestrial Lagerstätten placed in palaeogeographic context. Map used with permission © 2023 Colorado Plateau Geosystems Inc.19. Wam = Wamsutta Formation; Jo = Joggins Fossil Cliffs; Bi = Bickershaw Formation; Uc = Union Chapel Mine; Li = Linton Mines; Mc = Mazon Creek; Mo = Montceau-les-Mines; Hq = Hamilton Quarry, Eqr= Equator. C A comparison of the biota from these Lagerstätten. Note that for the Linton Diamond mines, plant material has been recovered but has yet to be reported in peer-reviewed literature. References used to compile this table are presented in Supplementary Note 2. Lepospondyl image courtesy of Gareth Monger, Temnospondyl by Scott Harman, Apterygotes by Birgit Lang, and Lepidodendrids by Eliana P. Coturel. No changes to original images were made. These organismal cliparts are under CC BY 3.0 license (see https://creativecommons.org/licences/by/3.0/).
Lagerstätten that sample terrestrial ecosystems from Bashkirian times, prior to the rapid accumulation of the Coal Measures peat substrates13, are uncommon. Examples of Bashkirian Lagerstätten include the Joggins Formation (Nova Scotia, Canada; e.g.,7) and the Bickershaw Formation (Lancashire, UK8,9), but these, too, are representative of basinal axes and humid lowlands. Palaeozoic Lagerstätten from clastic-dominated, dynamic environments along basin margins are extremely rare and can provide information about biotic and abiotic conditions that prevailed in ecosystems not sampled in the historical exploitation of the Coal Measures. Palaeontological data from this earlier, drier time in the Pennsylvanian record the daily habits of tetrapod, arthropod, and seed plant groups at a critical time in their diversification and expansion into a wide range of terrestrial environments (e.g.,6) that were changing rapidly due to expansion and contraction of Gondwanan ice sheets during the Late Palaeozoic Ice Age (LPIA).
In this work, we report on fossil discoveries from the Lantern North (LN) site in the Wamsutta Fm., an Early Pennsylvanian (~320–318 Mya) Lagersttäte from an intramontane setting near the axis of the Acadian highlands (Fig. 1B). The Wamsutta Formation of Massachusetts, USA (Fig. 1A; Supplementary Figs. 1 and 2) differs from typical Pennsylvanian coal swamp deposits in that it consists exclusively of clastic sediments that, by measure of thickness, are coarse-grained, highly oxidised redbeds deposited against the basin’s structural margin (see Supplementary Fig. 2). The diverse fossils at LN occur mainly as trace fossils, (an ichnofossil-Lagerstätte14) and plant impressions, recording detailed morphological features of organisms not typically available (e.g., full-body impressions) as well as interactions between organisms. The sheer abundance of in situ trace fossils at this site, in addition to plant impressions and body fossils, allow for palaeoecological inferences that traditional body fossil Lagerstätten seldom provide.
Age and geology of the Lantern North site, Wamsutta formation, Massachusetts, USA
The geologic and tectonic history of the New England Carboniferous basins adjacent to the Euramerican Variscan-Appalachian orogen has been summarised by Skehan et al.15,16, Mosher17, Murray et al.18, and references therein (see also Supplementary Note 1). While our understanding of the palaeogeographic context of these basins has improved over decades of enquiry, details regarding the fossils preserved in these rocks and their significance with regard to palaeotropical terrestrial ecosystem development remain understudied. Both the Narragansett and Norfolk basins are the southernmost of a series of apparently intramontane transtensional rifts17 situated in the overlap of Acadian (Devonian) and Alleghanian-Variscan (Carboniferous) deformation17 with their earliest structural origins in the Late Devonian17,19. During mid- to late Carboniferous times, the basins were filled with sediments delivered via terrestrial depositional systems shed from the highlands during the formation of Pangea. Plate tectonic reconstructions20 place Massachusetts basins within subtropical to tropical palaeolatitudes, seemingly isolated from the larger, coal-rich midwestern and Appalachian basins of the U.S., as well as the Canadian Maritimes (Fig. 1B).
Thousands of metres of interbedded conglomerate, sandstone, and shale fill these basins, most of which were segmented and offset by post-depositional faulting and folding18,21, making contiguous stratigraphic sections rare. Despite these difficulties, some key age relationships have been established. The Carboniferous strata lie upon a Late Devonian unconformity22 in the area studied. The basal Carboniferous sediments in both basins contain Pennsylvanian plant fossil assemblages23,24,25,26. Some ambiguity has existed as to the age of the Wamsutta Fm. because of the relative paucity of plant fossils in this coarser-grained and highly oxidised succession compared with the overlying shale-dominated, carbon-rich Rhode Island Fm. Recent efforts by Lyons and Sproule26 refined the age of the Wamsutta Fm. to Langsettian (317–315 Mya, Late Bashkirian), but these were based upon considerably less complete assemblages compared with those of the present study. Our collections from the Wamsutta Fm. in the LN field area greatly expand the list of known floral morphotypes (see Plants) that support a Bashkirian age of ~320–318 Mya, slightly older than previous estimates.
Of critical importance to its palaeoecological value, the Wamsutta Fm. has been interpreted as representing deposition on seasonally wet, possibly forested, low-gradient alluvial fans17,27,28. Conglomeratic sheets with imbricated cobbles likely represent large distributary channel bars that were activated during high-flow regimes. Subordinate cross-bedded sandstone, erosional surfaces, and mass movement deposits in the representative section indicate a dynamic, rapidly changing environment. Sedimentological evidence, as well as the position of the Wamsutta Fm. against the western bounding faults and its limited areal extent, lends support to the alluvial fan interpretation, possibly its medial to distal portions. The fossiliferous red shale facies may, therefore, represent deposition in shallow depressions near the base of a fan complex that hosted ephemeral or seasonal ponds and pools in which suspended sediment could settle after a flood. The clay minerals provided by colluvium in the contributing catchment were likely produced by the weathering of felsic rocks, abundant in the local area. Cohesive properties of the clay-rich layers, in turn, preserved impressions in fine detail. Raindrop impressions and occasional mud cracks on bedding plane surfaces associated with the plant and trace fossils indicate subaerial exposure after deposition and may be related to loss of water due to infiltration on a permeable fan deposit or to evaporation. In contrast with the overlying Rhode Island Formation, a carbonaceous shale- and sandstone-dominated fluvial lowland succession in the centre of the basin containing discontinuous coal seams, the Wamsutta Fm. clearly represents the clastic alluvial component of the Narragansett Basin from earlier in its Carboniferous history.
Results and discussion
Wamsutta biota
Amphibians, eureptiles, and non-mammalian synapsids
The Narragansett Basin lies at what is thought to have been the palaeoequatorial centre of terrestrial vertebrate radiations during the early Pennsylvanian Period29,30,31,32. Given that the skeletal record from this critical interval is poor, analysis of the tetrapod trace fossil record assumes paramount importance. Fortunately, work that quantitatively links Permo-Carboniferous track and trackway characteristics to osteological features of certain clades has progressed in recent years33,34,35, and the present study refers to these works in making our palaeobiological interpretations of the Wamsutta vertebrate track suite.
Discrete tracks and track pairs (manus/pes) are the most commonly recovered trace fossils due to the present difficulty of exposing wide bedding planes that would reveal complete trackways. Individual track lengths range from ~2 to ~170 mm (Fig. 2A–J), most commonly small tetrapod trackways with and without tail drag marks. Based on both tracks and trackways, at least three major tetrapod groups were present at the time of Wamsutta deposition, as represented by at least six ichnogenera. The total vertebrate track assemblage from the Wamsutta Formation represents a complex palaeoecosystem that hosted brachiosaur, cotylosaur and early amniote groups, all preserved alongside a diverse suite of invertebrate trace fossils and plant impressions.

Vertebrate Ichnotaxa from the Wamsutta Formation (A) Specimen MCZ:IP:197442 shows two plantigrade pedes in a straight, parallel orientation, with relatively short, equal length digits on each. B MCZ:IP:197441 (C) MCZ:IP:193461 (D) MCZ:IP:200396 (E) MCZ:IP:193669 (F) MCZ:IP:200794 (G) MCZ:IP:200766 (H) MCZ:IP:193466 (I) MCZ:IP:193480 (J) MCZ:IP:197180 (K) MCZ:IP:200766 (L) MCZ:IP:200138 (M) Specimen MCZ:IP:197349 showing multiple sinusoidal Undichna trails of different sizes crossing a muddy substrate. N A single, sinusoidal Undichna trail; MCZ:IP:197448 (O) Specimen MCZ:IP:200765 with sets of four to five curved striations ending in digit prints, indicating a swimming tetrapod that scratched the substrate. Line drawings of tracks modified from Buchwitz et al.35. Scale bars are 1 cm unless otherwise stated.
The most frequent trackways are those of temnospondyl amphibians (Fig. 2D, I, J, K), common and expected components of early Late Carboniferous tropical ecosystems. These tracks and trackways are characterised by a four-digit manus with distinct distal toe pads and a five-digit pes with high divarication that is wider and has longer digits. The presence of larger, herbivorous diadectomorphs (Fig. 2C) and carnivorous non-mammalian (pelycosaur-grade) synapsids (see Fig. 2B, G, L) are indicated by tracks over 100 mm in total length, but complete trackways with gait patterns have yet to be uncovered. Diadectomorphs (Ichniotherium) were an important group of early terrestrial herbivores, and these large primary consumers possibly represented prey of the large-bodied pelycosaur-grade synapsids. The Lantern North site in the Wamsutta Formation is interpreted to be somewhat older than the well-known Joggins Formation (Canada, Nova Scotia; ~315 Ma), Bochum Formation (Germany; ~315 Ma33;), and late Carboniferous vertebrate localities, making the occurrence of Ichniotherium and Dimetropus at the Wamsutta site the oldest for these important groups. Rare pentadactyl manus/pes pairs with long, thin digits reveal the presence of non-synapsid amniotes, as well, indicating that stem reptiles or true reptiles were living and diversifying in xeromorphic settings, in addition to more humid tropical ecotopes like those represented by the Joggins site.
Other Vertebrates and Behaviours
Certain laminated mudstone units of the Wamsutta Formation preserve multiple trace fossils that represent swimming vertebrate animals recorded by sinusoidal grooves (Undichna isp.) made in substrates at the bottom of ephemeral pools and ponds (Fig. 2M, N). All trails are diminutive, ranging from 13/24 mm (amplitude/wavelength) down to 3.6/7.2 mm. Multiple types of trails, differentiated by the number of overlapping grooves in each trail, are present on one slab (Fig. 2M), exhibiting different behaviours and potentially revealing varying ontogenetic stages of the same animal. Associated with the trail-bearing surface are sets of curved, parallel linear impressions that may be related to plant foliar material deposited after the trails were made, potentially enhancing their preservation. The trace fossil Undichna is understood to have been produced by a fish dragging one or more fins through the substrate. If the fossil is representative of fish locomotion, the ephemeral pools and ponds may have had occasional connections to more permanent waterways that served to support these populations. Associated undulatory striations suggest a maker that bore limbs with digits that were dragged across the substrate while swimming (Fig. 2O). Given the abundance of temnospondyl amphibian trackways in subaerial facies, these traces with their distinctive features are likely to have been made by swimming tetrapods.
Invertebrates
Insecta:Pterygota (winged insects)
The oldest trace fossil unequivocally produced by a winged insect, a full-body impression of a stem-group ephemeropteran (mayfly; Fig. 3A), was described from the Wamsutta Formation in 201136. It was not assigned a binomen at the time because of the rarity of such fossils (fewer than ten specimens of Palaeozoic pterygote full-body impressions have been recorded worldwide37), limiting confidence in the erection of a taxon based on a single specimen. However, since the discovery of this stem empheropteran, fourteen distinct full-body impressions of pterygote insects have been recovered from the Wamsutta Formation ranging in size from ~ 1.0 to 4.0 cm in length (e.g., Figure 3A–F) representing nymphal (Fig. 3E) to adult (Fig. 3A–D, F) forms. These remains are among the earliest examples of pterygote insect anatomies, as only a handful of insect ‘body’ fossils have been recovered from the early-mid Bashkirian, the vast majority consisting only of wings. While wing impression fossils are found at the Lantern North site, the full-body impressions are not only among the earliest records of insect bodies in the fossil record but also among the earliest examples of recorded behaviour directly attributable to this important group. Pterygote full-body impressions from the Wamsutta were made while the tracemakers were alive and in the case where movement cannot be discerned, characters such as “living posture” as defined in Knecht et al.38 distinguish these fossils from body fossils of deceased animals. This unique preservational style provides a rare window into the behaviour and anatomy of Palaeozoic insects otherwise represented solely by wings.

Invertebrate full-body impressions (FBI), body fossils, tracks and trails (A) unnamed ephemeropteran FBI (SEMC-F97) (B) (MCZ:IP:197443) (C) (MCZ:IP:197109) (D) (MCZ:IP:197415) (E) (SEMC-ATL-SM-6-34) (F) palaeodictyopteran FBI (MCZ:IP:197429) (G) Tonganoxichnus isp. (SEMC-F1028) (H) (MCZ:IP:197424) (I) Tonganoxichnus attleboroensis, holotype (SEMC-F1025) (J) Myriapod FBI (MCZ:IP:197417) K Inmontibusichnus charleshenryturneri, holotype (MCZ:IP:198045) (L) ricinuleid sp., (MCZ:IP:197444) (M) xiphosuran prosoma body fossil with pustulose ornamentation (SEMC:ATL-SM-2-115) (N) (MCZ:IP:197183) Natural cast of enroled xiphosuran embedded in substrate in a defensive posture, with meandering burrow leading to impression. Note segmentation on opisthosoma and rounded outline of prosoma superposed. Scale bars = 1 cm, unless stated otherwise.
Insecta:Apterygotes (Archaeognathans)
Distinguishing immature pterygotes from apterygotes can be problematic in some cases but here the presence of detailed anatomy and jumping behaviours visible in the traces, in addition to replication in multiple prior neoichnological studies involving apterygotes39,40 allow for this distinction. Fossil evidence for apterygote insects is abundant in the fine-grained facies of the Wamsutta Formation. Benner et al.41 described the new ichnospecies Tonganoxichnus attleboroensis (Fig. 3I), a full-body impression of an archaeognathan insect from Lantern North. They further revised the ichnogenus in order to accommodate a repeatedly-produced major behavioural variant, named Cardinichnus, based on abundant evidence for insect activity in the red beds. Clear details of the ventral portions of the bodies are recorded, including tergites, abdominal styli, terminal filaments, and anterior palpi, linking the trace fossils to archaeognathan tracemakers (Fig. 3H–G). The traces include full body impressions of small archaeognathans and their appendages (Fig. 3G, G1–G2) in repeated, linear, and semi-circular patterns as they performed daily locomotory and feeding behaviours on moist substrates40,41,42,43. When first recognised in the fossil record, Tonganoxichnus was predicted to appear primarily in tidally-influenced marginal marine facies with high levels of available moisture and decaying vegetation43,44, but their discovery in such abundance, albeit with diminutive individuals (Fig. 3G, G1–G2), in the clastic, proximal environments of the Wamsutta Formation reveals a broader environmental distribution39. Archaeognathans, which today are critical components of terrestrial detritivore ecospace, were major components of pre-Pennsylvanian terrestrial ecosystems45,46, prior to intense competition by terrestrial herbivores. As new forms of leaf-bearing plants evolved, competition may have pushed archaeognathans further into the detritus to seek out decaying vegetation and organics47. Their presence at LN is an indication of their adaptation to and success in the terrestrial detritivore role by the Pennsylvanian Period, as well as their close association with plant communities that were growing increasingly diverse and occupying new habitats.
Arachnida:Thelyphonida (Whip Scorpions)
Whip scorpions (Uropygi: Thelyphonida) are extremely rare in the fossils of Coal Measures. Only nine taxa (eight body fossils and one trace fossil) have been described, including two from the Narragansett Basin38. Inmontibusichnus charleshenryturneri (Fig. 3K), recovered from the Wamsutta Formation, is a full-body impression (trace fossil) of a whip scorpion preserved at the end of a partial trackway. To our knowledge, this is the first record of its kind that can be unquestionably attributed to a whip scorpion tracemaker. With a body length of < 10 mm, Inmontibusichnus charleshenryturneri is the smallest known Palaeozoic whip scorpion, although it is unclear whether it represents an adult or juvenile. Most Coal Measures whip scorpions come from England, Germany and the Czech Republic; the Narragansett Basin is only the second site in the Western Hemisphere to have Palaeozoic thelyphonids, filling an important biogeographical gap in the Euramerican distribution of Carboniferous whip scorpions between the westernmost site (Mazon Creek, Illinois, USA) and the easternmost sites in Europe38.
Arachnida:Ricinulei (Hooded Tick-spiders)
Ricinuleids are enigmatic arachnids, found today only in tropical forests in South and Central America, the Caribbean and West Africa, as well as in caves in Texas48. Palaeozoic fossil ricinuleids are known only from the Carboniferous Coal Measures of Europe and North America49. Specifically, all North American fossil ricinuleids come from Illinois and the Mazon Creek Lagerstätte, with the exception of a single specimen found in Oklahoma50. Thus, fossil ricinuleids in the Wamsutta Formation, which are coeval with the oldest ricinuleids known anywhere51, represent an important stratigraphic and geographic locality for a group with a sparse known distribution52. The new Wamsutta specimens are two individual traces oriented at right angles to one another on the same slab. The main specimen, a body fossil, is preserved dorsoventrally to the bedding, and shows the prosoma, with its distinctive cucullus, abdomen, and some legs (Fig. 3L). The other specimen, a full-body impression (trace fossil) appears to be lying on its side and is only vaguely expressed. It is possible that the second specimen represents a prod mark of the first, from when it initially fell onto the substrate, or it could represent the spinoff of a newly moulted individual. At ~11.0 mm body length (including the cucullus) the specimen is similar in size to other Pennsylvanian ricinuleids, which are, in general, far larger than their extant counterparts52. The size, fragility, and secretive behaviours of this group commonly preclude them from being fossilised. The Wamsutta ricinuleids are the first fossil ricinuleids, to our knowledge, to be found outside of amber or carbonate nodules52,53, both modes of exceptional preservation. Additionally, as a result of trace fossils made while the animal was alive, the Wamsutta specimens are the first fossil ricinuleids to our knowledge that have been found in situ, providing unequivocal evidence of their habitat and behaviour.
Myriapods (Millipedes and Centipedes)
Myriapoda are a well known constituent of Carboniferous terrestrial ecologies and the Wamsutta Formation, they are primarily represented as repichnia, trace fossils created as a result of locomotion. This includes the ichnogenera Diplichnites (Fig. 3O), Diplopodichnus, and Protichnites all which can be reasonably attributed to myriapods and reflect a diversity of behaviours, sediment saturation, and tracemaker bauplans. Dozens of trackways have been recovered, ranging in size from 2–25 mm with an average of 5 mm. One slab in particular, shows multiple Protichnites of similar widths that appear to be emerging from a central depression in the substrate, possibly representing communal living or a hatching event. In addition to trackways, partial full-body impressions made by living myriapods have been recovered (Fig. 3N) as well as body fossils of large (~27 mm wide) disarticulated myriapods and smaller (~10 mm wide) articulated but partial myriapod bodies.
Xiphosura (Horseshoe crabs)
Xiphosuran body impressions and trails are present within the formation, likely indicating at least ephemeral bodies of water. Some workers have suggested that certain Xiphosuran groups were capable of living among the leaf litter and dense vegetation of humid environments54. A hypichnial, full-body impression (prosoma, thoracetron, and telson) of one Xiphosuran specimen preserves details of the dorsal anatomy in what appears to be a defensive or protective posture (Fig. 3N). This specimen measures ~2.5 cm in length and is preceded by a burrowing trail made by the same individual. Another specimen preserves the dorsal view of a prosoma with pustulose cuticular ornamentation densely distributed across the surface (Fig. 3M). This feature is unusual within Xiphosurans and is only shared with one other species, Albalimulus bottoni, the oldest known limulid which comes from the lower Carboniferous of Scotland55. Some other arthropod groups possess this diagnostic character (e.g., arachnids, chasmataspidids, eurypterids, aglaspids, cheloniellids; (55 and references within); its presence in these early taxa suggests that this may have been an ancestral trait.
Invertebrate (worm and larval) tracks and trails
Epichnial invertebrate tracks and trails are abundant in the Wamsutta, representing resting (Cubichnia), locomotory (Repichnia), grazing (Pascichnia), and various feeding traces. Example ichnogenera include: Acanthorhaphe, Cochlichnus, Didymaulichnus, Diplichnites, Fustiglyphus, Gordia, Helminthoidichnites, Helminthopsis, Planolites, Protichnites, Octopodichnus, and Treptichnus. These traces are generally attributed to myriapods, arachnids, and larval insects based on previous morphological and behavioural studies. Endichnial traces are extremely rare and represented solely by the dwelling (Domichnia) ichnogenera Skolithos.
Plants
The first concerted attempt to describe the floral assemblage of the Wamsutta Fm. took place in 194423. In that study, 12 species were recovered and the relative abundance of Calamites and Cordaites in the total population was noted. Unfortunately, specimens from this work can no longer be located in any collection and are considered lost. Figures in the paper could not be used to validate the conclusions, and because of this, Lyons and Sproule26 conducted a new survey of the Wamsutta flora based on Sproule’s collections, identifying 22 species. Lyons and Sproule26 reported that their Wamsutta Fm. macroflora suggested a Late Bashkirian age of ~317–315 Ma. However, specimens from this study were also not reposited, and their whereabouts are unknown.
An updated LN floral assemblage study was conducted using fossils collected after 2007. Eighty-three distinct foliage morphotypes preserved as detailed impressions have been catalogued from the site. Fossilised stems, roots, seeds, and other reproductive structures bring the total to 131 taxa, a nearly six-fold increase in identified organ taxa, considered diverse among palaeofloras of any age (see Supplementary Fig. 3 for relative percentages of taxa). Our long-term sampling and larger dataset of Wamsutta fossil flora provides an age estimate of ~320–318 Ma; slightly older than that of Lyons and Sproule26. Among other things, preserved sphenopsid stems show longitudinal ribbing that is only partly offset across nodes, suggesting they belong to Mesocalamites, most characteristic of early Baskirian or even Serpukhovian vegetation.
Xeromorphic and mesomorphic-to-hygromorphic components are well presented in the assemblage (Fig. 4). Xeromorphic plants are represented by a substantial presence of cordaitaleans, along with possibly allochthonous remains of plants of uncertain affinities. Meanwhile, the mesomorphic-to-hygromorphic group encompasses sphenopsids, lycopsids, ferns and medullosalan pteridosperms, among other taxa56. Notably, xeromorphic and mesomorphic-to-hygromorphic groups occur on the same horizons but with substantial heterogeneity in their relative proportions and distributions, potentially driven by strong lateral variations in environmental conditions typical of rapidly-shifting, well-drained environments associated with proximal clastic environments.

Macrofloral fossils from the Wamsutta Formation (A) (?)Mesocalamites sp. (MCZ:IP:200252) (B) Cordaites sp. (MCZ:IP:200113) (C) Palaeostachya sp. (MCZ:IP:200056) (D) Neuropteris obliqua (MCZ:IP:200069) (E) Rhacopteridium sp. (MCZ:IP:200182) (F) Stigmaria sp. (MCZ:IP:193474) (G) Oligocarpia sp. (MCZ:IP:197372) (H) seed (Carpolithus sp.) associated with Cordaites (MCZ:IP:200057). All scale bars = 1 cm.
Such successions have historically been considered upland deposits, but in recent years, changes in plant communities like these have increasingly been interpreted as driven by climate oscillations rather than exclusively or mainly by elevation, as the term upland would imply57,58. The Wamsutta flora is dominated by Cordaites, a trend which, in resonance with observed physical sedimentary characteristics, has been interpreted to indicate sub-humid to dryland assemblages that may have been seasonal57.
Spores and palynomorphs
Maceration of Wamsutta material was performed in an attempt to extract fossil palynological content and spore samples, but was unsuccessful due to a combination of high thermal maturity and high levels of oxidation. Impressions of megaspores are preserved at LN, however, in both positive and negative relief. Size ranges fall into three generalised groupings based on diameter; < 0.5 mm (n = 26), 0.5 mm (n = 9), > 0.5 mm (n = 5). Spore impressions with the greatest detail occur in positive relief with diameters at or above 0.5 mm and contain details of possible trilete marks.
Organismal Interactions
Plant-fungi associations
Plant-fungal associations are well known within the modern literature59,60,61,62 with many examples of how fungi, particularly mycorrhizal fungi, form mutualisms with land plants63,64,65. Fungal rhizomorphs, consisting of large cords of fungal hyphae, belong to the Basidiomycota, a group which mainly consists of decomposers and ectomycorrhizal fungi. They act as pathways for transporting water, nutrients, and carbon into plants, as documented extensively for modern forests66. Within the Wamsutta Formation, there is evidence to support the presence of fungal rhizomorphs within the fossil landscape. Unlike previous studies which have documented microscopic rhizomorphs67,68, the clastic, proximal, and non-water inundated environment of the LN site preserves macroscopic evidence of cord forming fungi. Most notable are rhizomorphs associated with abundant Cordaites sp. leaves, with one particular example consisting of fungal nodes arising from a leaf and running across the sediment (Figs. 5E, F, S5A, S5B). As leaf compression fossils within this site likely represent the original fossilised forest floor, this setting may suggest that the rhizomorphs were working to decompose the leaf litter, acquiring, transporting, and supplying nutrients to the host-plant.

A A gall preserved on Sigillaria sp. (MCZ:IP:197357); B shows close up of gall seen in (A); C an oviposition lesion preserved on Cordaites sp. (MCZ:IP:197328); D piercing and sucking punctures on a Cordaites sp. (MCZ:IP:197395); E and F show rhizomorphs on Cordaites sp. (MCZ:IP:197310). E shows rhizomorphs originating from the leaf surface to the matrix as a branched network, typical of modern forms (also see Fig. S5). White scale bars denote 1 cm unless otherwise noted in yellow.
Plant-insect associations
Insect feeding recorded as damage types (DTs) represent distinct patterns of insect herbivory characterised by differences in the shape, size, position, and associated morphological features involved in feeding (69 and subsequent addenda). These distinct patterns can be further grouped into larger categories called functional feeding groups (FFGs69; and subsequent addenda) which represent major differences in mouthpart structure and feeding ecologies. Three well-preserved FFGs have been found within the diverse host-plant assemblage of the Wamsutta Formation: oviposition (Fig. 5C), piercing and sucking (Fig. 5D), and galling (Fig. 5A, B). It should be noted that, although oviposition does not represent a feeding interaction per se between a host-plant and its insect herbivore, its presence indicates a different, yet equally important interaction between an insect and plant resources. Plant-insect interactions recorded within the Wamsutta Formation represent some of the oldest documented occurrences of non-cryptogam gall damage and oviposition.
The earliest galling activity can be traced to a liverwort in the Middle Devonian Period (385 Ma70;), attributable to a fungal galler. Earlier work has shown that, after a 70-million-year hiatus in the record, gall activity was recorded in the Moscovian stage of the Pennsylvanian (315–312 Ma), on multiple examples of Calamites sp. stems found in Limburg (Netherlands). Late Pennsylvanian galls were subsequently found on other horsetails, marattialean ferns, and, possibly, conifers70,71,72. Eventually, small, early hemipteroid gallers diversified on seed-plant foliage during the Permian Period73. Preserved galls from the Wamsutta Formation occur on stem tissues of Sigillaria sp., and is consistent with the observation that galls did not begin to utilise leaf laminae until the early Permian (after ~300 Ma)70,74.
The earliest external ovipositors capable of slicing into plant tissues are seen in insects during the Mississippian–Pennsylvanian transition74. During this time and into the Pennsylvanian, ovipositing insects capable of producing lesions on plants included Odonatoptera, Palaeodictyopterida, Polyneoptera (Dictyoptera, Orthopterida), Paraneoptera, and some holometabolan clades74,75,76. While slicing ovipositors are found in Late Palaeozoic insects, the fossil record of endophytic oviposition lesions is rare compared to the Late Mesozoic and Cenozoic records77. One instance of endophytic oviposition known from the Late Carboniferous (Late Pennsylvanian; 304–299 Ma) was documented in the Graissessac Basin (Hérault, France) by Béthoux et al.75. Oviposition lesions occurring on Cordaites sp. in the Wamsutta Formation (Fig. 5C), therefore, pushes back observed occurrences of these insect-plant interactions by ~14 Ma.
The earliest piercing and sucking damage can be traced to the Early Devonian on a trimerophyte host, but other instances of this damage type do not appear until the early to middle Pennsylvannian74. We observe numerous examples of piercing and sucking damage on Cordaites sp. from the Wamsutta Formation, demonstrating that this type of interaction was relatively common by that time (e.g., Figure 5D). By the Pennsylvanian-Permian boundary, piercing-and-sucking damage occurred prominently on a number of free sporing and seed plant species78.
A most informative result of our coarse-grained herbivory analysis of 880 individual leaf specimens from the Lantern North assemblage is the presence of three arthropod functional feeding groups that accessed internal tissues: piercing-and-sucking, ovipositing, and galling. By contrast, damage from external modes of feeding such as the hole, margin, and surface feeding as well as skeletonization are largely absent. Internal modes of feeding such as seed predation and borings are also absent, although pathogens are present (see below). The reason for this overall pattern of feeding is initially puzzling. The oldest known winged insect is an orthopteridan79 that had mandibulate (chewing) mouthparts and existed at 324 Ma during the latest Mississippian (Lower Carboniferous), associated with palaeodictyopterid insects that had piercing-and-sucking mouthparts80. Both groups likely had sword-like, slicing ovipositors allowing for the placement of eggs in hidden locations such as within plant tissues and below the soil surface.
The evidence we present is consistent with the inference that if herbivorous mandibulate insects were present in the Early Pennsylvanian, then we should expect to see some evidence of this feeding style in the plant-insect interaction record in the form of margin feeding, hole feeding, or surface feeding. We do not observe evidence for this, even though the fossil record clearly indicates that several lineages of mandibulate insects were present during the Early Pennsylvanian. The presence of herbivorous margin feeding damage in Mississippian age deposits that preceded the earliest winged insects81 could be attributed to apterygote insects such as bristletails, or possibly myriapods. Nevertheless, the predominance of piercing-and-sucking feeding modes throughout the Pennsylvanian is demonstrated in the permineralized82 and compression72 fossil records, as well as stylate mouthpart occurrence, indicated by the observation that somewhat more than 50 percent of Palaeozoic insect species had stylate mouthparts83, and many of those species bore slicing ovipositors. With the fossil record of oviposition lesions72,75, it appears that puncturing and slicing through plant tissues was the predominant mode of accessing plant resources before chewing modes of feeding became dominant, as seen in the later Pennsylvanian78,84, including externally feeding, eruciform caterpillars76. Galling70,72 likely was a lesser feeding mode. Taken together, these data suggest that external modes of insect herbivory appeared relatively late in the Palaeozoic.
While Middle Pennsylvanian coal-bearing deposits such as Mazon Creek have been pivotal in our understanding of early terrestrial ecosystems1,2,3, the Early Pennsylvanian Wamsutta Formation illuminates the complexity of late Palaeozoic terrestrial ecosystems by revealing anatomy, behaviour, and organism interactions within the context of a rarely preserved proximal clastic environment. Exceptionally well-preserved trace fossils of vertebrates (synapsids, diadectomorphs, amniotes, temnospondyl amphibians, fish) as well as trace and body fossils of invertebrates (adult and nymphal pterygote and apterygote insects, myriapods, xiphosurans, thelyphonids, ricinuleids), along with a recently expanded list of 131 plant fossil-taxa, that includes 83 distinct foliage morphotypes, support this conclusion. Plant-insect interactions (3 functional feeding groups) preserved in the Wamsutta include the earliest currently known evidence of insect oviposition and some of the oldest documented occurrences of non-cryptogam gall damage in the fossil record. Indeed, by the very nature of its taphonomy, Wamsutta fossils provide data seldom seen at other body-fossil-dominated Lagerstätten36,38,39. Most types of trace fossils inherently occur in situ, including those of the Wamsutta, which allows for a more accurate reconstruction of the depositional environment and community-level interactions. Additionally, given that the trace fossils, by definition, were made while their producers were alive, they provide direct anatomical, behavioural, and interactional data and insights that cannot be extracted from body fossils alone. Thus, the Wamsutta Lagerstätte will serve as a key reference for future palaeoecological and phylogenetic reconstructions of these pivotal groups and increase our understanding of terrestrial life in a seldom sampled Palaeozoic environment.
Methods
Fossil specimens used in this work were collected in the field at the LN site in southeastern Massachusetts, USA (Fig. 1) beginning in 2007. Where outcrops likely to host fossil material (i.e., fine sandstones and shale) were identified, bedding planes were exposed and closely inspected. Fossils were extracted from these units using hand tools. Each specimen (trace fossil, body fossil, plant fossil) recovered was located in the stratigraphy, mapped with GPS coordinates and later georeferenced to aerial photographs. Areas of wider exposure near the LN site were correlated to the site stratigraphy using the local structural dip and linked to the fossil-bearing strata. These larger outcrops were then used to describe the stratigraphy and sedimentology of the fossil-bearing units (Supplementary Note 1). For identification and analysis, fossils were photographed in the lab using low-angle lighting and a Nikon D3X digital camera equipped with a Micro-Nikkor AF-S 60 mm f/2.8 G ED macro lens. A Keyence digital 130 microscope was used to investigate the smallest features, and photos from it were focus-stacked using Adobe Photoshop 2021 (version 133 22.4.2).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Materials availability
All specimens mentioned in this text are accessioned at the Harvard University’s Museum of Comparative Zoology (MCZ) except for four invertebrate fossils which are from the Fossil Insect Collection of Division of Entomology at the University of Kansas Natural History Museum (these numbers start with SEMC). To access this material please contact the Collection Manager of Invertebrate Palaeontology Jessica Cundiff at the MCZ or Zack Falin, Collections Manager at the University of Kansas Natural History Museum.
References
DiMichele and Hook. Paleozoic Terrestrial Ecosystems. In: Terrestrial Ecosystems Through Time: Evolutionary Paleoecology of Terrestrial Plants and Animals, Behrensmeyer, (eds. A. K., Damuth, J. D., DiMichele, W. A., Potts, R., Sues, H-D., and Wing, S. L.) 205–325, Ch. 5 (The University of Chicago Press, 1992).
Scott, A. C. et al. The legacy of Charles Lyell: advances in our knowledge of coal and coal-bearing strata. In: Blundell, (eds. D. J. & Scott, A. C.) Lyell: The Past is the Key to the Present, 43, 243–260. (Geological Society of London, 1998).
Cleal, C. J., Opluštil, S., Thomas, B. A. & Tenchov, Y. Late Moscovian terrestrial biotas and palaeoenvironments of Variscan Euramerica. Neth. J. Geosci. 88, 181–278 (2010).
Wagner, R. H. & Winkler Prins, C. F. History and current status of the Pennsylvanian chronostratigraphic units: problems of definition and interregional correlation. Newsl. Stratigr. 49, 281–320 (2016).
Knight, J. A., Cleal, C. J. & Álvarez-Vázquez, C. The challenge of relating the Kasimovian to west European chronostratigraphy: a critical review of the Cantabrian and Barruelian substages of the stephanian. Geol. Soc., Lond., Spec. Publ. 535, 31–71 (2023).
Anderson, J. M., Anderson, H. M. & Cleal, C. J. Brief History of the Gymnosperms: Classification, Biodiversity, Phytogeography and Ecology. (South African National Biodiversity Institute, Pretoria, 2007).
Falcon-Lang, H. J., Benton, M. J., Braddy, S. J. & Davies, S. J. The Pennsylvanian tropical biome reconstructed from the Joggins Formation of Nova Scotia, Canada. J. Geol. Soc. 163, 561–576 (2006).
Anderson, L. I., Dunlop, J. A., Horrocks, C. A., Winkelman, H. M. & Eagar, R. M. C. Exceptionally preserved fossils from Bickershaw, Lancashire, UK (Upper Carboniferous, Westphalian A (Langsettian)). Geol. J. 32, 197–210 (1997).
Lomax, D. R., Robinson, P., Cleal, C. J., Bowden, A. & Larkin, N. R. Exceptional preservation of upper carboniferous (lower Westphalian) fossils from Edlington, Doncaster, South Yorkshire, UK. Geol. J. 51, 42–50 (2016).
Wagner, R. H. The extrabasinal elements in lower Pennsylvanian floras of the Maritime Provinces, Canada: description of Adiantites, Pseudadiantites and Rhacopteridium. Rev. Española de. Paleontol. ía 16, 187–207 (2001).
Gupta, N. S. & Briggs, D. E. Taphonomy Of Animal Organic Skeletons Through Time. In: Taphonomy: Aims and Scope. Topics in Geobiology. (eds. P. A. Allison, and D. J. Bottjer), 32, 199–221. (Dordrecht: Springer, 2011).
Karr, J. A. & Clapham, M. E. Taphonomic biases in the insect fossil record: shifts in articulation over geologic time. Paleobiology 41, 16–32 (2015).
van den Belt, F. J., van Hoof, T. B. & Pagnier, H. J. Revealing the hidden Milankovitch record from Pennsylvanian cyclothem successions and implications regarding late Paleozoic chronology and terrestrial-carbon (coal) storage. Geosphere 11, 1062–1076 (2015).
Savrda, C. E. Taphonomy of trace fossils. In Trace fossils 92–109. (Elsevier, 2007).
Skehan, J. W. et al. The Missisippian and Pennsylvanian (Carboniferous) systems in the United States–Massachusetts, Rhode Island, and Maine. (1979).
Skehan, J. W., J, S., Rast, N. & Mosher, S. Paleoenvironmental and tectonic controls of sedimentation in coal forming basins of southeastern New England. Geol. Soc. Am. Spec. Pap. 210, 9–30 (1986).
Mosher, S. Kinematic history of the Narragansett Basin, Massachusetts and Rhode Island: constraints on late Paleozoic plate reconstructions. Tectonics 2, 327–344 (1983).
Murray, D. P., Skehan, J. W. & Raben, J. Tectonostratigraphic relationships and coalification trends in the Narragansett and Norfolk basins, New England. J. Geodynamics 37, 583–611 (2004).
Thompson, M. & Hermes, O. D. Early rifting in the Narragansett Basin, Massachusetts-Rhode Island: evidence from Late Devonian bimodal volcanic rocks. J. Geol. 111, 597–604 (2003).
Scotese, C. R. An atlas of Phanerozoic paleogeographic maps: the seas come in and the seas go out. Annu. Rev. Earth Planet. Sci. 49, 679–728 (2021).
Lyons, P. C. Report on the bedrock geology of the Narragansett Basin, Massachusetts and Rhode Island 77, 816 (US Geological Survey, 1977).
Chute, N. E. 1964. Geology of the Norfolk Basin Carboniferous Sedimentary Rocks and the Various Igneous Rocks of the Norwood and Blue Hills Quadrangles. in Guidebook to Field Trips in the Boston Area and Vicinity, edited by J. W. Skehan, S. J., 91–114, (New England Intercollegiate Geol. Conf., 56th Ann. Mtg., Boston College, Chestnut Hill, MA, 1964).
Knox, A. S. A Carboniferous flora from the Wamsutta Formation of southeastern Massachusetts. Am. J. Sci. 242, 1390–138 (1944).
Lyons, P. C., Tiffney, B. & Cameron, B. Early Pennsylvanian age of the Norfolk Basin, southeastern Massachusetts, based on plant megafossils. Geological Society of America Memoirs, 146, 181–197 (1976).
Lyons, P. C. & Darrah, W. C. A late Middle Pennsylvanian flora of the Narragansett basin, Massachusetts. Geol. Soc. Am. Bull. 89, 433–438 (1978).
Lyons, P. C. & Sproule, R. G. The fossil flora and age of the Wamsutta red beds (Middle Pennsylvanian), Narragansett Basin, southeastern Massachusetts, USA and correlation with the Cumberland Group of the Maritime Provinces of Canada. Atl. Geol. 54, 315–334 (2018).
Severson, R. H. & Boothroyd, J. C. Depositional environments, facies associations, and coal occurrence in Carboniferous sediments of the Narragansett Basin. Geol. Soc. Am. Abstr. Prog. 13, 176 (1981).
Cazier, E. C. Late Paleozoic tectonic evolution of the Norfolk Basin, southeastern Massachusetts. J. Geol. 95, 55–73 (1987).
Müller, J. & Reisz, R. R. An early captorhinid reptile (Amniota, Eureptilia) from the Upper Carboniferous of Hamilton, Kansas. J. Vertebrate Paleontol. 25, 561–568 (2005).
Müller, J., Berman, D. S., Henrici, A. C., Martens, T. & Sumida, S. S. The basal reptile Thuringothyris mahlendorffae (Amniota: Eureptilia) from the Lower Permian of Germany. J. Paleontol. 80, 726–739 (2006).
Müller, J., Li, J. L. & Reisz, R. R. A new bolosaurid parareptile, Belebey chengi sp. nov., from the Middle Permian of China and its paleogeographic significance. Naturwissenschaften 95, 1169–1174 (2008).
Ford, D. P. & Benson, R. B. The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae. Nat. Ecol. Evolution 4, 57–65 (2020).
Voigt, S. & Ganzelewski, M. Toward the origin of amniotes: Diadectomorph and synapsid footprints from the early Late Carboniferous of Germany. Acta Palaeontologica Polonica 55, 57–72 (2009).
Marchetti, L. et al. Footprints of the earliest reptiles: Notalacerta missouriensis–Ichnotaxonomy, potential trackmakers, biostratigraphy, palaeobiogeography and palaeoecology. Annales Societatis Geologorum Poloniae 90, 271–290 (2020).
Buchwitz, M., Jansen, M., Renaudie, J., Marchetti, L. & Voigt, S. Evolutionary change in locomotion close to the origin of amniotes inferred from trackway data in an ancestral state reconstruction approach. Front. Ecol. Evolution 9, 674779 (2021).
Knecht, R. J., Engel, M. S. & Benner, J. S. Late Carboniferous paleoichnology reveals the oldest full-body impression of a flying insect. Proc. Natl Acad. Sci. 108, 6515–6519 (2011).
Logghe, A. et al. A twig-like insect stuck in the Permian mud indicates early origin of an ecological strategy in Hexapoda evolution. Sci. Rep. 11, 20774 (2021).
Knecht, R. J., Benner, J. S., Dunlop, J. A. & Renczkowski, M. D. The largest Palaeozoic whip scorpion and the smallest (Arachnida: Uropygi: Thelyphonida); a new species and a new ichnospecies from the Carboniferous of New England, USA. Zoological Journal of the Linnean Society, (2023).
Minter, N. J. & Braddy, S. J. Walking and jumping with Palaeozoic apterygote insects. Palaeontology 49, 827–835 (2006).
Getty, P. R., Sproule, R., Wagner, D. L. & Bush, A. M. Variation in wingless insect trace fossils: insights from neoichnology and the Pennsylvanian of Massachussetts. Palaios 28, 243–258 (2013).
Benner, J. S., Knecht, R. J. & Engel, M. S. Tonganoxichnus: a revision of the ichnogenus with new material from Massachusetts. Ichnology: papers from Ichnia III. Edited by D. McIllroy. Geological Association of Canada, Miscellaneous Publication, 9, 31–43 (2015).
Getty, P. R., Sproule, R., Stimson, M. R. & Lyons, P. C. Invertebrate trace fossils from the Pennsylvanian Rhode Island formation of Massachusetts, USA. Atl. Geol. 53, 185–206 (2017).
Mángano, M. G., Buatois, L. A., Maples, C. G. & Lanier, W. P. Tonganoxichnus a new insect trace from the Upper Carboniferous of eastern Kansas. Lethaia 30, 113–125 (1997).
Mángano, M. G., Labandeira, C. C., Kvale, E. P. & Buatois, L. A. The insect trace fossil Tonganoxichnus from the Middle Pennsylvanian of Indiana: paleobiologic and paleoenvironmental implications. Ichnos: Int. J. Plant Anim. 8, 165–175 (2001).
Labandeira, C., Beall, B. & Hueber, F. Early insect diversification: evidence from a lower Devonian bristletail from Quebec. Science 242, 913–916 (1988).
Kukalová-Peck, J. Fossil history and the evolution of hexapod structures. In Naumann, I. D., Carne, P. B., Lawrence, J. F., Nielsen, E. S., Spradbery, J. P., Taylor, R. W., Whitten, M. J. and Littlejohn, M. J. (eds.): Insects of Australia, 2nd Edn., 141–179. (Melbourne University Press and Cornell University Press, 1990).
Labandeira, C. C. & Currano, E. D. The fossil record of plant-insect dynamics. Annu. Rev. Earth Planet. Sci. 41, 287–311 (2013).
Valdez-Mondragón, A. & Juárez-Sánchez, A. R. A new epigean species of ricinuleid of the genus Pseudocellus (Arachnida: Ricinulei: Ricinoididae) from a tropical sub-deciduous forest in Oaxaca, Mexico. J. Arachnol. 48, 329–338 (2021).
Selden, P. A. Revision of the fossil ricinuleids. Trans. R. Soc. Edinb.: Earth Sci. 83, 595–634 (1992).
Petrunkevitch, A. I. Palaeozoic Arachnida of Illinois. An inquiry into their evolutionary trends. Ill. State Sci. Pap. 3, 8–72 (1945).
Brauckmann, C. Neue Arachniden (Ricinuleida, Trigonotarbida) aus dem Namurium B von Hagen-Vorhalle (Ober-Karbon; West-Deutschland). Dortm. Beitr. äge der Landeskd., Naturwissenschaftliche Mitteilungen 21, 97–109 (1987).
Whalen, N. & Selden, P. A. A new, giant Ricinuleid (Arachnida, Ricinulei), from the Pennsylvanian of Illinois, and the identification of a new, ontogenetically stable, diagnostic character. J. Paleontol. 95, 601–612 (2021).
Wunderlich, J. New extinct taxa of the arachnid order Ricinulei, based on new fossils preserved in mid Cretaceous Burmese amber. Beitr. äge zur. Araneol. 10, 48–71 (2017).
Bicknell, R. D. & Pates, S. Pictorial atlas of fossil and extant horseshoe crabs, with focus on Xiphosurida. Front. Earth Sci. 8, 449663 (2020).
Bicknell, R. D. & Pates, S. Xiphosurid from the Tournaisian (Carboniferous) of Scotland confirms deep origin of Limuloidea. Sci. Rep. 9, 17102 (2019).
Koll, R. A. & DiMichele, W. A. Dominance-diversity architecture of a mixed hygromorphic-to-xeromorphic flora from a botanically rich locality in western equatorial Pangea (lower Permian Emily Irish site, Texas, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 563, 110132 (2021).
Thomas, B. A. & Cleal, C. J. Distinguishing Pennsylvanian-age lowland, extra-basinal and upland vegetation. Palaeobiodiversity Palaeoenvironments 97, 273–293 (2017).
Bashforth, A. R. et al. The environmental implications of upper Paleozoic plant-fossil assemblages with mixtures of wetland and drought-tolerant taxa in tropical Pangea. Geobios 68, 1–45 (2021).
Kazuhide, Nara Ectomycorrhizal networks and seedling establishment during early primary succession. N. Phytologist 169, 169–178 (2006).
Hogberg, P., Plamboeck, A. H., Taylor, A. F. & Fransson, P. M. A. Natural 13C abundance reveals trophic status of fungi and host-origin of carbon in mycorrhizal fungi in mixed forests. Proc. Natl Acad. Sci., USA. 96, 8534–8539 (1999).
Dickie, Ian A. et al. The emerging science of linked plant–fungal invasions. N. Phytologist 215, 1314–1332 (2017).
Zeilinger, S. et al. Friends or F\foes? Emerging insights from fungal interactions with plants. Microbiol. Rev. 40, 182–207 (2016).
Gorzelak, M. A., Asay, A. K., Pickles, B. J. & Simard, S. W. Inter-plant communication through mycorrhizal networks mediates complex adaptive behaviour in plant communities. AoB Plants 7, plv050 (2015).
Simard, S. W. et al. Mycorrhizal networks: mechanisms, ecology and modelling. Fungal Biol. Rev. 26, 39–60 (2012).
Horton, T. R. (ed). Mycorrhizal networks. Ecological Studies. 224, Dordrecht: Springer Netherlands. (2015).
Duddridge, J. A., Malibari, A. & Read, D. J. Structure and function of mycorrhizal rhizomorphs with special reference to their role in water transport. Nature 287, 834–836 (1980).
Smith, M. R. Cord-forming palaeozoic fungi in terrestrial assemblages. Botanical J. Linn. Soc. 180, 452–460 (2016).
Krings, M., Dotzler, N., Taylor, T. N. & Galtier, J. A. late Pennsylvanian fungal leaf endophyte from Grand-Croix, France. Rev. Palaeobot. Palynol. 156, 449–453 (2009).
Labandeira, C. C., Wilf, P., Johnson, K. R. & Finnegan Marsh. Guide to Insect (and other) Damage Types On Compressed Plant Fossils. (Smithsonian Institution, Washington D.C, 2007).
Labandeira, C. Ecology and evolution of gall-inducing arthropods: the pattern from the terrestrial fossil record. Front. Ecol. Evolution 9, 632449 (2021).
Labandeira, C. C. & Phillips, T. L. A Carboniferous petiole gall: insight into early ecologic history of the Holometabola. Proc. Natl Acad. Sci., USA. 93, 8470–8474 (1996).
Correia, P. et al. The history of herbivory on sphenophytes: a new calamitalean with an insect gall from the Upper Pennsylvanian of Portugal and a review of arthropod herbivory on an ancient lineage. Int. J. Plant Sci. 181, 387–418 (2020).
Schachat, S. R. & Labandeira, C. C. Evolution of a complex behavior: the origin and initial diversification of foliar galling by Permian insects. Sci. Nat. 102, 14 (2015).
Labandeira, C. The four phases of plant-arthropod associations in deep time. Geologica Acta 4, 409–438 (2006).
Bethoux, O., Galtier, J. & Nel, A. Earliest evidence of insect endophytic oviposition. PALAIOS 19, 408–413 (2004).
Haug, J. T., Labandeira, C. C., Santiago-Blay, J. A., Haug, C. & Brown, S. Life habits, hox genes, and affinities of a 311 million-year-old holometabolan larva. BMC Evolut. Biol. 15, 1–10 (2015).
Vasilenko, D. V. & Rasnitsyn, A. P. Fossil ovipositions of dragonflies: review and interpretation. Paleontological J. 41, 1156–1161 (2007).
Xu, Q.-Q., Jin, J.-H. & Labandeira, C. C. Williamson drive: herbivory of a north-central Texas flora of latest Pennsylvanian age showing discrete component community structure, early expansion of piercing and sucking, and plant counterdefenses. Rev. Palaeobot. Palynol. 251, 28–72 (2018).
Prokop, J., Nel, A. & Hoch, I. Discovery of the oldest known Pterygota in the lower Carboniferous of the Upper Silesian Basin in the Czech Republic (Insecta: Archaeorthoptera). Geobios 38, 383–387 (2005).
Donovan, M. P., Schachat, S. R. & Monarrez, P. M. Ecological and evolutionary responses of terrestrial arthropods to Middle–Late Pennsylvanian environmental change. Geol. Soc., Lond., Spec. Publ. 535, 439–480 (2023).
Iannuzzi, R. & Labandeira, C. C. The oldest record of external foliage feeding and the expansion of insect folivory on land. Ann. Entomological Soc. Am. 101, 79–94 (2008).
Labandeira, C. C. & Phillips, T. L. Insect fluid-feeding on Upper Pennsylvanian tree ferns (Palaeodictyoptera, Marattiales) and the early history of the piercing-and-sucking functional feeding group. Ann. Entomological Soc. Am. 89, 157–183 (1996).
Carpenter, F. M. Adaptations among Paleozoic insects. In proceedings of the first North American Paleontological Convention, Lawrence, Kansas, Allen Press 2, 1236–1251 (1971).
Scott, A. C. & Taylor, T. N. Plant/animal interactions during the upper Carboniferous. Botanical Rev. 49, 259–307 (1983).
Acknowledgements
The authors thank Sebastian Voigt, who provided helpful discussions on the vertebrate trace fossil material. David Brask who provided access to the site. Help with field relationships was provided by Anne Gardulski, Meg Thompson, and Daniel Murray. RJK acknowledges funding support from: the National Science Foundation (NSF) GRFP, Tufts University Olmstead Fellowship, a Stephen J.Gould Grant from the Palaeontological Society, and a Student Research Grant from the Geological Society of America Northeast Section. AS acknowledges the Harvard Society of Fellows and the James S. McDonnell Foundation for research support. The participation of MSE was supported by CONCYTEC through the PROCIENCIA (Interinstitutional Alliances for Doctorate Programmes) programme (contract PE501084299-2023-PROCIENCIA-BM). MMF was supported by an NSF OEB summer research REU programme (award DBI-1757780). Published by a grant from the Wetmore Colles fund (to RJK), the University of Tennessee Open Publishing Support Fund (to JSB), and the Harvard William F. Milton Fund (to AS).
Author information
Authors and Affiliations
Contributions
R.J.K. and J.S.B. discovered L.N. site and strata; R.J.K., J.S.B., A.S., M.D.R., D.A.W., M.M.F., and S.L.E. collected specimens from the site; R.J.K., J.S.B., and A.H.K. analysed and wrote about the geology; R.J.K., J.S.B., and M.M.F. analysed and wrote about the vertebrates; R.J.K., J.S.B., M.S.E., J.A.D., P.A.S., and N.E.P. analysed and wrote about the invertebrates; R.J.K., A.S., L.A., C.J.C., C.C.L., C.F.E., M.D.R., D.A.W., and S.L.E. analysed and wrote about the plants, spores, and fungal rhizomorphs; R.J.K., A.S., L.A., C.C.L., and D.A.W. analysed and wrote about the organismal interactions; all authors edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks David Broussard, Joachim Haug, Hermann Pfefferkorn and Artai Santos for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Knecht, R.J., Benner, J.S., Swain, A. et al. Early Pennsylvanian Lagerstätte reveals a diverse ecosystem on a subhumid, alluvial fan. Nat Commun 15, 7876 (2024). https://doi.org/10.1038/s41467-024-52181-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-52181-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
