


















« Prev Next »
"When in the offspring of two different animal races one sex is absent, rare, or sterile, that sex is the heterozygous [heterogametic] sex." -J. B. S. Haldane (1922)
Haldane's simple observation, now known as Haldane's rule, still interests and perplexes biologists studying the genetics of speciation. Why should hybrids of the heterogametic sex, or the one with different sex chromosomes (e.g., XY in mammals), be more afflicted than hybrids of the homogametic sex (e.g., XX in mammals)? One reason that biologists who study speciation focus on Haldane's rule is that it is one of the few empirical generalizations about speciation that holds true in many different groups of animals. For instance, this pattern is followed in groups of organisms in which males are the heterogametic sex (such as flies and mammals), as well as in groups of organisms in which females are the heterogametic sex (e.g., birds and butterflies). The prevalence of this pattern suggests that it is generated by common mechanisms in diverse organisms.
Haldane's original observations concerned just the fertility and viability of hybrids, but Haldane's rule can also be extended to other traits. Indeed, in hybrids, the members of the heterogametic sex tend to suffer more adverse morphological effects than their homogametic counterparts. For instance, Wade and Johnson (1994) found that hybrid males produced by the mating of two species of flour beetle (Tribolium castaneum and Tribolium freemani) often have deformed legs and antennae. Such deformities are rare within either parent species of flour beetle. Interestingly, at the normal rearing temperature (29°C), hybrid females produced by this same mating have deformities only occasionally. However, at higher temperatures (e.g., 34°C), a considerable fraction of hybrid females are also deformed (Wade et al., 1999).
But what are the genetic and evolutionary causes of this pattern? Specifically, is Haldane's rule due to properties of sex chromosomes or properties related to sex itself? Most evolutionary geneticists think Haldane's rule is the result of multiple factors in both categories.
Haldane's Rule and Sex Chromosomes
One simple way that Haldane's rule could arise via properties of sex chromosomes is if the interactions between an X chromosome from one species and a Y chromosome from the other species lead to hybrid incompatibility; here, such X-Y interactions would only occur in the heterogametic sex. Unfortunately, this simple explanation is not well-supported. First, this XY model would predict that male and female hybrids are equally affected in groups of species in which males lack an X chromosome (i.e., species in which females are XX and males are XO). In grasshoppers, however, male hybrids are more frequently inviable or sterile than female hybrids (Wu & Davis, 1993; Coyne & Orr, 2004). Second, genetic analyses reveal that X-Y interactions rarely contribute to hybrid incompatibility in Drosophila. In fact, in many cases, the Y chromosome does not contribute to hybrid sterility and inviability at all (Johnson et al., 1993). Moreover, even when the Y chromosome does have an effect on hybrids, it may be interacting with genes on the autosomes and not with X-linked genes (Johnson et al., 1992).
Given these problems with the XY model, investigators have proposed various other models in the attempt to explain Haldane's rule. For instance, in the 1940s, Herman Joseph Muller proposed that Haldane's rule arose because the heterogametic sex had an imbalance of X chromosomes and autosomes. Consider the example of a cross between two species in which males are the heterogametic sex. In this case, female F1 hybrids would have one X chromosome and one set of autosomes from each parent; meanwhile, male F1 hybrids would have one X chromosome from their female parent and one Y chromosome from their male parent, as well as one set of autosomes from each parent. The F1 males will thus suffer an X-A imbalance because they lack an X chromosome from one of their parental species but have autosomes from both of the parental species.
In the 1990s, Allen Orr and Michael Turelli proposed a refinement of Muller's model in which they noted that hybrid males would suffer more than females only if the alleles that were involved in hybrid incompatibility were somewhat to completely recessive in their effects on the hybrids (Turelli & Orr, 1995). According to this model, hybrid females with X chromosomes from both parental species would be sheltered from the effects of recessive negative alleles, but males (who have only one X from one of the parental species) would not. In contrast, if the negative effects were dominant, then hybrid females would actually suffer more than males. Orr and Turelli's model, now known as the dominance model (or dominance theory), put Muller's ideas into an explicit theoretical framework, specifying exactly when allele effects should cause Haldane's rule.
Hybrid Inviability and Sterility
Most evolutionary biologists currently think that the dominance model explains at least some features of Haldane's rule, but it may not be the sole explanation for the pattern. For example, Chung-I Wu and colleagues noted that in Drosophila and in mammals, hybrid male sterility not only evolves much faster than hybrid female sterility, but it also evolves faster than hybrid inviability in either sex (Wu & Davis, 1993; Wu et al., 1996). This faster rate of hybrid male sterility does not occur because sterility is more mutable; in fact, mutagenesis studies with Drosophila reveal that the number of mutations that cause inviability greatly exceeds the number that cause sterility. Nonetheless, Haldane's rule is usually more closely followed in cases of sterility than in cases of inviability. For instance, in one study, investigators noted 199 cases in which hybrid F1 males were sterile and hybrid F1 females were fertile, and three cases in which the F1 males were fertile and the F1 females were sterile. In contrast, when looking at hybrid inviability, the researchers noted only 14 cases that followed Haldane's rule and nine that disagreed with it (Wu et al., 1996). These findings led to the argument that some property of maleness must also drive Haldane's rule in groups in which males are the heterogametic sex (Wu & Davis, 1993; Wu et al., 1996).
But what could be driving a faster rate of hybrid sterility—and hence, Haldane's rule—in groups of organisms with male heterogamety? One likely cause is sexual selection. As a rule, sexual selection is more intense for males than females. If traits in males have an accelerated rate of evolution because of sexual selection, these traits will more likely be affected negatively in the hybrids. Although there is no direct proof that sexual selection has contributed to the sterility aspect of Haldane's rule, a substantial body of circumstantial evidence supports this idea. One line of supporting evidence is that many genes expressed in the male reproductive tract in both mammals and Drosophila have an accelerated rate of evolution (Torgerson et al., 2002). However, it is important to note that sexual selection, or any other explanation for faster male evolution, can only explain the acceleration of hybrid male sterility in organisms wherein males are the heterogametic sex. In birds and other groups of organisms with females as the heterogametic sex, faster male evolution would lead to exceptions to Haldane's rule.
Testing Haldane's Rule with Hybrid Fruit Flies
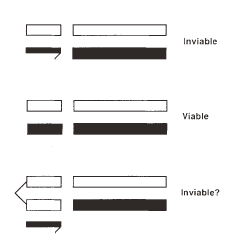
Figure 1: Test of the genetic basis of Haldane's rule
The chromosomes of one species (e.g., D. simulans) are shown in white and those of the other species (e.g., D. teissieri) in black. Sex chromosomes are shown at the left (X on top; Y, with a "hook", on the bottom). Haploid sets of autosomes shown at the right. All genotypes carry cytoplasm from the "white" species. The top genotype represents F1 male hybrids and the middle genotype represents F1 female hybrids. The bottom genotype depicts hybrid females who carry an attached X chromosome from the white species. The traditional explanation of Haldane's rule predicts that this "unbalanced" genotype will be inviable. (Population genetics models show that this theory requires an additional assumption: the alleles causing reproductive isolation act as loss-of-function mutations; that is, more recessive alleles must have greater homozygous effects on hybrid fitness.)
© 1993 Nature Publishing Group Orr, H. A. Haldane's rule has multiple genetic causes. Nature 361, 532 (1993). All rights reserved. 
Over the years, investigators have carried out a number of experiments using various species of Drosophila in an attempt to test possible causes of Haldane's rule. For instance, in one notable study, researcher H. Allen Orr tested the dominance model by making F1 hybrid females as imbalanced as F1 hybrid males. Orr knew that if imbalance was a sufficient explanation for Haldane's rule, then the imbalanced hybrid females would be as inviable as their male counterparts. He also knew that in crosses between D. simulans and D. teissieri, F1 hybrid males were typically inviable, while females were viable. To construct his imbalanced hybrid females, Orr took advantage of a special stock of D. simulans. In this stock, called attached-X, females have both of their X chromosomes attached to one another. Female hybrids resulting from crosses between attached-X D. simulans females and D. teissieri males therefore receive both of their X chromosomes from D. simulans, a Y chromosome from D. teissieri, and one set of autosomes from both D. simulans and D. teissieri. Orr found that these females were, in fact, inviable, a result that supported the dominance model (Figure 1; Orr, 1993).
Similar tests using attached-X females to produce imbalanced F1 females have been carried out with other species pairs of Drosophila; in general, the imbalanced females are inviable in crosses in which the F1 males are inviable, but fertile in crosses in which the F1 males are sterile. These results suggest that chromosome imbalance, presumably due to the dominance model, is sufficient to explain the viability aspect of Haldane's rule, but it is not sufficient to explain the sterility aspect (Coyne, 1985; Orr, 1993; Wu & Davis, 1993).
Thus, to learn more about the sterility portion of Haldane's rule, Orr and his then-graduate student Daven Presgraves compiled comparative data from two different genera of mosquitoes—Anopheles and Aedes—that differ in the nature of their sex chromosomes (Presgraves & Orr, 1998). Those species in the genus Anopheles have Y chromosomes that are morphologically different from their X chromosomes. These mosquitoes' Y chromosomes also have fewer genes than their X chromosomes. Moreover, in the members of this species, large regions of the Y chromosome do not recombine with the X chromosome. Such distinct chromosomes are known as degenerative sex chromosomes. In contrast, the X and Y chromosomes in the genus Aedes are quite similar; these chromosomes are said to be nondegenerate. Orr and Presgraves hypothesized that if only the dominance model is operating, the mosquito species pairs with degenerate sex chromosomes should exhibit Haldane's rule, but the species pairs with nondegenerate sex chromosomes should not. Thus, if Haldane's rule appeared in Aedes species pairs, some other mechanism (like sexual selection) must be at work.
Indeed, in Aedes, Orr and Presgraves found many more cases of unisexual hybrid sterility in which the males were sterile than cases in which the females were sterile. These data demonstrated that the presence of degenerate sex chromosomes (and thus the dominance model) is not needed for Haldane's rule for sterility. The likely mechanism for the increased hybrid male sterility in this genus is faster male evolution, presumably by sexual selection. On the other hand, male-biased hybrid sterility is more pronounced in Anopheles mosquitoes than it is in Aedes mosquitoes. This finding suggests that dominance may in fact play a role in the sterility aspect of Haldane's rule. Furthermore, Haldane's rule for inviability is present in Anopheles but not in Aedes; the most likely explanation for this finding is that the dominance model is sufficient to explain the viability aspect of Haldane's rule in mosquitoes.
The Faster-X Model
The dominance and faster-male models are well supported in Drosophila and, to some extent, in mammals. Support for these models doesn't rule out the prospect that other factors may also be at work, however.
For example, a third model explains Haldane's rule as a consequence of faster evolution on the X chromosome (Coyne & Orr, 1989). This faster-X model assumes that most advantageous alleles are partially recessive. Note that this recessive effect is in the pure species, not in the hybrids, as is postulated in the dominance model above. Population genetics theory predicts that partially recessive alleles would accumulate more readily on the X chromosome because when new alleles enter the population through mutation, they generally reside in heterozygotes. Recessive advantageous alleles on autosomes would not be expressed in heterozygotes and thus would not display a selective advantage when at low frequency. On the other hand, such X-linked alleles would be expressed immediately in the heterogametic sex. If these alleles are sex-biased, and alleles that are beneficial in the pure-species genetic background are deleterious in the hybrid genetic background, then the heterogametic sex would therefore be more afflicted in hybrids (per Haldane's rule). This faster-X model recently received notable empirical support from Drosophila genetic studies, which showed that the density of genetic factors that cause hybrid male sterility when introgressed from one species to another is greater on the X chromosome than on the autosomes (Masly & Presgraves, 2007).
Remaining Challenges in the Study of Haldane's Rule
Haldane's rule is often accurately described as a composite phenomenon; the relative importance of different causes (dominance model vs. faster-male model vs. faster-X model) may differ among the viability, morphological, and sterility aspects of Haldane's rule. Another challenge to the study of the genetic and evolutionary causes of Haldane's rule is that, although the pattern is very general (i.e., it is seen in flies, mammals, birds, butterflies, and other major groups of organisms), the most detailed genetic data come mainly from just one genus, Drosophila. Can the conclusions drawn from Drosophila studies be generalized to other groups? Exacerbating this problem is the lack of genetic studies in a hybrid that has heterogametic females. The insect order Lepidoptera (butterflies and moths) may contain the best candidate species for such studies in the years to come.
References and Recommended Reading
Coyne, J. A. The genetic basis of Haldane's rule. Nature 314, 736–738 (1985) (link to article)
Coyne, J. A., & Orr, H. A. Two rules of speciation. In Speciation and Its Consequences, ed. D. Otte and J. Endler (Sunderland, MA, Sinauer Associates, 1989), 180–207
———. Speciation (Sinauer Associates, Sunderland, MA, 2004.)
Haldane, J. B. S. Sex-ratio and unisexual sterility in hybrid animals. Journal of Genetics 12, 101–109 (1922)
Johnson, N. A., et al. A test of reciprocal X-Y interactions as a cause of hybrid sterility in Drosophila. Nature 358, 751–753 (1992) (link to article)
———. The effects of interspecific Y chromosome replacements on hybrid sterility within the Drosophila simulans clade. Genetics 135, 443–453 (1993)
Laurie, C. C. The weaker sex is heterogametic: 75 years of Haldane's rule. Genetics 147, 937–951 (1997)
Masly, J. P., & Presgraves, D. C. High-resolution genome-wide dissection of the two rules of speciation in Drosophila. PLoS Biology 5, e243 (2007) (link to article)
Muller, H. J. Isolating mechanisms, evolution, and temperature. Biological Symposium 6, 71–125 (1942)
Orr, H. A. Haldane's rule has multiple genetic causes. Nature 361, 532–533 (1993) (link to article)
Presgraves, D. C., & Orr, H. A. Haldane's rule in taxa lacking a hemizygous X. Science 282, 952–954 (1998)
Torgerson, D. G., et al. Mammalian sperm proteins are rapidly evolving: Evidence of positive selection in functionally diverse genes. Molecular Biology and Evolution 19, 1973–1980 (2002)
Turelli, M., & Orr, H. A. The dominance theory of Haldane's rule. Genetics 140, 389–402 (1995)
Wade, M. J., & Johnson, N. A. Reproductive isolation between two species of flour beetles, Tribolium castaneum and T. freemani: Variation among and within geographical populations of T. castaneum. Heredity 72, 155–162 (1994)
Wade, M. J., et al. Temperature effects and genotype by environment interactions (GEI) in hybrids: Haldane's rule in flour beetles. Evolution 53, 855–865 (1999)
Wu, C. I., & Davis, A. W. Evolution of postmating reproductive isolation—The composite nature of Haldane's rule and its genetic bases. American Naturalist 142, 187–212 (1993)
Wu, C. I., et al. Haldane's rule and its legacy: Why are there so many sterile males? Trends in Ecology and Evolution 11, 281–284 (1996)










