





















« Prev Next »

Like many other social mammals, primates often live in stable groups containing individuals of both sexes and of various ages. While much information concerning the social relationships of individuals can be amassed by patient direct observation of members of wild groups habituated to human observation, genetic analysis can provide important insights into movement of individuals between groups, how successful individuals are at breeding, and how individuals are related to one another.
Noninvasive Sampling of Primates
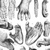
Because many wild primates are endangered, elusive, or both, researchers have found creative ways to collect DNA from their study subjects without handling them. Like forensic scientists, primate researchers use shed hair, chewed fruit, and even feces for DNA analysis (Figure 1). Arboreal primates pose particular challenges, but may be sampled by plucking hair using sticky blow darts, or by collecting shed hairs in ape night nests. The DNA extracted from noninvasive samples tends to be low in quantity and quality, making it challenging — but not impossible — to use it to conduct reliable analyses (Taberlet et al. 1999).

Figure 1: Noninvasive fecal sampling.
Gorilla (a) and chimpanzee (b) fecal samples in the field.
© 2012 Nature Education Photos courtesy of: (a) Martha Robbins, (b) Kevin Langergraber. All rights reserved. 
Rapidly Evolving DNA Sequences are Most Useful
The members of one or several wild primate groups may be closely related, and so to tell individuals apart genetically it is usually best to analyze segments of DNA that mutate rapidly and are therefore highly variable. Microsatellite loci, which are particular segments in the nuclear genome containing different numbers of repeated elements, are relied upon in studies focusing on the determination of family relationships between individuals (Figure 2). Another very useful and highly variable piece of DNA is found in the mitochondria, the organelles outside the nucleus where energy for the cell is generated. Mitochondrial DNA (mtDNA) can often be easily isolated from low-quality and low-quantity samples, since each mitochondrion usually contains multiple mtDNA copies, and each cell — depending on organism and tissue type — can harbor many mitochondria. Because these mtDNAs are only passed down from mother to offspring, and thus directly track the history of the maternal line, many studies of dispersal patterns have employed mtDNA markers (Di Fiore 2003). Paternal genetic history and male movements can be tracked by analyses of microsatellite or DNA sequence variation on the Y-chromosome (Vigilant & Guschanski 2009).
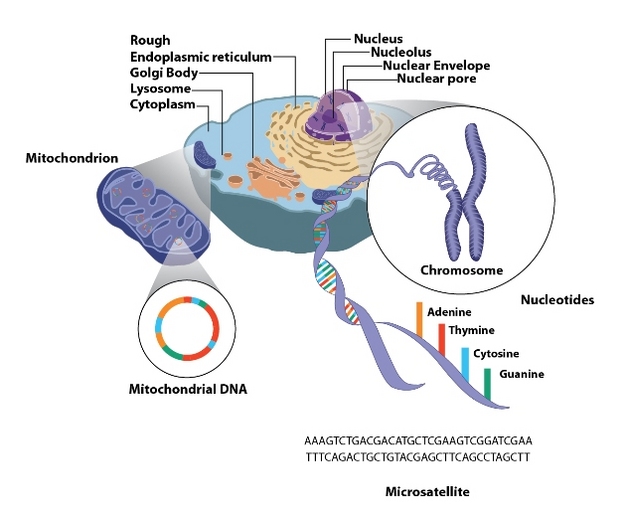
Figure 2: Location of mtDNA and nuclear DNA in an animal cell.
The mitochondria of an animal cell harbor maternally transmitted mtDNA. Nuclear DNA, which is transmitted by both parents, is found in the nucleus, and contains repeated elements termed microsatellites.
© 2012 Nature Education All rights reserved.
Dispersal Patterns
Individuals living in social groups often disperse into new groups before beginning breeding, probably to avoid the potentially deleterious effects of breeding with close relatives (inbreeding avoidance). But in many primate species, only the members of one sex leave their natal group (e.g., bonobos, chimpanzees). The ‘philopatric' members of the other sex spend their entire lives in their natal group while engaging in social relationships with lifelong associates of the same sex.
Because mtDNA sequences reflect the history and relationships of females, if females are moving between social groups, we expect the mtDNA sequences found in those groups to be more similar than we would find if females do not disperse. If instead, males are moving between groups, very little differentiation of Y-chromosomes between groups is expected. Unfortunately, because the structure of Y-chromosome variation varies greatly among species, it is currently difficult to assess in most primates. Therefore, inferences on dispersal rely on examining mtDNA variation or microsatellite variation at autosomal loci inherited on the chromosomes from both parents.
Like the majority of group-living mammals, most primates exhibit female philopatry and male emigration (Greenwood 1980), although patterns in New World primates are less well investigated (Fernandez-Duque et al. 2012). In Old World monkeys, such as baboons or macaques, social groups thus contain one or more extended matrilines of females and a changing cast of immigrant adult males. In addition to mtDNA, female Old World monkeys also inherit their social rank from their mothers, so members of particular matrilines are socially dominant over all members of other matrilines. When groups of rhesus macaques become too large, group splits typically result in members of matrilines staying together, underlining the importance of social relations among philopatric female kin (Widdig et al. 2006).
In species in which both sexes disperse, interesting patterns can emerge when genetic data are analyzed separately for each sex. For example, although both sexes disperse in mountain gorillas, the resulting structure of genetic variation in the entire adult population is consistent with that observed in just females, while no structure is observed in the males (Guschanski et al. 2008). Furthermore, the female pattern tracks the altitude and types of vegetation available. These observations suggest that while males disperse throughout the population, females do not disperse as far, and are influenced in their dispersal decisions by the type of habitat available. So instead of just choosing a group containing congenial potential mates, dispersing primates — especially females who need to find sufficient food to raise offspring — may take ecological factors into account as well.
Social Behaviour and Paternity
Male primates typically compete intensively for mating opportunities, and the advent of microsatellite genotyping using DNA from noninvasive samples in the 1990s allowed researchers to evaluate the consequences of male and female reproductive behaviour. Males may compete intensively for high social rank, and such social dominance appears beneficial for gaining access to females and excluding other males (Ellis 1995). However, male coalitions, alternative male mating strategies (such as sneaking copulations), and female preferences may reduce the effect of rank upon reproductive success (Wroblewski et al. 2009). In lemurs, females often become simultaneously sexually receptive, making it more difficult for any one male to monopolize reproduction (Kappeler, 2012).
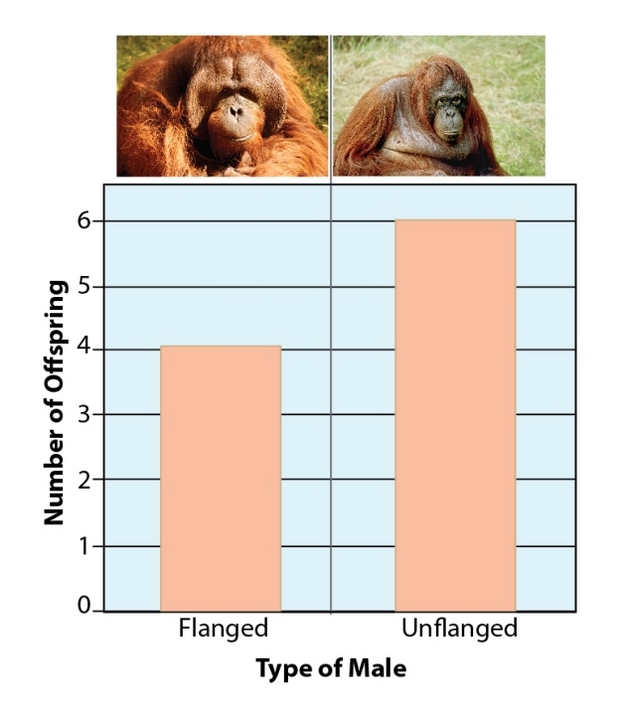
It is particularly hard to infer patterns of reproduction from observational data in primates that are largely solitary, such as orangutans. Adult male orangutans can differ greatly in appearance, and large males sporting cheek flanges call and advertise their presence on their territories, while smaller unflanged males quietly roam the forest. Genetic analysis of paternity in a sample of 10 wild orangutan offspring found that both types of males are successful in producing offspring, and it is suspected that, while the large males are attractive to females, the small males can employ coercive mating tactics (Figure 3) (Utami et al. 2002).
Figure 3: Number of offspring sired by flanged vs unflanged males.
Utami et al. (2002) found that unflanged males fathered about half of the offspring born over 15 years, by carrying out paternity analyses.
© 2012 Nature Education Photos courtesy of: flanged, Rudi Delvaux; unflanged, Marc Ancrenaz. All rights reserved.
Patterns of male social and reproductive dominance and dispersal are closely linked. If a male maintains social dominance for a long time, there is a risk of his producing offspring with his resident maturing daughters in the group. Accordingly, females commonly disperse in taxa — such as gorillas — in which the tenure of a dominant male normally exceeds the time needed for females to mature. In many other primate species, females can stay in their natal groups because male tenure is typically short. However, male tenure can vary greatly, and genetic analysis was used to show that even when a dominant white-handed capuchin male enjoys an unusually long spell of reproductive monopolization, he very rarely sires the offspring of his mature adult daughters (Muniz et al. 2006). The means by which inbreeding is avoided is puzzling, and raises the question of how primate individuals may recognize kin (Widdig 2007).
Social Relationships and Kin Preferences
Social relationships with members of the same sex are extraordinarily important for group-living primates. This has been particularly well demonstrated in baboons, where philopatric females with strong social bonds to other females —who are often matrilineal kin — live longer and produce more offspring (Silk et al. 2003). While it is easy to imagine that philopatric females become bonded with their sisters through their common relationship with the mother, it is less easy to understand how primates might recognize paternal kin. Nevertheless, studies in baboons and macaques have combined genetic determination of paternal relationships with observational data to suggest that females do indeed exhibit preferences for paternal kin, although not as strong as for maternal kin (Langergraber, 2012). In contrast, philopatric male chimpanzees exhibit no preference for social interactions with paternal brothers, and although they typically form strong social bonds with maternal brothers when available, the majority of social bonds are formed between unrelated individuals (Langergraber et al. 2007) (Figure 4).

Figure 4: Male chimpanzees of the Ngogo community in Kibale National Park, Uganda, engaging in a bout of grooming.
© 2012 Nature Education Photo credits: John Mitani. All rights reserved.
Researchers have more recently turned attention to the nature of social bonds between members of the dispersing sex. While dispersing males are expected to compete fiercely for reproductive opportunities in their new groups, it was recently shown that unrelated Assamese macaque males that cooperated in coalitions were more likely to improve their rank and subsequently reproduce (Schülke et al. 2010). As expected, pairs of kin are exceedingly rare among post-dispersal female chimpanzees, but female chimpanzees nonetheless form enduring, highly differentiated social bonds with one another, again emphasizing the flexible nature of social bonding in our closest living relatives (Langergraber et al. 2009). Overall, a combination of genetic analysis and behavioural observation highlights that kin ties are important, but not necessary, for social bonding.
Future Directions
The long lifespan of many primate species means that researchers require a good deal of time and patience to amass interesting genetic and observational data. In addition, because social groups occur under different ecological conditions and vary in the number of individuals of different ages and each sex, it is important to obtain data from a number of sites in order to gain a broad understanding of the characteristics of a species. Genetic analysis of noninvasive samples is becoming common, and effectively allows for the monitoring of groups unhabituated to human observation, thus increasing the range of possible analyses. The incipient use of next generation sequencing approaches, particularly if these can be extended to the use of DNA from noninvasive samples, will allow the analysis of vastly larger amounts of genetic data and increase the scope of possible research questions (Perry et al. 2010). Unfortunately, a constraint on future research is the severely fragmented distribution and decreasing number of many wild primates.
References and Recommended Reading
Di Fiore, A. Molecular genetic approaches to the study of primate behavior, social organization, and reproduction. American Journal of Physical Anthropology 122, 62-99 (2003).
Fernandez-Duque, E., Di Fiore, A., Huck, M. "The behavior, ecology and social evolution of New World monkeys," in The Evolution of Primate Societies, eds. J. Mitani et al. (Chicago, IL: University of Chicago Press, 2012).
Ellis, L. Dominance and reproductive success among nonhuman animals: A cross-species comparison. Ethology and Sociobiology 16, 257-333 (1995).
Greenwood, P. J. Mating systems, philopatry and dispersal in birds and mammals. Animal Behaviour 28, 1140-1162 (1980).
Guschanski K. et al. Females shape the genetic structure of a gorilla population. Current Biology 18, 1809-1814 (2008).
Kappeler, P. M. "The behavioral ecology of strepsirrhines and tarsiers," in The Evolution of Primate Societies, eds. J. Mitani et al. (Chicago, IL: University of Chicago Press, 2012).
Langergraber, K. E., Mitani, J. C. & Vigilant, L. The limited impact of kinship on cooperation in wild chimpanzees. Proceedings of the National Academy of Sciences of the United States of America 104, 7786-7790 (2007).
Langergraber, K. E. et al. Kinship and social bonds in female chimpanzees (Pan troglodytes). American Journal of Primatology 71, 1-12 (2009).
Langergraber, K. E. "Cooperation among kin," in The Evolution of Primate Societies, eds. J. Mitani et al. (Chicago, IL: University of Chicago Press, 2012).










