
Key Points
-
Lipids act as extracellular and intracellular messengers to control cell fate in normal physiology and disease. When deregulated, lipid signalling contributes to inflammation and cancer, metabolic, cardiovascular and degenerative disease.
-
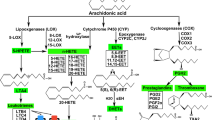
Signals such as growth factors, cytokines and chemokines, but also constituents of nutrients, modulate the activity of lipid-modifying enzymes: phosphoinositide 3-kinase (PI3K), sphingosine kinase (SphK), phospholipase C (PLC) and PLD function upstream of the activation of phospholipase A2 (PLA2), prostaglandin H2 synthase (PGH2S) and 5-lipoxygenase (5-LO), which are required for eicosanoid release.
-
Eicosanoids such as prostaglandins and leukotrienes have been known to have a role in inflammation. Recently, their extracellular action on G protein-coupled receptors (GPCRs) has been deciphered, and intracellular binding partners of eicosanoids have also been identified, which opens new possibilities for more selective drug-targeting strategies in inflammation and cancer.
-
Inflammation and cancer are both characterized by excess activation of PI3K and SphK pathways, which enforces growth-factor-receptor signalling, cell growth and survival, cell motility and degranulation. This overrides the pro-apoptotic actions of ceramide and sphingosine.
-
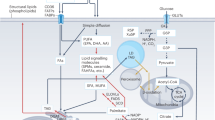
Although the nutritional energy supply normally increases insulin and lipid signalling through PI3K, excess circulating fatty acids induce long-term insulin resistance and function as pro-inflammatory agents. This occurs through signalling via the Toll-like receptor TLR4 and through endoplasmic reticulum stress promoted by intracellular lipid accumulation. It results in the production of reactive oxygen species and the activation of inflammatory kinase pathways (PKC, IKKβ), terminating the relay of signals through insulin receptor substrate (IRS).
-
Lipid-modifying enzymes, pathways and their downstream targets, including nuclear receptors and lipid-binding proteins, form a complex signalling network. Although many of these lipid mediators emerge from the same membranes, they are usually studied in isolation. Here, we present an integrated overview of lipid signalling in disease and highlight nodes of lipid signalling pathway interaction, and discuss emerging strategies for therapeutic interventions.
Abstract
Signalling lipids such as eicosanoids, phosphoinositides, sphingolipids and fatty acids control important cellular processes, including cell proliferation, apoptosis, metabolism and migration. Extracellular signals from cytokines, growth factors and nutrients control the activity of a key set of lipid-modifying enzymes: phospholipases, prostaglandin synthase, 5-lipoxygenase, phosphoinositide 3-kinase, sphingosine kinase and sphingomyelinase. These enzymes and their downstream targets constitute a complex lipid signalling network with multiple nodes of interaction and cross-regulation. Imbalances in this network contribute to the pathogenesis of human disease. Although the function of a particular signalling lipid is traditionally studied in isolation, this review attempts a more integrated overview of the key role of these signalling lipids in inflammation, cancer and metabolic disease, and discusses emerging strategies for therapeutic intervention.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

References
Feldberg, W., Holden, H. F. & Kellaway, C. H. The formation of lysocithin and of a muscle-stimulating substance by snake venoms. J. Physiol. 94, 232–248 (1938).
von Euler, U. S. On the specific vaso-dilating and plain muscle stimulating substances from accessory genital glands in man and certain animals (prostaglandin and vesiglandin). J. Physiol. 88, 213–234 (1936).
Boyce, J. A. Mast cells and eicosanoid mediators: a system of reciprocal paracrine and autocrine regulation. Immunol. Rev. 217, 168–185 (2007).
Mackay, H. J. & Twelves, C. J. Targeting the protein kinase C family: are we there yet? Nature Rev. Cancer 7, 554–562 (2007).
Wymann, M. P., Zvelebil, M. & Laffargue, M. Phosphoinositide 3-kinase signalling — which way to target? Trends Pharmacol. Sci. 24, 366–376 (2003).
Engelman, J. A., Luo, J. & Cantley, L. C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nature Rev. Genet. 7, 606–619 (2006).
Di Paolo, G. & De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 443, 651–657 (2006).
Taha, T. A., Mullen, T. D. & Obeid, L. M. A house divided: ceramide, sphingosine, and sphingosine-1-phosphate in programmed cell death. Biochim. Biophys. Acta 1758, 2027–2036 (2006).
Futerman, A. H. & Hannun, Y. A. The complex life of simple sphingolipids. EMBO Rep. 5, 777–782 (2004).
Rosen, H. & Goetzl, E. J. Sphingosine 1-phosphate and its receptors: an autocrine and paracrine network. Nature Rev. Immunol. 5, 560–570 (2005).
Cyster, J. G. Chemokines, sphingosine-1-phosphate, and cell migration in secondary lymphoid organs. Annu. Rev. Immunol. 23, 127–159 (2005).
Ishii, I., Fukushima, N., Ye, X. & Chun, J. Lysophospholipid receptors: signaling and biology. Annu. Rev. Biochem. 73, 321–354 (2004).
Meyer zu Heringdorf, D. & Jakobs, K. H. Lysophospholipid receptors: signalling, pharmacology and regulation by lysophospholipid metabolism. Biochim. Biophys. Acta 1768, 923–940 (2007).
Beaven, S. W. & Tontonoz, P. Nuclear receptors in lipid metabolism: targeting the heart of dyslipidemia. Annu. Rev. Med. 57, 313–329 (2006).
Semple, R. K., Chatterjee, V. K. & O'Rahilly, S. PPARγ and human metabolic disease. J. Clin. Invest. 116, 581–589 (2006).
Rivera, J. & Gilfillan, A. M. Molecular regulation of mast cell activation. J. Allergy Clin. Immunol. 117, 1214–1225; quiz 1226 (2006).
Ali, K. et al. Essential role for the p110δ phosphoinositide 3-kinase in the allergic response. Nature 431, 1007–1011 (2004). Reports a role for PI3Kδ in IgE-mediated mast cell activation and c-Kit signalling. Because mast cell differentiation requires full c-Kit function, PI3Kδ exerts an effect on mast cell numbers in certain tissues.
Okkenhaug, K., Ali, K. & Vanhaesebroeck, B. Antigen receptor signalling: a distinctive role for the p110δ isoform of PI3K. Trends Immunol. 28, 80–87 (2007).
Rivera, J. & Olivera, A. Src family kinases and lipid mediators in control of allergic inflammation. Immunol. Rev. 217, 255–268 (2007).
Choi, O. H., Kim, J. H. & Kinet, J. P. Calcium mobilization via sphingosine kinase in signalling by the FcɛRI antigen receptor. Nature 380, 634–636 (1996).
Young, K. W. & Nahorski, S. R. Sphingosine 1-phosphate: a Ca2+ release mediator in the balance. Cell Calcium 32, 335–341 (2002).
Subramanian, P. et al. Ceramide 1-phosphate acts as a positive allosteric activator of group IVA cytosolic phospholipase A2 α and enhances the interaction of the enzyme with phosphatidylcholine. J. Biol. Chem. 280, 17601–17607 (2005).
Chalfant, C. E. & Spiegel, S. Sphingosine 1-phosphate and ceramide 1-phosphate: expanding roles in cell signaling. J. Cell Sci. 118, 4605–4612 (2005).
Ghosh, M., Tucker, D. E., Burchett, S. A. & Leslie, C. C. Properties of the group IV phospholipase A2 family. Prog. Lipid Res. 45, 487–510 (2006).
Yedgar, S., Krimsky, M., Cohen, Y. & Flower, R. J. Treatment of inflammatory diseases by selective eicosanoid inhibition: a double-edged sword? Trends Pharmacol. Sci. 28, 459–464 (2007).
Kostenis, E. A glance at G-protein-coupled receptors for lipid mediators: a growing receptor family with remarkably diverse ligands. Pharmacol. Ther. 102, 243–257 (2004).
Brinkmann, V. Sphingosine 1-phosphate receptors in health and disease: Mechanistic insights from gene deletion studies and reverse pharmacology. Pharmacol. Ther. 115, 84–105 (2007).
Di Cristofano, A. & Pandolfi, P. P. The multiple roles of PTEN in tumor suppression. Cell 100, 387–390 (2000).
Rossi, D. J. & Weissman, I. L. Pten, tumorigenesis, and stem cell self-renewal. Cell 125, 229–231 (2006).
Wymann, M. P. & Marone, R. Phosphoinositide 3-kinase in disease: timing, location, and scaffolding. Curr. Opin. Cell Biol. 17, 141–149 (2005).
Maehama, T. & Dixon, J. E. The tumor suppressor, PTEN/MMAC1, dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-trisphosphate. J. Biol. Chem. 273, 13375–13378 (1998).
Ali, I. U., Schriml, L. M. & Dean, M. Mutational spectra of PTEN/MMAC1 gene: a tumor suppressor with lipid phosphatase activity. J. Natl Cancer Inst. 91, 1922–1932 (1999).
Samuels, Y. et al. High frequency of mutations of the PIK3CA gene in human cancers. Science 304, 554 (2004). Along with reference 37, introduces the concept of constitutive activation of the PI3K pathway through mutations in PI3Kα or the PH domain of PKB/Akt in human tumours.
Miled, N. et al. Mechanism of two classes of cancer mutations in the phosphoinositide 3-kinase catalytic subunit. Science 317, 239–242 (2007). The co-crystallized fragments of the p85 regulatory and the p110 catalytic subunits of PI3Kα reveal the mechanism for how mutations in PI3Kα that occur in human tumors cause constitutive activation of the p85–p110α heterodimer.
Brazil, D. P., Park, J. & Hemmings, B. A. PKB binding proteins. Getting in on the Akt. Cell 111, 293–303 (2002).
Manning, B. D. & Cantley, L. C. AKT/PKB signaling: navigating downstream. Cell 129, 1261–1274 (2007).
Carpten, J. D. et al. A transforming mutation in the pleckstrin homology domain of AKT1 in cancer. Nature 448, 439–444 (2007). References 33 and 37 introduce the concept of constitutive activation of the PI3K pathway through mutations in PI3Kα or the PH domain of PKB/Akt in human tumours.
Guertin, D. A. & Sabatini, D. M. Defining the role of mTOR in cancer. Cancer Cell 12, 9–22 (2007).
Fritz, G. & Kaina, B. Rho GTPases: promising cellular targets for novel anticancer drugs. Curr. Cancer Drug Targets. 6, 1–14 (2006).
Miyaji, M. et al. Role of membrane sphingomyelin and ceramide in platform formation for Fas-mediated apoptosis. J. Exp. Med. 202, 249–259 (2005).
Gomez-Munoz, A. Ceramide 1-phosphate/ceramide, a switch between life and death. Biochim. Biophys. Acta 1758, 2049–2056 (2006).
Suzuki, E., Handa, K., Toledo, M. S. & Hakomori, S. Sphingosine-dependent apoptosis: a unified concept based on multiple mechanisms operating in concert. Proc. Natl Acad. Sci. USA 101, 14788–14793 (2004).
Gomez-Munoz, A. et al. Ceramide-1-phosphate promotes cell survival through activation of the phosphatidylinositol 3-kinase/protein kinase B pathway. FEBS Lett. 579, 3744–3750 (2005).
Semenkovich, C. F. Insulin resistance and atherosclerosis. J. Clin. Invest. 116, 1813–1822 (2006).
Taniguchi, C. M., Emanuelli, B. & Kahn, C. R. Critical nodes in signalling pathways: insights into insulin action. Nature Rev. Mol. Cell. Biol. 7, 85–96 (2006).
White, M. F. IRS proteins and the common path to diabetes. Am. J. Physiol. Endocrinol. Metab. 283, E413–E422 (2002).
Um, S. H. et al. Absence of S6K1 protects against age- and diet-induced obesity while enhancing insulin sensitivity. Nature 431, 200–205 (2004). Mice that are deficient in S6 kinase-1, an effector of the mammalian target of rapamycin (mTOR), are protected against obesity owing to enhanced β-oxidation. On a high-fat diet, these mice remain insulin sensitive owing to the loss of feedback inhibition of IRS by S6K1.
Um, S. H., D'Alessio, D. & Thomas, G. Nutrient overload, insulin resistance, and ribosomal protein S6 kinase 1, S6K1. Cell Metab. 3, 393–402 (2006).
Boden, G. et al. Free fatty acids produce insulin resistance and activate the proinflammatory nuclear factor-κB pathway in rat liver. Diabetes 54, 3458–3465 (2005).
Makowski, L. et al. Lack of macrophage fatty-acid-binding protein aP2 protects mice deficient in apolipoprotein E against atherosclerosis. Nature Med. 7, 699–705 (2001).
Rosen, E. D. & Spiegelman, B. M. PPARγ: a nuclear regulator of metabolism, differentiation, and cell growth. J. Biol. Chem. 276, 37731–37734 (2001).
Hotamisligil, G. S. et al. Uncoupling of obesity from insulin resistance through a targeted mutation in aP2, the adipocyte fatty acid binding protein. Science 274, 1377–1379 (1996).
Maeda, K. et al. Adipocyte/macrophage fatty acid binding proteins control integrated metabolic responses in obesity and diabetes. Cell Metab. 1, 107–119 (2005).
Furuhashi, M. et al. Treatment of diabetes and atherosclerosis by inhibiting fatty-acid-binding protein aP2. Nature 447, 959–965 (2007). Describes an orally active small-molecule inhibitor of FABP4 as an effective therapeutic agent against severe atherosclerosis and type 2 diabetes in mouse models. These results suggest that FABP4 inhibitors may represent a new class of powerful therapeutic agents to prevent and treat metabolic diseases.
Summers, S. A. Ceramides in insulin resistance and lipotoxicity. Prog. Lipid Res. 45, 42–72 (2006).
Wei, Y., Wang, D., Topczewski, F. & Pagliassotti, M. J. Saturated fatty acids induce endoplasmic reticulum stress and apoptosis independently of ceramide in liver cells. Am. J. Physiol. Endocrinol. Metab. 291, E275–E281 (2006).
Li, Y. et al. Enrichment of endoplasmic reticulum with cholesterol inhibits sarcoplasmic–endoplasmic reticulum calcium ATPase-2b activity in parallel with increased order of membrane lipids: implications for depletion of endoplasmic reticulum calcium stores and apoptosis in cholesterol-loaded macrophages. J. Biol. Chem. 279, 37030–37039 (2004).
Feng, B. et al. The endoplasmic reticulum is the site of cholesterol-induced cytotoxicity in macrophages. Nature Cell Biol. 5, 781–792 (2003).
Ron, D. & Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nature Rev. Mol. Cell. Biol. 8, 519–529 (2007).
Ozcan, U. et al. Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science 313, 1137–1140 (2006). Together with reference 85, shows that obesity causes ER stress, which in turn leads to hyperactivation of JNK, inhibitory phosphorylation of IRS and suppression of insulin receptor signalling. Chemical chaperones that enhance the adaptive capacity of the ER act as potent antidiabetic modalities with a potential application in the treatment of type 2 diabetes.
Cullinan, S. B. & Diehl, J. A. Coordination of ER and oxidative stress signaling: the PERK/Nrf2 signaling pathway. Int. J. Biochem. Cell Biol. 38, 317–332 (2006).
Xue, X. et al. Tumor necrosis factor α (TNFα) induces the unfolded protein response (UPR) in a reactive oxygen species (ROS)-dependent fashion, and the UPR counteracts ROS accumulation by TNFα. J. Biol. Chem. 280, 33917–33925 (2005).
Furukawa, S. et al. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest. 114, 1752–1761 (2004).
Houstis, N., Rosen, E. D. & Lander, E. S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 440, 944–948 (2006). Indicates that antioxidants that lower ROS levels may be useful to treat type 2 diabetes and other insulin-resistant states.
Lowell, B. B. & Shulman, G. I. Mitochondrial dysfunction and type 2 diabetes. Science 307, 384–387 (2005).
Hotamisligil, G. S. Inflammation and metabolic disorders. Nature 444, 860–867 (2006).
Weisberg, S. P. et al. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Invest. 112, 1796–1808 (2003).
Xu, H. et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 112, 1821–1830 (2003).
Shoelson, S. E., Lee, J. & Goldfine, A. B. Inflammation and insulin resistance. J. Clin. Invest. 116, 1793–1801 (2006).
Hotamisligil, G. S., Shargill, N. S. & Spiegelman, B. M. Adipose expression of tumor necrosis factor-α: direct role in obesity-linked insulin resistance. Science 259, 87–91 (1993).
Uysal, K. T., Wiesbrock, S. M., Marino, M. W. & Hotamisligil, G. S. Protection from obesity-induced insulin resistance in mice lacking TNF-α function. Nature 389, 610–614 (1997).
Ventre, J. et al. Targeted disruption of the tumor necrosis factor-α gene: metabolic consequences in obese and nonobese mice. Diabetes 46, 1526–1531 (1997).
Krogh-Madsen, R., Plomgaard, P., Moller, K., Mittendorfer, B. & Pedersen, B. K. Influence of TNF-α and IL-6 infusions on insulin sensitivity and expression of IL-18 in humans. Am. J. Physiol. Endocrinol. Metab. 291, E108–E114 (2006).
Beutler, B. Innate immunity: an overview. Mol. Immunol. 40, 845–859 (2004).
Lee, J. Y., Sohn, K. H., Rhee, S. H. & Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. J. Biol. Chem. 276, 16683–16689 (2001).
Shi, H. et al. TLR4 links innate immunity and fatty acid-induced insulin resistance. J. Clin. Invest. 116, 3015–3025 (2006).
Yu, C. et al. Mechanism by which fatty acids inhibit insulin activation of insulin receptor substrate-1 (IRS-1)-associated phosphatidylinositol 3-kinase activity in muscle. J. Biol. Chem. 277, 50230–50236 (2002).
Chawla, A., Repa, J. J., Evans, R. M. & Mangelsdorf, D. J. Nuclear receptors and lipid physiology: opening the X-files. Science 294, 1866–1870 (2001).
Glass, C. K. & Ogawa, S. Combinatorial roles of nuclear receptors in inflammation and immunity. Nature Rev. Immunol. 6, 44–55 (2006).
Aguirre, V., Uchida, T., Yenush, L., Davis, R. & White, M. F. The c-Jun NH2-terminal kinase promotes insulin resistance during association with insulin receptor substrate-1 and phosphorylation of Ser(307). J. Biol. Chem. 275, 9047–9054 (2000).
Gao, Z. et al. Serine phosphorylation of insulin receptor substrate 1 by inhibitor κB kinase complex. J. Biol. Chem. 277, 48115–48121 (2002).
Griffin, M. E. et al. Free fatty acid-induced insulin resistance is associated with activation of protein kinase Cθ and alterations in the insulin signaling cascade. Diabetes 48, 1270–1274 (1999).
Hotamisligil, G. S. et al. IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-α- and obesity-induced insulin resistance. Science 271, 665–668 (1996).
Zick, Y. Ser/Thr phosphorylation of IRS proteins: a molecular basis for insulin resistance. Sci. STKE 2005, pe4 (2005).
Ozcan, U. et al. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 306, 457–461 (2004). Together with reference 60, shows that obesity causes ER stress, which in turn leads to hyperactivation of JNK, inhibitory phosphorylation of IRS and suppression of insulin receptor signalling. Chemical chaperones that enhance the adaptive capacity of the ER act as potent antidiabetic modalities with a potential application in the treatment of type 2 diabetes.
Hirosumi, J. et al. A central role for JNK in obesity and insulin resistance. Nature 420, 333–336 (2002).
Ricci, R. et al. Requirement of JNK2 for scavenger receptor A-mediated foam cell formation in atherogenesis. Science 306, 1558–1561 (2004).
Kaneto, H. et al. Possible novel therapy for diabetes with cell-permeable JNK-inhibitory peptide. Nature Med. 10, 1128–1132 (2004).
Liu, G. & Rondinone, C. M. JNK: bridging the insulin signaling and inflammatory pathway. Curr. Opin. Investig. Drugs 6, 979–987 (2005).
Yuan, M. et al. Reversal of obesity- and diet-induced insulin resistance with salicylates or targeted disruption of IKKβ. Science 293, 1673–1677 (2001).
Kim, J. K. et al. PKC-θ knockout mice are protected from fat-induced insulin resistance. J. Clin. Invest. 114, 823–827 (2004).
Baud, V. & Karin, M. Signal transduction by tumor necrosis factor and its relatives. Trends Cell Biol. 11, 372–377 (2001).
Urano, F. et al. Coupling of stress in the ER to activation of JNK protein kinases by transmembrane protein kinase IRE1. Science 287, 664–666 (2000).
Deng, J. et al. Translational repression mediates activation of nuclear factor κB by phosphorylated translation initiation factor 2. Mol. Cell. Biol. 24, 10161–10168 (2004).
Hu, P., Han, Z., Couvillon, A. D., Kaufman, R. J. & Exton, J. H. Autocrine tumor necrosis factor α links endoplasmic reticulum stress to the membrane death receptor pathway through IRE1α-mediated NF-κB activation and down-regulation of TRAF2 expression. Mol. Cell. Biol. 26, 3071–3084 (2006).
Hirsch, E. et al. Central role for G protein-coupled phosphoinositide 3-kinase γ in inflammation. Science 287, 1049–1053 (2000).
Li, Z. et al. Roles of PLC-β2 and -β3 and PI3Kγ in chemoattractant-mediated signal transduction. Science 287, 1046–1049 (2000).
Sasaki, T. et al. Function of PI3Kγ in thymocyte development, T cell activation, and neutrophil migration. Science 287, 1040–1046 (2000).
Ruckle, T., Schwarz, M. K. & Rommel, C. PI3Kγ inhibition: towards an 'aspirin of the 21st century'? Nature Rev. Drug Discov. 5, 903–918 (2006).
Camps, M. et al. Blockade of PI3Kγ suppresses joint inflammation and damage in mouse models of rheumatoid arthritis. Nature Med. 11, 936–943 (2005).
Barber, D. F. et al. Class IB-phosphatidylinositol 3-kinase (PI3K) deficiency ameliorates IA-PI3K-induced systemic lupus but not T cell invasion. J. Immunol. 176, 589–593 (2006). References 100 and 101 validated the pharmacological targeting of PI3Kγ to treat rheumatoid arthritis and lupus using preventive and semi-therapeutic protocols in mouse models.
Lee, K. S., Lee, H. K., Hayflick, J. S., Lee, Y. C. & Puri, K. D. Inhibition of phosphoinositide 3-kinase δ attenuates allergic airway inflammation and hyperresponsiveness in murine asthma model. FASEB J. 20, 455–465 (2006).
Laffargue, M. et al. Phosphoinositide 3-kinase γ is an essential amplifier of mast cell function. Immunity 16, 441–451 (2002). Shows that PI3Kγ has an essential role in IgE-mediated mast cell activation and passive systemic anaphylaxis in vivo . IgE–antigen complexes initially trigger a weak activation of mast cells. This causes a release of adenosine, which acts in an autocrine or paracrine manner to activate trimeric G proteins and PI3Kγ.
Patrucco, E. et al. PI3Kγ modulates the cardiac response to chronic pressure overload by distinct kinase-dependent and -independent effects. Cell 118, 375–387 (2004).
Doukas, J. et al. Phosphoinositide 3-kinase γ/δ inhibition limits infarct size after myocardial ischemia/reperfusion injury. Proc. Natl Acad. Sci. USA 103, 19866–19871 (2006).
Kappos, L. et al. Oral fingolimod (FTY720) for relapsing multiple sclerosis. N. Engl. J. Med. 355, 1124–1140 (2006).
Massberg, S. & von Andrian, U. H. Fingolimod and sphingosine-1-phosphate — modifiers of lymphocyte migration. N. Engl. J. Med. 355, 1088–1091 (2006).
Miller, D. Multiple sclerosis: new insights and therapeutic progress. Lancet Neurol. 6, 5–6 (2007).
Baumruker, T., Billich, A. & Brinkmann, V. FTY720, an immunomodulatory sphingolipid mimetic: translation of a novel mechanism into clinical benefit in multiple sclerosis. Expert Opin. Investig. Drugs 16, 283–289 (2007).
Wei, S. H. et al. Sphingosine 1-phosphate type 1 receptor agonism inhibits transendothelial migration of medullary T cells to lymphatic sinuses. Nature Immunol. 6, 1228–1235 (2005).
Sanna, M. G. et al. Enhancement of capillary leakage and restoration of lymphocyte egress by a chiral S1P1 antagonist in vivo. Nature Chem. Biol. 2, 434–441 (2006).
Rosen, H., Sanna, M. G., Cahalan, S. M. & Gonzalez-Cabrera, P. J. Tipping the gatekeeper: S1P regulation of endothelial barrier function. Trends Immunol. 28, 102–107 (2007).
Reid, R. C. Inhibitors of secretory phospholipase A2 group IIA. Curr. Med. Chem. 12, 3011–3026 (2005).
Juni, P. et al. Risk of cardiovascular events and rofecoxib: cumulative meta-analysis. Lancet 364, 2021–2029 (2004). Reports on the withdrawal of the cyclooxygenase-2 inhibitor rofecoxib from the market owing to adverse cardiovascular effects.
Pettipher, R., Hansel, T. T. & Armer, R. Antagonism of the prostaglandin D2 receptors DP1 and CRTH2 as an approach to treat allergic diseases. Nature Rev. Drug Discov. 6, 313–325 (2007).
Steinke, J. W. & Culp, J. A. Leukotriene synthesis inhibitors versus antagonists: the pros and cons. Curr. Allergy Asthma Rep. 7, 126–133 (2007).
Peters-Golden, M. & Henderson, W. R. J. Leukotrienes. N. Engl. J. Med. 357, 1841–1854 (2007).
Neshat, M. S. et al. Enhanced sensitivity of PTEN-deficient tumors to inhibition of FRAP/mTOR. Proc. Natl Acad. Sci. USA 98, 10314–10319 (2001).
Fan, Q. W. et al. A dual PI3 kinase/mTOR inhibitor reveals emergent efficacy in glioma. Cancer Cell 9, 341–349 (2006).
Raynaud, F. I. et al. Pharmacologic characterization of a potent inhibitor of class I phosphatidylinositide 3-kinases. Cancer Res. 67, 5840–5850 (2007).
Yaguchi, S. et al. Antitumor activity of ZSTK474, a new phosphatidylinositol 3-kinase inhibitor. J. Natl Cancer Inst. 98, 545–556 (2006).
Stauffer, F. et al. Biochemical, cellular, and in vivo profiling of a new PI3K inhibitor from the imidazoquinoline series [abstract]. In American Association for Cancer Research Annual Meeting: Proceedings, 14–18 Apr 2007, Los Angeles, California. Abstract no. 269.
Nagata, Y. et al. PTEN activation contributes to tumor inhibition by trastuzumab, and loss of PTEN predicts trastuzumab resistance in patients. Cancer Cell 6, 117–127 (2004).
Visentin, B. et al. Validation of an anti-sphingosine-1-phosphate antibody as a potential therapeutic in reducing growth, invasion, and angiogenesis in multiple tumor lineages. Cancer Cell 9, 225–238 (2006). Highlights the therapeutic potential of anti-S1P antibodies in a mouse cancer model.
Milstien, S. & Spiegel, S. Targeting sphingosine-1-phosphate: a novel avenue for cancer therapeutics. Cancer Cell 9, 148–150 (2006).
French, K. J. et al. Antitumor activity of sphingosine kinase inhibitors. J. Pharmacol. Exp. Ther. 318, 596–603 (2006).
Bandhuvula, P. & Saba, J. D. Sphingosine-1-phosphate lyase in immunity and cancer: silencing the siren. Trends Mol. Med. 13, 210–217 (2007).
Oskouian, B. et al. Sphingosine-1-phosphate lyase potentiates apoptosis via p53- and p38-dependent pathways and is down-regulated in colon cancer. Proc. Natl Acad. Sci. USA 103, 17384–17389 (2006).
Neviani, P. et al. FTY720, a new alternative for treating blast crisis chronic myelogenous leukemia and Philadelphia chromosome-positive acute lymphocytic leukemia. J. Clin. Invest. 117, 2408–2421 (2007).
Skoura, A. et al. Essential role of sphingosine 1-phosphate receptor 2 in pathological angiogenesis of the mouse retina. J. Clin. Invest. 117, 2506–2516 (2007).
Ying, L. & Hofseth, L. J. An emerging role for endothelial nitric oxide synthase in chronic inflammation and cancer. Cancer Res. 67, 1407–1410 (2007).
Nissen, S. E. & Wolski, K. Effect of rosiglitazone on the risk of myocardial infarction and death from cardiovascular causes. N. Engl. J. Med. 356, 2457–2471 (2007).
Ishii, H. et al. Amelioration of vascular dysfunctions in diabetic rats by an oral PKCβ inhibitor. Science 272, 728–731 (1996).
Aiello, L. P. et al. Inhibition of PKCβ by oral administration of ruboxistaurin is well tolerated and ameliorates diabetes-induced retinal hemodynamic abnormalities in patients. Invest. Ophthalmol. Vis. Sci. 47, 86–92 (2006).
Bansode, R., Huang, W., Roy, S., Mehta, M. & Mehta, K. D. Protein kinase C deficiency increases fatty acid oxidation and reduces fat storage. J. Biol. Chem. 283, 231–236 (2008).
Balla, T. & Varnai, P. Visualizing cellular phosphoinositide pools with GFP-fused protein-modules. Sci. STKE 2002, pl3 (2002).
Funk, C. D. Prostaglandins and leukotrienes: advances in eicosanoid biology. Science 294, 1871–1875 (2001).
Serhan, C. N. Resolution phase of inflammation: novel endogenous anti-inflammatory and proresolving lipid mediators and pathways. Annu. Rev. Immunol. 25, 101–137 (2007).
Cantley, L. C. The phosphoinositide 3-kinase pathway. Science 296, 1655–1657 (2002).
Clarke, C. J. & Hannun, Y. A. Neutral sphingomyelinases and nSMase2: bridging the gaps. Biochim. Biophys. Acta 1758, 1893–1901 (2006).
Morales, A., Lee, H., Goni, F. M., Kolesnick, R. & Fernandez-Checa, J. C. Sphingolipids and cell death. Apoptosis 12, 923–939 (2007).
Hait, N. C., Oskeritzian, C. A., Paugh, S. W., Milstien, S. & Spiegel, S. Sphingosine kinases, sphingosine 1-phosphate, apoptosis and diseases. Biochim. Biophys. Acta 1758, 2016–2026 (2006).
Mangelsdorf, D. J. et al. The nuclear receptor superfamily: the second decade. Cell 83, 835–839 (1995).
Berger, J. & Moller, D. E. The mechanisms of action of PPARs. Annu. Rev. Med. 53, 409–435 (2002).
Morris, A. J. Regulation of phospholipase D activity, membrane targeting and intracellular trafficking by phosphoinositides. Biochem. Soc. Symp. 74, 247–257 (2007).
Lee, J. S. et al. Phosphatidylinositol (3,4,5)-trisphosphate specifically interacts with the phox homology domain of phospholipase D1 and stimulates its activity. J. Cell Sci. 118, 4405–4413 (2005).
Pitson, S. M. et al. Phosphorylation-dependent translocation of sphingosine kinase to the plasma membrane drives its oncogenic signalling. J. Exp. Med. 201, 49–54 (2005).
Acknowledgements
We apologize for not citing many original publications due to space restrictions. We thank P. Fotiadou and R. Marone for corrections and discussions. M.P.W. is supported by grants from Oncosuisse, the Krebsliga beider Basel, the Swiss National Science Foundation and the EU FP6 programme. R.S. is supported by grants from the Swiss National Science Foundation and Novartis Stiftung.
Author information
Authors and Affiliations
Corresponding author
Related links
Related links
DATABASES
UniProtKB
FURTHER INFORMATION
Lipid Metabolites and Pathways Strategy
Novartis Oncology: Targeted Kinase Inhibitors – BEZ235
POSTER
Glossary
- G protein-coupled receptor
-
(GPCR). A seven-transmembrane helix cell-surface receptor that signals through heterotrimeric GTP-binding proteins. GPCRs trigger the dissociation of trimeric G proteins into Gα and Gβγ subunits by facilitating loading of GTP onto Gα. Both Gα and Gβγ subunits interact with downstream effector proteins. Relay of GPCR signals terminates when GTP is hydrolysed by the intrinsic GTPase activity of Gα.
- Lysolipid
-
An asymmetrical lipid that contains only one acyl chain.
- GTPase-activating protein
-
(GAP). One of a family of proteins that inactivate small GTP-binding proteins, such as Ras family members, by increasing their rate of GTP hydrolysis.
- Neutrophil
-
The most abundant phagocytic cell of the myeloid lineage, which has an important role in host defence and chronic inflammatory disease. It undergoes chemotaxis towards sites of infection, which involves GPCR signalling.
- Mast cell
-
A tissue-resident cell with large secretory granules that contain histamine and various inflammatory mediators. Mast cells are key in inflammation and IgE-mediated allergy.
- Granulocyte
-
A type of white blood cell (including neutrophils, eosinophils and basophils) that is dedicated to the release of granules.
- Lipid raft
-
A membrane microdomain that is enriched in cholesterol, sphingolipids and lipid-modified proteins such as GPI-linked proteins, as well as palmitoylated proteins. These microdomains often function as platforms for distinct signalling events.
- SH2 domain
-
(Src-homology-2 domain). A protein domain that recognizes and binds specific tyrosine-phosphorylated sequences, and thereby has a key role in the directed delivery of signal transduction effector proteins. The SH2 domains of the p85 regulatory subunit of PI3K, for example, specifically recognize phosphoTyr-X-X-Met motifs.
- PH domain
-
(pleckstrin homology domain). A sequence of ∼100 amino acids that is present in many signalling molecules and binds to lipid products of phosphoinositide kinases. PH domains can have individual specificities for polyphospho-inositides.
- Store-operated calcium channel
-
A plasma membrane ion channel of uncertain identity that opens in response to depletion of internal Ca2+ stores.
- Peroxisome
-
An organelle that contains the machinery to oxidize fatty acids. A site of H2O2 production and destruction.
- Statin
-
One of a class of cholesterol-lowering drugs that inhibit the key enzyme HMG-CoA reductase in the synthesis of cholesterol.
- Triterpenoid
-
A hydrocarbon that results from the condensation of 5 carbon isoprene units. Triterpenoids contain 6 isoprene units and 30 carbon atoms, as found in the steroid precursor squalene.
- Lipodystrophy
-
A disorder that features the loss of adipocytes.
- Acanthosis nigricans
-
A velvety light-brown to black pigmentation of the skin caused by insulin spill-over from excessive production due to obesity or insulin resistance, which results in abnormal growth.
- Basal lamina
-
A thin sheet of extracellular matrix molecules, which consists mainly of collagens, laminins, nidogen and proteoglycans, and forms a region between cells and adjacent connective tissue. It has an important influence on the organization of tissues.
- FOXO transcription factor
-
One of a family of >40 proteins that belong to the winged-helix class of DNA-binding proteins. They are involved in diverse cellular functions, including glucose metabolism, apoptosis and cell-cycle regulation.
- Guanine nucleotide-exchange factor
-
(GEF). A protein that facilitates the exchange of GDP for GTP in the nucleotide-binding pocket of a GTP-binding protein.
- Rho GTPase
-
A Ras-related small GTPase that functions as a molecular switch to control signal transduction pathways. Traditionally it is known as a regulator of actin polymerization.
- 14-3-3 protein
-
An adaptor or scaffold protein that forms homo- and heterodimers. It binds through specialized phosphorylated peptide motifs (mostly containing phosphoserine) to various proteins that are involved in signal transduction and cell-cycle control.
- Hepatosteatosis
-
Fat accumulation in liver cells, which gives rise to fatty liver disease.
- Innate immune system
-
Components of the immune system that constitute the immediate, first line of defence against antigens. Pathogen recognition is mediated by pattern-recognition receptors, such as Toll-like receptors.
- Salicylate
-
A group of compounds with anti-inflammatory properties. A prominent member is acetylsalicylate, also known as aspirin.
- Complement
-
Nine interacting serum proteins (C1–C9), many of them with proteolytic activity, that are activated in a coordinated way and participate in bacterial lysis and, as fragments, in leukocyte chemotaxis.
- Anaphylaxis
-
A severe, potentially fatal, allergic reaction that involves histamine release from mast cells. This triggers dilation of blood vessels, a drop in blood pressure and increased vascular permeability.
Rights and permissions
About this article
Cite this article
Wymann, M., Schneiter, R. Lipid signalling in disease. Nat Rev Mol Cell Biol 9, 162–176 (2008). https://doi.org/10.1038/nrm2335
Issue Date:
DOI: https://doi.org/10.1038/nrm2335
This article is cited by
-
Human Cerebrospinal Fluid Sample Preparation and Annotation for Integrated Lipidomics and Metabolomics Profiling Studies
Molecular Neurobiology (2024)
-
Lipidomic signatures in Colombian adults with metabolic syndrome
Journal of Diabetes & Metabolic Disorders (2024)
-
LC/MS-based untargeted lipidomics reveals lipid signatures of nonpuerperal mastitis
Lipids in Health and Disease (2023)
-
CD1-mediated immune responses in mucosal tissues: molecular mechanisms underlying lipid antigen presentation system
Experimental & Molecular Medicine (2023)
-
Ferroptosis in lung cancer: a novel pathway regulating cell death and a promising target for drug therapy
Cell Death Discovery (2023)
