
Abstract
Darwin used the metaphor of a ‘tangled bank’ to describe the complex interactions between species. Those interactions are varied: they can be antagonistic ones involving predation, herbivory and parasitism, or mutualistic ones, such as those involving the pollination of flowers by insects. Moreover, the metaphor hints that the interactions may be complex to the point of being impossible to understand. All interactions can be visualized as ecological networks, in which species are linked together, either directly or indirectly through intermediate species. Ecological networks, although complex, have well defined patterns that both illuminate the ecological mechanisms underlying them and promise a better understanding of the relationship between complexity and ecological stability.
Similar content being viewed by others


Main
A food web maps which species eat which other species1,2,3. Such trophic interactions—henceforth ‘links’—are typically antagonistic predator–prey ones, but we expand the discussion to include webs of mutualists4,5, including, for example, the interactions of flowers and their pollinators, and fruits and their seed dispersers.
Ecological networks are complex, with each species typically closely linked to all others, either directly or indirectly6,7. For example, links tend to be ‘nested’8,9—that is, the diet of the most specialized species is a subset of the diet of the next more generalized species, and its diet a subset of the next more generalized, and so on. The most generalized species may include most of the prey species present in its diet. Within this pattern of close links to other species, there are reasons to expect even denser clusters of links. For example, coevolution is expected to generate clustered links that stem from the reciprocal mutual specialization between plants and their pollinators or seed dispersers4,5. The general existence and underlying causes of dense clusters is, however, still a matter of debate3,10.
In this Review, we compare ecological networks to non-ecological networks, and consider their similarities, differences and underlying causes. Along with networks of interacting computers, genes or humans, ecological networks display well-defined, similar patterns of organization11,12,13. On close inspection, ecological networks are unlike other networks. Their assembly follows different rules, and the processes of predation, competition and mutualism constrain them in unique ways. Other networks nonetheless help us understand why ecological ones are special in the constraints that apply to them and how they develop.
Finally, there is a paradox. Theory predicts that complex ecological networks are likely to be ‘fragile’ in various ways. For example, complexity affects the chance that species can coexist at a stable equilibrium. It also affects resilience (how fast populations recover from disturbances), the variability of population densities over time and the persistence of community composition. It should also affect the resistance to change, when species invade or are driven to extinction14,15,16. Every species is closely linked to every other6,7, so—metaphorically—when a tree falls in a rainforest, every species in that species-rich, complex system would seem to ‘hear’ that event quickly. Every disturbance buffets every other species, so how do species persist in this ‘noisy’ world? And which species will persist in the now extensively modified and increasingly species-poor world we are creating for them? We address these questions in the penultimate section, where we highlight both the extent of our present understanding, and the important gaps in our knowledge.
Complexity
No species is distant from the most connected species, nor any top predator from a species at the base of the web2,6 (Fig. 1; Box 1 defines ‘distance’ and other technical terms). In seven species-rich webs, 80% and 97% of species are, respectively, within two or three links from each other7. Both the cascade model2, and the niche model17,18 it inspired, predict short distances between top predators and plants or detritivores at the base of a web19. The cascade model posits nested diets, with the top predator potentially exploiting all the other species, the next predator exploiting all but the top predator, and so on.

The Ythan estuary food web66,73. a, Node colour indicates the length of the shortest path linking the most connected species (the flounder Platichthys flesus, in red) and each other species from the network. (The trophic direction of the links—what eats what—is ignored). Dark green, species are one link apart; light green, two links; and blue, three links. The central circle represents the densest sub-web75, which consists of 28 species with at least 7 links with the rest of the species from the sub-web. This sub-web contributes most to the observed clustering. b, Food chains between basal species of Enteromorpha (red node at the bottom) and the top predator, the cormorant Phalacrocorax carbo (red node at the top). Links corresponding to the shortest path connecting them are in blue (2-links), and those corresponding to the longest food chain between these two species are in red (6-links).
Given that species in a web are typically closely connected, are there even denser patterns of links within each web? The first approach to this question considers detailed patterns, and can be applied to webs with few or many species. (Many early studies involved webs with few species. Box 2 considers the limitations on what data known food webs provide.)
Food webs straddling different habitats generate compartments corresponding to habitat boundaries20. But no agreement exists when looking for compartments within a habitat20. Some visual inspections for compartments find them21,22, while others do not23. Using statistics, one study24 finds compartments within the food web of an estuary. Another20 finds roughly equal numbers of more and less compartmented webs than expected by chance. A third study identified compartments in three of five species-rich food webs25.
Analysing clusters is more tractable than detecting compartments. In a random web with S species and an average of k links per species, clustering scales as k/S (refs 12, 13). Generally, webs with many species and few links per species are more clustered than are random counterparts6,10. Embedded in the most highly clustered webs, however, are two classes of systems long known3 and theoretically expected3 to have tight clusters of links. In aquatic systems, many fish are ‘life-history omnivores’ that feed from several trophic levels at various life stages—on phytoplankton after hatching to other piscivorous fish as adults—including those that ate them when young26 (see Fig. 1). Tight clusters are common in host–parasitoid systems, where a parasitoid may feed on a host, but also hyper-parasitize other parasitoids23,27.
In sum, important classes of webs involve densely linked groups of species. Life-history omnivores dominate the marine ecosystems that cover most of the planet (as well as many freshwater ecosystems), whereas plant–herbivore–parasitoid systems account for the great majority of Earth's species28. From these perspectives, densely clustered webs are nearer the norm, not exceptions. Moreover, compartments are likely to correspond to habitat boundaries, if only because of the difficulties of species feeding successfully in two or more habitats. This begs the question, not addressed here, of what constitutes a habitat for anything less subtle than major categories such as ‘ocean, lake, and land’20.
These exceptions noted, there is no clear consensus about compartments and clusters. Whether the ‘species’ that the food web depicts are actually species or taxonomically aggregated groups or ‘trophic species’—species that apparently share the same (or similar) predators and prey2,3—confounds the results. Krause et al.25 provide the most complete analysis, and show that the detection of compartments requires new algorithms. Increasing taxonomic resolution and details on the strength of interactions increased their chances of finding compartments. These complexities inevitably create controversy. Its resolution will come from applying their ideas to other quantified and taxonomically specific webs.
A second approach applies to only species-rich webs and considers P(k), the probability of a species having k links (Fig. 2). Power law, ‘scale-free’ distributions, P(k)∼k-γ (we use ∼ here to mean ‘follows a function’), describe the links between human social contacts and internet connections11,12,13. They suggest a ‘popularity principle’ whereby the ‘rich get richer’. As these non-ecological networks grow, nodes are likely to get more links the more they have already12, and dense clusters develop. In these systems, the constant γ is typically a value between 2 and 3 (refs 12, 13).

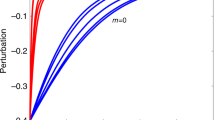
a–e, The cumulative probabilities Pc(k), for ≥k, where P(k) is the probability a species has k links to other species, and is given by P(k)∼k-γe-k/γ where e-k/γ introduces a cut-off at some characteristic scale γ. Panels a (log–log) and b (log–linear) show three different modelled networks. Black lines, single-scale networks; when γ is very small, the distribution has a fast decaying tail, typically exponential, P(k)∼e-k/γ. Green lines, truncated scale-free networks; these correspond to intermediate values of γ where the distribution has a power law regime followed by a sharp cut-off, with an exponential decay of the tail. Red lines, scale-free networks; for large values of γ the number of connections per species decays as a power law, P(k)∼k-γ, a function with a relatively ‘fat tail’. c–e, Experimental data (filled circles) and best fits (lines); c, a frugivore–plant web76; d, a pollinator–plant web77; and e, the food web from El Verde rainforest78. In c and d, for red circles, k is the number of plants species visited by an animal, and for black circles, k is the number of pollinator species visiting each plant species. In e, we sum prey–predator links and predator–prey links for each species. Best fits to the data in c–e are as follows: c, animals, exponential, P(k) = e-k/3.998; plants, truncated power law, P(k) = k-0.013e-k/11.22; d, animals, power law, P(k) = k-1.512; plants, truncated power law, P(k) = k-0.2822e-k/42.55; e, exponential, P(k) = e-k/8.861. f–h, Photographs of a frugivore–plant (f), insect–flower (g) and predator–prey (h) interaction of webs depicted in c–e, respectively.
Alternatively, many factors constrain a predator's diet, so truncating the linkage distribution, preventing the ‘rich from getting richer’ and suggesting an exponential trimming of linkages: P(k)∼e-k/ξ. How large is the truncation measured by the parameter ξ ? One can fit the combined model P(k)∼k-γe-k/ξ (refs 29, 30). Intuitively, the value ξ measures the number of links below which power laws describe the systems and above which the number of links per species drops off more steeply (Fig. 2).
For a set of 86 mutualistic webs30 ranging from 17 to nearly 1,000 species, this truncation is >4 times the average number of links per species in two-thirds of the examples. In these examples, the power-law exponent γ is typically ∼1; thus, the distributions are substantially less steep than are the non-ecological networks discussed above. For the set of species-rich food webs (29 to 238 species), ξ is smaller, roughly equal to the average number of links per species6,10,31, consistent with the distributions arising from the niche model and a generalized version of the cascade model32.
In sum, the distributions of links in food webs do not match other networks quantitatively33, and only match qualitatively for small ranges of numbers of links. Moreover, the ‘rich get richer’ mechanism is at odds with ecological principles5,10. For example, as more species of frugivore feed upon a fruit species, then competition for that fruit will increase, and the less likely another frugivore would include it in its diet and the more likely it would feed on other fruits instead.
Species with many links to other species in the web—the ‘richest’ ones in the analogy—may get that way simply by being the most abundant. Within a given trophic level, abundant species are more likely to receive the attention of predators, pollinators or frugivores, than are rare ones. Abundant predators probably feed on many prey species34.
Perversely, across trophic levels, the larger a species' body size, the more species on which it can feed, and thus the higher its trophic level9. Large body size and high trophic level mean lower abundance9,35, so across trophic levels it is the trophically most generalized species that are the rarest.
Whether within or across trophic levels, these patterns of strongly nested diets—whether by abundance or body size (or both)—do not imply a process of continuing community assembly. That said, Sugihara36 and colleagues37 do envision a process whereby species sequentially partition resources as they invade an ecological community. Rarer, trophically specialized species enter the community later than do generalists.
The major gap in knowledge is surely the scarcity of models exploring dynamics and rules for coexistence of large sets of species with trophic links suitably nested across trophic levels by body size (and associated attributes) and within trophic levels by abundance. Recent descriptions of food webs detailing body-size, abundance and trophic linkages9,35,38 provide a good base to develop these models.

Evolutionary patterns in diet specialization
Webs of specialists are less complex than those of generalists, so what determines whether a species specializes on few prey species or whether it takes many? The trade-offs between specialization and generalization are complex and multi-factorial4,39. Monophages (species that eat just one other species) are probably more efficient consumers than are generalists—they usually have higher per capita ingestion and assimilation rates40. Prey species also differ in the number of predators that feed on them. Experimental studies find that species consumed by multiple predators have lower predation rates than species with single predators, in contrast to the expected sum of the individual effect of each predator41. Combined, these results suggest that trophic specialists would have dynamically stronger interactions with individual prey species (and vice versa) than would more generalized species. We discuss the implications for food web dynamics below.
How are feeding interactions organized, given these patterns of relative specialization or generalization? Broad patterns are clear. For example, hummingbirds exploit different species of flowers than do insects. Within the class of insect-pollinated flowers, bees and butterflies feed on different sets of plant species4. Yet conventional wisdom suggests that such patterns should also apply within broad taxonomic groupings. Compelling examples42 of hummingbirds with long or strangely curved bills and the corresponding morphological specialization of the flowers they pollinate beg an obvious question: how ubiquitous are tight links between species of pollinator (or frugivore) and the flowers (or fruits) on which they feed?
The idea of reciprocal specializations reinforced by co-evolution seems so well entrenched that it is surprising that studies have not assessed its generality. The problem has probably been the exact measure of ‘guilds’ (groups of species exploiting the same resources by similar means, such as seed-eating birds) and the lack of a suitable null hypothesis against which to judge their improbability. Fortunately, there is an exact mathematical parallel to studies of the presence or absence of species on islands—a topic with a long, controversial history in ecology43. Both problems involve a binary matrix where, under the null hypothesis, the row and column sums must remain the same as those observed in nature. Specifically, a given species must occur on a set total number of islands, whereas a given island will host a set total number of species. Likewise, a pollinator exploits a set total number of plant species and a plant offers nectar to a set total number of species. As with biogeographical questions, the computational problem involves generating null hypotheses that obey these constraints but are not artefactually close to the observed pattern. This issue is now solved43, but no one has applied this technique to webs of mutualists. Nor, for that matter, has it been applied to sets of predators and their prey, though there are abundant sources of such information—from fish and their prey44, for example.
Mutualistic webs are strongly nested5,8. Nested links constrain reciprocal specializations, but do not completely preclude them (see Box 1). Nonetheless, the ubiquity of nestedness is a surprise, showing as it does that specialized pollinators (for example) typically exploit flowers used by many other species. Moreover, the interactions are not symmetric: the plant species that is most important to a particular pollinator may receive more attention from other species that are not so dependent upon it45.
Fragility and the paradox of complex interactions
Most differential equation models must be sufficiently ‘simple’ if the species are to persist at a locally stable equilibrium3,15,16,46. Indeed, there is a striking coincidence in the simple, sparse patterns of links observed in webs and those that models predict15,16. Sensibly parameterized models of real food webs tend to be locally stable, despite their complexity, suggesting that real webs have statistically special parameters and patterns of interactions47,48,49,50. Natural history explanations pre-date dynamical models15 in suggesting that species that overlap too extensively in their diets cannot coexist. Importantly though, models do permit tightly clustered links and extensive omnivory in systems dominated by parasitoids3 and life-history omnivores26. Because these classes of interaction dominate the group of unusually clustered models, then perhaps this is a sufficient answer: such clusters are not prohibited by the dynamics as we understand them.
Another way of phrasing the problem of complex webs is to notice that the effect of one species on the density of another diminishes with their separation in the food web, measured by the distance connecting them51. The commonness of short paths in food webs6,7 suggests that disturbances spread rapidly throughout the food web. Certainly, species may not be close if the shortest links between them are weak or intermittent. As a species feeds on more prey species, or suffers the attention of more predators, the strength of the interactions may decrease. The ‘many weak and few strong’ pattern of interactions in quantified empirical food webs52 and webs of mutualists34,53 usually correlates with diet breadth. Polyphages (species eating many other species) and prey with many predators present weak interactions, whereas strong interactions correspond to monophages and prey with only a few predators. Intriguingly, Berlow54 finds that in some systems the weak interactions have the greatest variance: they yield strong effects intermittently.
There is a long-recognized paradox: in models with random, but sensibly chosen, parameters, the probability of a locally stable equilibrium declines sharply as complexity increases46. Yet, the ever-smaller fraction of models that are stable tend to be more resilient3. (Resilient in the narrow sense that the largest eigenvalue in the linearized system will be more negative and disturbances will dissipate more quickly14,15.) McCann et al.55 developed this idea to argue the importance of weak linkages and their particular arrangement.
Indirect effects—those mediated by intermediate species—are often in the opposite direction to those expected on the basis of direct interactions alone15,51,56. They are best known from experiments in the intertidal zones of sea-shores. Menge's review57 of 23 of them is key. As species richness increased, a typical species interacted strongly with more species and through more indirect pathways.
Complex webs also pose a challenge when one considers severe changes such as the addition or removal of species. What will be the consequences of extinction rates that are now hundreds to thousands of times faster than normal58? Consider the apparent paradox of two classic results: MacArthur's model59 and Paine's experiment of removing a top predator from the inter-tidal zones60. Dynamical models of species removals show that the more polyphagous the predators, the less the effect caused by removing one of its prey species (measured as the probability that the predator will be lost3). Thus, MacArthur's model is correct, but it is incomplete. Paine showed, and models confirm15 that removing the predators of polyphages eliminates many of the latter's prey species. Thus, whether simple or highly connected model food webs are robust to the loss of species depends entirely on whether one looks at top predators or plant species15. Moreover, changes in species composition may be large while changes in the biomass may be small, and vice versa15.
Into this already complex story come recent studies using strictly topological approaches (that is, without population dynamics) to actual webs, as against abstractions of them. When the most connected species are successively removed from the web, the web is not ‘species deletion stable’3. Many species lose their only prey source (and so must become extinct in turn), and the web quickly breaks into many disconnected sub-webs61,62,63,64. By contrast, when disasters extirpate species randomly, these webs are robust, showing both little fragmentation and few secondary extinctions61,62,63,64. Perhaps well-connected species are those that are relatively abundant at their particular trophic level and so are unlikely to be lost.
In sum, we do not completely understand how species persist in complex webs when disturbances propagate quickly and apparently strongly. More realistic models that incorporate the relationships between link strength, abundance and trophic generalization may provide the answers.
Conclusions
Our knowledge of the structure of ecological networks is still incomplete in important areas that include compartments and reciprocal specialization. Species are closer than previously thought, and many webs have very dense clusters of linkages. The relationships between body size, abundance, trophic specialization and nested diets (both across and within trophic levels) are not only interesting in themselves9,35,38: they may constrain web dynamics in theoretically unexplored ways. Such constraints may be essential to explain the persistence of species in a constantly changing world, and the tolerance of current ecosystems to natural gains and losses of species as well as their vulnerability to unnaturally inflated extinction rates.
References
Pimm, S. L., Lawton, J. H. & Cohen, J. E. Food web patterns and their consequences. Nature 350, 669–674 (1991)
Cohen, J. E., Briand, F. & Newman, C. M. Community Food Webs: Data and Theory (Springer, Berlin, 1990)
Pimm, S. L. Food Webs (Univ. Chicago Press, Chicago, 2003)
Thompson, J. N. The Coevolutionary Process (Univ. Chicago Press, Chicago, 1994)
Jordano, P., Bascompte, J. & Olesen, J. M. in Plant-Pollinator Interactions: From Specialization to Generalization (eds Waser, N. M. & Ollerton, J.) (Univ. Chicago Press, Chicago, 2005)
Montoya, J. M. & Solé, R. V. Small world patterns in food webs. J. Theor. Biol. 214, 405–412 (2002)
Williams, R. J., Berlow, E. L., Dunne, J. A., Barabàsi, A.-L. & Martinez, N. D. Two degrees of separation in complex food webs. Proc. Natl Acad. Sci. USA 99, 12913–12916 (2002)
Bascompte, J., Jordano, P., Melian, C. J. & Olesen, J. The nested assembly of plant–animal mutualistic networks. Proc. Natl Acad. Sci. USA 100, 9383–9387 (2003)
Woodward, G. et al. Body-size in ecological networks. Trends Ecol. Evol. 20, 402–409 (2005)
Dunne, J. E., Williams, R. J. & Martinez, N. D. Food web structure and network theory: the role of connectance and size. Proc. Natl Acad. Sci. USA 99, 12917–12922 (2002)
Strogatz, S. Exploring complex networks. Nature 410, 268–276 (2001)
Álbert, R. & Barabàsi, A.-L. Statistical mechanics of complex networks. Rev. Mod. Phys. 74, 47–97 (2002)
Solé, R. V., Ferrer, R., Montoya, J. M. & Valverde, S. Selection, tinkering and emergence in complex networks. Complexity 8, 20–33 (2002)
Pimm, S. L. The complexity and stability of ecosystems. Nature 307, 321–326 (1984)
Pimm, S. L. The Balance of Nature? Ecological Issues in the Conservation of Species and Communities (Univ. Chicago Press, Chicago, 1991)
Solé, R. V. & Bascompte, J. Self-organization in Complex Ecosystems (Princeton Univ. Press, Princeton, 2006)
Warren, P. H. in Aspects of the Genesis and Maintenance of Biological Diversity (eds Hochberg, M. E. et al.) 142–161 (Oxford Univ. Press, Oxford, 1996)
Williams, R. J. & Martinez, N. D. Simple rules yield complex food webs. Nature 404, 180–183 (2000)
Williams, R. J. & Martinez, N. D. Limits to trophic levels and omnivory in complex food webs: theory and data. Am. Nat. 163, 458–468 (2004)
Pimm, S. L. & Lawton, J. H. Are food webs compartmented? J. Anim. Ecol. 49, 879–898 (1980)
Thomas, J. D. Mutualistic interactions in freshwater modular systems with molluscan components. Adv. Ecol. Res. 20, 125–178 (1990)
Fonseca, C. R. & Ganade, G. Asymmetries, compartments and null interactions in an Amazonian ant-plant community. J. Anim. Ecol. 65, 339–347 (1996)
Memmott, J., Godfray, H. C. J. & Gould, I. D. The structure of a tropical host-parasitoid community. J. Anim. Ecol. 63, 521–540 (1994)
Raffaelli, D. G. & Hall, S. J. Compartments and predation in estuarine food web. J. Anim. Ecol. 61, 551–560 (1992)
Krause, A. E., Frank, K. A., Mason, D. M., Ulanowicz, R. E. & Taylor, W. W. Compartments revealed in food-web structure. Nature 426, 282–285 (2003)
Pimm, S. L. & Rice, J. A. The dynamics of multispecies, multi-life-stage models of aquatic food webs. Theor. Popul. Biol. 32, 303–325 (1987)
Askew, R. R. On the biology of inhabitants of oak galls of Cynipidae (Hymenoptera) in Britain. Trans. Soc. Br. Entomol. 14, 237–269 (1961)
Strong, D. R., Lawton, J. H. & Southwood, R. Insects on Plants (Harvard Univ. Press, New York, 1984)
Amaral, L. A. N., Scala, A., Barthélemy, M. & Stanley, H. E. Classes of behavior of small-world networks. Proc. Natl Acad. Sci. USA 97, 11149–11152 (2000)
Jordano, P., Bascompte, J. & Olesen, J. M. Invariant properties in coevolutionary networks of plant-animal interactions. Ecol. Lett. 6, 69–81 (2003)
Montoya, J. M. & Solé, R. V. Topological properties of food webs: from real data to community assembly models. Oikos 102, 614–622 (2003)
Stouffer, D. B., Camacho, J., Guimera, R., Ng, C. A. & Nunes Amaral, L. A. Quantitative patterns in the structure of model and empirical food webs. Ecology 86, 1301–1311 (2005)
Garlaschelli, D., Caldarelli, G. & Pietronero, L. Universal scaling relations in food webs. Nature 423, 165–168 (2003)
Vázquez, D. P. & Aizen, M. A. Null model analyses of specialization in plant-pollinator interactions. Ecology 84, 2493–2501 (2003)
Cohen, J. E., Jonsson, T. & Carpenter, S. R. Ecological community description using the food web, species abundance, and body size. Proc. Natl Acad. Sci. USA 100, 1781–1786 (2003)
Sugihara, G., Bersier, L.-F., Southwood, T. R. E., Pimm, S. L. & May, R. M. Predicted correspondence between species abundances and dendrograms of niche similarities. Proc. Natl Acad. Sci. USA 100, 5246–5251 (2003)
Cattin, M. F., Bersier, L.-F., Banasek-Richter, C., Baltensperger, M. & Gabriel, J.-P. Phylogenetic constraints and adaptation explain food-webs structure. Nature 427, 835–839 (2004)
Jonsson, T., Cohen, J. E. & Carpenter, S. R. Food webs, body size and species abundance in ecological community description. Adv. Ecol. Res. 36, 1–84 (2005)
Hawkins, B. A. Pattern and Process in Host-Parasitoid Communities (Cambridge Univ. Press, Cambridge, UK, 1994)
Rana, J. S., Dixon, A. F. G. & Jarosik, V. Costs and benefits of prey specialization in a generalist insect predator. J. Anim. Ecol. 71, 15–22 (2002)
Sih, A., Englund, G. & Wooster, D. Emergent impacts of multiple predators on prey. Trends Ecol. Evol. 13, 350–355 (1998)
Temeles, E. J. & Kress, W. J. Adaptation in a plant-hummingbird association. Science 300, 630–633 (2003)
Sanderson, J. G. Testing ecological patterns. Am. Sci. 80, 332–339 (2000)
Froese, R. & Pauly, D. (eds) FishBase. http://www.fishbase.org (2005)
Bascompte, J., Jordano, P. & Olesen, J. M. Asymmetric coevolutionary networks facilitate biodiversity maintenance. Science 312, 431–433 (2006)
May, R. M. Stability and Complexity in Model Ecosystems (Princeton Univ. Press, Princeton, 1974)
Yodzis, P. Local trophodynamics and the interaction of marine mammals and fisheries in the Benguela ecosystem. J. Anim. Ecol. 67, 635–658 (1998)
McCann, K. S. The diversity-stability debate. Nature 405, 228–233 (2000)
Neutel, A.-M., Heesterbeek, J. A. P. & de Ruiter, P. C. Stability in real food webs: weak links in long loops. Science 296, 1120–1223 (2002)
Emmerson, M. & Raffaelli, D. Body size, patterns of interaction strength and the stability of a real food web. J. Anim. Ecol. 73, 399–409 (2004)
Abrams, P., Menge, B. A., Mittelbach, G. G., Spiller, D. & Yodzis, P. in Food Webs: Integration of Patterns and Dynamics (eds Polis, G. A. & Winemiller, K. O.) 371–395 (Chapman & Hall, New York, 1996)
Berlow, E. et al. Interaction strengths in food webs: issues and opportunities. J. Anim. Ecol. 73, 585–598 (2004)
Jordano, P. Patterns of mutualistic interactions in pollination and seed dispersal: connectance, dependence asymmetries, and coevolution. Am. Nat. 129, 657–677 (1987)
Berlow, E. L. Strong effects of weak interactions. Nature 398, 330–334 (1999)
McCann, K., Hastings, A. & Huxel, G. R. Weak trophic interactions and the balance of nature. Nature 395, 794–798 (1998)
Wootton, J. T. Indirect effects in complex ecosystems: recent progress and future challenges. J. Sea Res. 48, 157–172 (2002)
Menge, B. A. Indirect effects in marine rocky intertidal interaction webs: patterns and importance. Ecol. Monogr. 65, 21–74 (1995)
Pimm, S. L. World According to Pimm: a Scientist Audits the Earth (McGraw-Hill, New York, 2001)
MacArthur, R. H. Fluctuations of animal populations and a measure of community stability. Ecology 36, 533–536 (1955)
Paine, R. T. A note on trophic complexity and community stability. Am. Nat. 103, 91–93 (1969)
Álbert, R., Jeong, H. & Barabàsi, A.-L. Error and attack tolerance of complex networks. Nature 406, 378–382 (2000)
Solé, R. V. & Montoya, J. M. Complexity and fragility in ecological networks. Proc. R. Soc. Lond. B 268, 2039–2045 (2001)
Dunne, J. A., Williams, R. J. & Martinez, N. D. Network structure and biodiversity loss in food webs: robustness increases with connectance. Ecol. Lett. 5, 558–567 (2002)
Memmot, J., Waser, N. M. & Price, M. V. Tolerance of pollination networks to species extinctions. Proc. R. Soc. Lond. B 271, 2605–2611 (2004)
Watts, D. J. & Strogatz, S. H. Collective dynamics in ‘small-world’ networks. Nature 393, 440–442 (1998)
Hall, S. J. & Raffaelli, D. Food web patterns: lessons from a species-rich web. J. Anim. Ecol. 60, 823–842 (1991)
Polis, G. A. & Strong, D. R. Food web complexity and community dynamics. Am. Nat. 147, 813–846 (1996)
Root, R. B. The niche exploitation pattern of the blue-gray gnatcatcher. Ecol. Monogr. 37, 317–350 (1967)
Kitching, R. L. in Phytotelmata: Terrestrial Plants as Hosts of Aquatic Insect Communities (eds Frank, H. & Lounibos, P.) 205–222 (Plexus, New Jersey, 1983)
Kitching, R. L. Food Webs and Container Habitats (Cambridge Univ. Press, Cambridge, UK, 2000)
Polis, G. A. Complex desert food webs: an empirical critique of food web theory. Am. Nat. 138, 123–155 (1991)
Cohen, J. E. et al. Improving food webs. Ecology 74, 252–258 (1993)
Raffaelli, D. G. & Hall, S. J. in Food Webs: Integration of Patterns and Dynamics (eds Polis, G. A. & Winemiller, K. O.) 185–191 (Chapman & Hall, New York, 1996)
Emmerson, M. E., Montoya, J. M. & Woodward, G. in Dynamic Food Webs (eds de Ruiter, P. C., Wolters, V. & Moore, J. C.) 179–197 (Academic, San Diego, 2005)
Melian, C. J. & Bascompte, J. Food web cohesion. Ecology 85, 352–358 (2004)
Wheelwright, N. T., Haber, W. A., Murray, K. G. & Guindon, C. Tropical fruit eating birds and their food plants: a survey of a Costa Rican lower montane forest. Biotropica 16, 173–192 (1984)
Inoue, T., Kato, M., Kakutani, T., Suka, T. & Itino, T. Insect-flower relationship in the temperate deciduous forest of Kibune, Kyoto: an overview of the flowering phenology and the seasonal pattern of insect visits. Contrib. Biol. Lab. Kyoto Univ. 27, 377–463 (1990)
Reagan, D. P. & Waide, R. B. The Food Web of a Tropical Rain Forest (Univ. Chicago Press, Chicago, 1996)
Acknowledgements
We thank J. Dunne, D. Rafaelli, M. Emmerson and G. Woodward for comments; P. Jordano and R. Waide for the pictures and data for Fig. 2; and S. Valverde for assistance with the figures. This work was supported by an NERC Fellowship (J.M.M.) and the Santa Fe Institute (grant to R.V.S.).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
Reprints and permissions information is available at npg.nature.com/reprintsandpermissions. The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Montoya, J., Pimm, S. & Solé, R. Ecological networks and their fragility. Nature 442, 259–264 (2006). https://doi.org/10.1038/nature04927
Issue Date:
DOI: https://doi.org/10.1038/nature04927
This article is cited by
-
Biological, ecological and trophic features of invasive mosquitoes and other hematophagous arthropods: What makes them successful?
Biological Invasions (2024)
-
Co-Occurrence Patterns of Soil Fungal and Bacterial Communities in Subtropical Forest-Transforming Areas
Current Microbiology (2024)
-
An overview of the ecological roles of Mediterranean chondrichthyans through extinction scenarios
Reviews in Fish Biology and Fisheries (2024)
-
Soil bacterial and fungal communities resilience to long-term nitrogen addition in subtropical forests in China
Journal of Forestry Research (2024)
-
Erosion and deposition significantly affect the microbial diversity, co-occurrence network, and multifunctionality in agricultural soils of Northeast China
Journal of Soils and Sediments (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
